










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.2 San José Jun. 2012
Biodiversidad íctica de praderas de pasto marino de la costa noroeste del Golfo de Cariaco, Venezuela
*Dirección para correspondencia:
Abstract
Seagrasses are highly productive coastal ecosystems with a high diversity and abundance of fishes, very important to support artisanal fisheries. We analyzed the fish community structure of Thalassia testudinum in the communities of Manzanillo (M) and La Brea (LB), Northwest coast of Cariaco Gulf, Venezuela. Samples were taken monthly (Nov. 2006-Oct. 2007) from each place, using a beach net. A total of 34 810 fishes were captured, grouped into 13 orders, 36 families and 83 species. In both areas (M and LB), the number of species was similar, but a variation in their abundance was found: a total of 55 species and 13 210 organisms for M, and 58 species and 21 600 organisms for LB. The most abundant species and those with the highest biomasses in both areas were: Nicholsina usta, Haemulon boschmae, H. steindachneri, Harengula jaguana, Halichoeres bivittatus and Hemiramphus brasiliensis. The occasional visitors were the most frequent community components with a 59%, the cyclical and permanent residents were represented by the 22% and 19%, respectively. The H’n average for M was of 1.71±0.64bits/ind., while for LB was of 1.95±0.51bits/ind. The diversity values were directly related to the evenness and inversely related to the dominance. The low values of similarity indexes among localities allow us to assert that these fish communities are dissimilar, because of the structure of each Thalassia meadow and their connectivity with other systems. Rev. Biol. Trop. 60 (2): 635-648. Epub 2012 June 01.
Key words: fishes, Cariaco Gulf, ecology, seagrass meadows, diversity.
Resumen
Los pastos marinos son ecosistemas costeros de alta productividad, con gran diversidad y abundancia de peces, la cual es aprovechada por pescadores artesanales. En este estudio se analizó la estructura comunitaria íctica de praderas de Thalassia testudinum en Manzanillo (M) y La Brea (LB), costa noroeste del Golfo de Cariaco, Venezuela. También, se realizaron muestreos mensuales (11-2006 al 10-2007), con la utilización de una red playera. Se capturaron 34 810 organismos agrupados en 13 órdenes, 36 familias y 83 especies. En ambas zonas, el número de especies fue similar, aunque el número de organismos vario, y se encontró para M un total de 55 especies y 13 210 organismos y para LB 58 especies y 21 600 organismos. Las especies más abundantes y de mayor biomasa en el área muestreada fueron: Nicholsina usta, Haemulon boschmae, H. steindachneri, Harengula jaguana, Halichoeres bivittatus y Hemiramphus brasiliensis. Los visitantes ocasionales fue el componente comunitario más frecuente con 59%, los cíclicos y los residentes permanentes obtuvieron 22 y 19%, respectivamente. En M la H’n fue de 1.71±0.64bits/ind; entretanto en LB fue 1.95±0.51bits/ind. Los valores de la diversidad estuvieron relacionados directamente con la equitabilidad e inversamente con la dominancia. Los bajos valores del índice de similaridad, entre localidades permite establecer que estas comunidades ícticas son disimiles, debido quizás a la estructuración de cada pradera de Thalassia y a la conectividad con otros sistemas.
Palabras clave: peces, Golfo de Cariaco, ecología, pastos marinos, diversidad.
Los hábitats de pastos marinos constituyen un ecosistema marino costero esencial alrededor del mundo (Den Hartog & Kuo 2006), están entre los biotopos más productivos de la biosfera y pueden incrementar la biodiversidad de sus organismos asociados (Boström & Bonsdorff 2000), debido a que, son importantes sitios de criadero y alimentación para numerosas poblaciones de peces y de invertebrados (Nagelkerken et al. 2000), adicionalmente, representan áreas muy favorables para el establecimiento de especies de peces de interés comercial, por lo que se consideran entornos de una diversa comunidad biótica, donde la macrofauna acuática más importante la constituye este grupo, dadas sus características de abundancia, diversidad y distribución espacio-temporal (Soberón-Chávez 1984).
La complejidad estructural del sustrato es un factor determinante en la organización de las comunidades de peces, y en los pastos marinos dicha complejidad está determinada, principalmente, por la talla de las hojas y la cobertura vegetal; factores que han sido relacionados con la riqueza y abundancia de peces (Aliaume et al. 1990).
Desde el trabajo pionero de Petersen, realizado a principios del siglo XX, muchos estudios muestran una mayor abundancia y riqueza de especies de invertebrados bentónicos y de peces en el interior de los fondos cubiertos por fanerógamas marinas (Pollard 1984), razón por la cual es ampliamente conocido que las áreas costeras en donde se presentan estas fanerógamas se encuentren habitadas por una ictiofauna más diversa y abundante, que aquellas zonas sin vegetación, además muestran similares magnitudes de productividad y biomasa de peces, como las de arrecifes de coral y las de manglares, aunque han tenido menor atención en términos de manejo e investigación (Gullström & Dahlberg 2004).
La fauna asociada a pastos marinos puede presentar modificaciones en cuanto a su estructura y depende de la ubicación con respecto a los límites del sistema y la interacción con hábitats adyacentes. En los trópicos es común encontrar praderas, de pastos marinos, muy cercanas a otros ecosistemas como los arrecifes de coral y los manglares, por lo tanto, también se han estudiado las interacciones y fluctuaciones que se dan entre ellos, como los trabajos realizados por Pollard (1984), Baelde (1990) y Nagelkerken et al. (2002). A su vez, esta fauna puede presentar diversas manifestaciones como una respuesta a las presiones ambientales o antropogénicas.
En el Mar Caribe, son muchos los estudios que se han realizado sobre la estructura comunitaria de peces asociados a pastos marinos, Vargas-Maldonado & Yáñez-Arancibia (1987) en la Laguna de Términos, México; González-Gándara et al. (2006), quienes mediante censos visuales estructuraron la comunidad de peces ligados a T. testudinum de Arrecife Lobos, y Acosta et al. (2007), que determinaron las características espaciales y temporales de peces de pastos marinos en los Cayos de Florida. Por su parte, Baelde (1990) estableció diferencias en las estructuras de peces en Guadalupe, India Occidental Francesa, y Bouchon et al. (1992) estudiaron los peces de la Isla de Martinica. Mientras que en las costas de Colombia, Acero (1980) examinó los peces de la bahía del Parque Nacional Tayrona; Victoria & Gómez (1984) en la Isla de San Andrés; y Acero & Garzón (1986) en las Islas del Rosario y de San Bernardo, en las cuales señalan tres nuevos registros de peces para el Caribe.
En Venezuela se han realizado trabajos los cuales han contribuido al conocimiento de los peces de praderas marinas como los de Villamizar (1993), Díaz (1997) y López-Ordaz et al. (2009a), en los Parques Nacionales: Archipiélago los Roques y Morrocoy, respectivamente. Sin embargo, la mayor parte de las investigaciones se han concentrado en las costas orientales del país, se pueden mencionar el de Martínez (1971) que comparó la fauna ictiológica de la laguna Grande del Obispo y la Bahía de Mochima; San Cristóbal (1984) analizó la estructura de la comunidad ictiológica en dos praderas de Thalassia en la Bahía de Mochima; Gómez (1987) analizó la taxocenosis de peces en praderas de T. testudinum en la Bahía de Charagato; Ruiz (1992) evaluó dos comunidades de peces diurno-nocturno en el Golfo de Cariaco, Valecillos (1993) determinó la estructura ecológica de la comunidad de peces en el sistema costero Chacopata-Bocaripo; Méndez (1995) estudió la ictiofauna en una pradera en el Saco del Golfo de Cariaco; De Grado et al. (2000) analizó la ictiofauna de la ensenada Grande del Obispo (Laguna Grande); Allen et al. (2004) estudiaron la estructura de peces en la parte sureste del Golfo de Cariaco. Allen et al. (2007) analizaron las comunidades ícticas en Thalassia de la costa norte del golfo de Santa Fe, estado Sucre. Mientras que los peces asociados a T. testudinum de la zona costera de Cumaná fue examinada por López et al. (2009b).
Los estudios de biodiversidad íctica en las comunidades de pastos marinos son básicos y de gran relevancia, puesto que, permiten obtener registros temporales en áreas que pueden verse afectadas por el uso irracional de los recursos naturales. Debido a la importancia ecológica del Golfo de Cariaco, a la existencia de praderas de T. testudinum en diferentes zonas del mismo y a lo bien representados que están los peces en dichas áreas, se realizó un inventario de la ictiofauna que aloja este sistema en su costa noroccidental, se evalúo la composición y estructura comunitaria, en las localidades de Manzanillo y La Brea, no sólo por la importancia ecológica y pesquera real y potencial, sino como punto de comparación para futuras evaluaciones en el desarrollo del sistema.
Materiales y métodos
Área de estudio: El estudio se llevó a cabo en la zona costera noroccidental del Golfo de Cariaco, donde se seleccionaron dos localidades con extensas praderas de Thalassia testudinum: Manzanillo (10°30’58” N - 64°13’41”W) y La Brea (10°33’34” N - 64°11’54” W).
Manzanillo es una playa amplia y abierta, con oleaje fuerte, la pradera muy compacta, sin parches de arena, se ubica desde los 40m de la orilla hasta los 500m de distancia, la cual abarca alrededor de 300m2, con una profundidad promedio 3.2±0.4m. Las hojas de la Thalassia miden 10.2±2.0cm de largo. Por su parte, La Brea es una playa semicerrada, tipo ensenada, con poco oleaje; su pradera es muy densa, se encuentra desde los 50m de la orilla hasta los 250m de distancia y ocupa aproximadamente 180m2, con una profundidad de 1.3±0.1m. Las hojas miden 16.4±1.3cm de largo; hacia el lado este de la localidad existe un gran complejo arrecifal muy bien desarrollado.
Trabajo de campo: Ahora bien, se ejecutó un muestreo diurno mensual, por estación, entre las nueve y 11 de la mañana, en la segunda quincena de cada mes, desde noviembre 2006-octubre 2007. Asimismo, se determinó la temperatura y salinidad del agua, con la utilización de una sonda multiparamétrica, (YSI modelo 30/10 FT), en cada muestreo. Para la captura de las muestras biológicas se empleó una red de arrastre playero de 60mx4m, con un diámetro de malla de una pulgada en los extremos y ¼ en el centro de la red. La técnica de muestreo consistió en colocar la red en el mar para formar un semicírculo, además se utilizó una embarcación artesanal (peñero) con motor fuera de borda en constante movimiento; posteriormente, la red fue arrastrada de forma manual por los bordes hasta la orilla, luego se realizó una calada por estación, en cada muestreo. Los peces capturados fueron colocados en bolsas plásticas previamente etiquetadas (localidad y fecha), y se guardaron en cavas con hielo hasta su traslado al laboratorio.
Trabajo de laboratorio: Los organismos fueron identificados hasta la categoría de especie, y se utilizaron las claves taxonómicas descritas por Cervigón (1991, 1993, 1994, 1996). A cada ejemplar se le determinó la masa corporal en gramos y la longitud total. Un ejemplar de cada especie fue depositado en el Laboratorio de Ecología de Peces Marinos, del Departamento de Biología, Universidad de Oriente.
Por consiguiente, se obtuvo la Abundancia (A) y Biomasa (B), y se determinaron los Índices de Diversidad numérica (H’n) de Shannon-Wiener, según Krebs (1989) y de Diversidad por biomasa (H’b) de Wilhm (1968). También se calcularon los índices de Equitabilidad (J’n) de Lloyd & Ghelardi, según Krebs (1989), Constancia o Frecuencia de ocurrencia, según Amezcua-Linares & Yáñez-Arancibia (1980), el Índice de Dominancia de McNaughton (Krebs 1989), el Índice Biológico de Sanders (1960) y el Índice cuantitativo de Similaridad de Morisita-Horn (Krebs 1989). A todas las variables se les aplicaron las pruebas de Normalidad de Kolmogorov-Smirnov y Homocedasticidad de las varianzas de Cochrans (Zar 1996). Una vez comprobado que los datos cumplían los supuestos, se emplearon análisis de varianza multifactorial (ANOVA) con la utilización del paquete estadístico Statgraphics plus 5.1, para detectar la existencia de diferencias espacio-temporales del número de especies, número de organismos, los parámetros comunitarios y los parámetros físicos-químicos. A las variables que presentaron diferencias estadísticas se les aplicó la prueba a posteriori LSD. Para observar el efecto que tenían los parámetros físico-químicos sobre las variables ecológicas se aplicó un análisis de componentes principales (Johnson & Wichern 1992).
Resultados
Parámetros físico-químicos: La salinidad fluctuó, para las dos estaciones, entre 34-37 en octubre y febrero respectivamente, con promedio de 35.9±0.9 para Manzanillo y 35.4±1.1 para La Brea. La temperatura media para Manzanillo fue 26.5±1.3°C, la misma varió entre 23.8°C (febrero) y 28.6°C (octubre), mientras que en La Brea la temperatura promedio fue 27.6±1.9°C, la cual osciló entre 24.4°C (febrero) y 30.5°C (septiembre). Tanto la salinidad como la temperatura, presentaron diferencias estadísticamente significativas entre meses (p=0.0019 y p=0.0020, respectivamente), mientras que entre estaciones solamente la temperatura presentó diferencias significativas (p=0.0103) según las pruebas de ANOVA. El análisis de componentes principales mostró que no había relación entre los parámetros físicoquímicos y los comunitarios, estos últimos tienen un comportamiento que es independiente de las fluctuaciones ambientales imperantes.
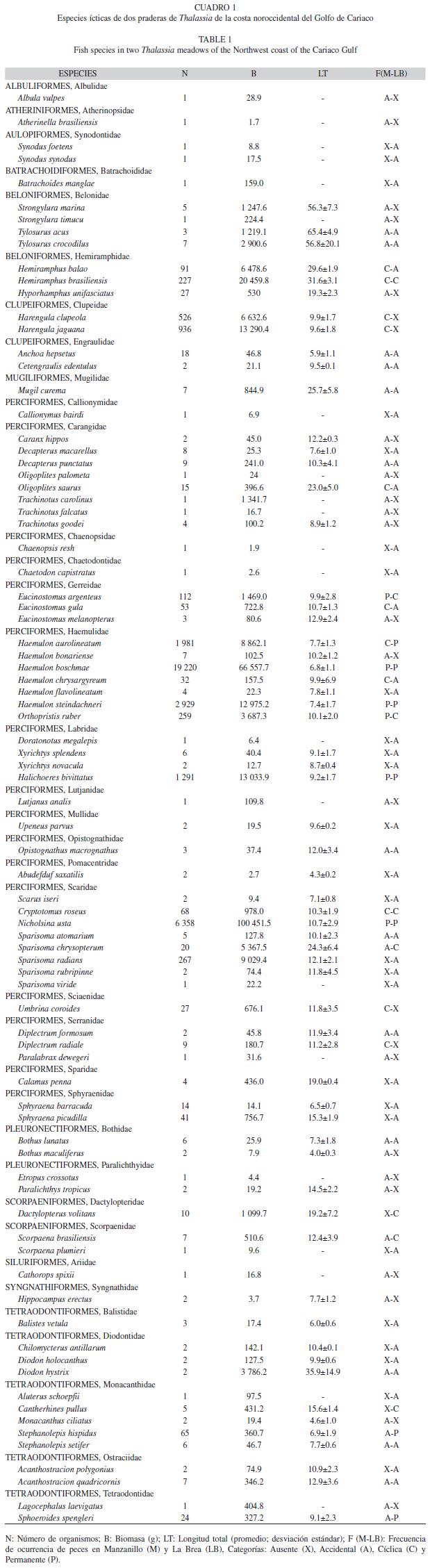
Composición taxonómica: En total se capturaron 34 810 organismos, con una biomasa de 291.1kg. Se identificaron 83 especies de peces, distribuidas en 13 órdenes y repartidas en 36 familias. Del total de las especies capturadas, 30 (36.1%) fueron comunes en ambas estaciones, 25 (30.2%) estuvieron restringidas a playa Manzanillo, mientras que 28 (33.7%) fueron exclusivas de La Brea (Cuadro 1).
De manera general, la mayoría de los peces capturados fueron juveniles (85.5%), por lo tanto, los valores promedios de sus longitudes totales (LT) fueron bajos, aunque se presentaron ejemplares adultos en algunas especies. Las tallas de LT se distribuyeron en un rango de 1.0 a 106.0cm para Haemulon boschmae y Tylosurus crocodilus, respectivamente. Sin embargo, la talla promedio para todos los ejemplares fue 9.3cm y solamente 12 especies tuvieron una longitud total media de más de 15.0cm.
El 20% del total de las especies de peces identificadas son consideradas importantes para las pesquerías comerciales, entre estas resaltan el pargo criollo Lutjanus analis, el pámpano amarillo Trachinotus carolinus, los cuales constituyen grandes rubros pesqueros a nivel mundial, mientras que el 61% corresponde a especies de escaso interés comercial o que son capturados como pesca de subsistencia, tales como: el futre Lagocephalus laevigatus y el agujón o marao Strongylura marina. También, se encontró una gran abundancia de la especie de loro Sparisoma radians, la cual presenta un interés potencial para ser explotado.
La Riqueza de especies en playa Manzanillo fue 15.6±4.9, osciló entre ocho y 26 en febrero y agosto, respectivamente, mientras que en La Brea fue 15.4±4.0, el cual varió entre nueve-22 en agosto y septiembre, respectivamente. Para cada estación, el número de especies fue similar, entre tanto que el número de organismos varió, y se encontró para Manzanillo un total de 55 especies y 13 210 organismos, y para La Brea 58 especies y 21 600 organismos. En Manzanillo la abundancia fue 1 100.8±1 039.1, osciló entre 97 organismos (marzo) y 4 630 (julio), mientras que para La Brea fue 1 800.2±1 100.8 individuos, el mismo varió entre 76 (diciembre) y 6 741 (julio).
El análisis de varianza indicó la existencia de diferencias estadísticamente significativas de la abundancia con respecto a los meses de muestreo (p=0.0095). La prueba a posteriori LSD evidenció la formación de tres grupos: el primero formado por los valores promedios mensuales más bajos, el segundo integrado por los meses que presentaron los valores intermedios, y por último, el mes que presentó el valor promedio más alto (julio). Las menores abundancias se mostraron en aquellos meses en donde se ve involucrado el proceso de surgencia y las abundancias moderadamente altas, las cuales corresponden a los meses de época de calma o no surgencia, con la excepción del mes de julio, donde se pescó un número importante de Haemulon boschmae.
Por su parte, la biomasa íctica en Manzanillo fue 90.0kg, el mismo varió entre 2kg (marzo) y 24.2kg (julio), con un promedio de 7.5±7.0kg; mientras que en La Brea se obtuvieron 201.1kg, con valores más bajos en marzo (3.1kg), mayores en julio (34.1kg), y un valor medio 16.8±12.0kg. Para la biomasa, el análisis de varianza mostró diferencias estadísticamente significativas entre los meses (p=0.0261), y entre las estaciones (p=0.0056).
Tanto para la abundancia como para la biomasa, los mayores valores se obtuvieron en época de calma (no surgencia), con excepción del mes de enero donde la biomasa tuvo un repunte debido a un pequeño cardumen de Hemiramphus brasiliensis que aportó elevado peso al total de la captura.
De manera general, las familias de peces con mayor riqueza pertenecen al orden Perciformes, estas fueron: Carangidae y Scaridae (8), Haemulidae (7), Monacanthidae (5), Belonidae y Labridae (4). En ambas estaciones, las familias más representativas con respecto a la abundancia y biomasa fueron: Haemulidae y Scaridae, otras familias de importancia relativa fueron Clupeidae y Belonidae para Manzanillo (Fig. 1) y Hemiramphidae y Labridae para La Brea (Fig. 2).
Diez especies: Haemulon boschmae, Nicholsina usta, H. steindachneri, H. aurolineatum, Halichoeres bivitattus, Harengula jaguana, H. clupeola, Sparisoma radians, Orthopristis ruber y Hemiramphus brasiliensis, representaron el 97% de la abundancia total y el 88% de la biomasa total para las dos praderas de pasto marino estudiadas. Estas mismas especies, de manera general, presentan los valores más elevados de dominancia, en donde el valor promedio más alto fue en julio (95.72%) y el más bajo en octubre (41.37%). En Manzanillo H. boschmae presenta el mayor porcentaje de dominancia en nueve de los 12 muestreos, mientras que N. usta domina en 11 de los 12 muestreos en La Brea.
Generalmente, dentro de los componentes comunitarios los más frecuentes fueron los visitantes ocasionales con 59%, seguido por los visitantes cíclicos 22% y los residentes permanentes 19%. Los visitantes accidentales u ocasionales predominaron en ambas estaciones, con 69% en Manzanillo y 72% en La Brea.
Con respecto a la diversidad (H’n y H’b) y a la equitabilidad (J’n y J’b), se encontró que en Manzanillo, la H’n media fue 1.71±0.64bits/ind., el cual fluctuó entre 0.53 y 2.61bits/ind., para julio y octubre, respectivamente; la J’n promedio fue 0.44±0.16, y osciló entre 0.13 (julio) y 0.67 (marzo), H’b y J’b fueron 2.50±0.63bits/g y 0.64±0.13, respectivamente. Por otra parte en La Brea, la H’n promedio fue 1.95±0.51bits/ind., y fluctuó entre 1.07 (octubre) y 2.60bits/ind. (marzo), la J’n media fue 0.50±0.11, y osciló entre 0.29 (octubre) y 0.67 (marzo). Los promedios de H’b y J’b fueron 1.97±0.43bits/g y 0.51±0.10, respectivamente. Los análisis de varianza tanto para meses como para estaciones indicaron que no existían diferencias estadísticamente significativas con respecto a H’n, H’b, J’n, ni J’b (p>0.05).
Asimismo, la diversidad muestra cambios a lo largo del año, pero estos no siguen un patrón estacional bien marcado, debido a que, puede apreciarse una tendencia oscilatoria moderada entre los meses de muestreo. Además, se cree que el aumento o disminución de este índice no presentó asociación particular con alguna época del año. Estas variaciones se deben, principalmente, a la presencia de especies dominantes. Para julio, en Manzanillo se presentó una H’n mínima de 0.53bits/ind., en donde se puede observar que el muestreo fue dominado por H. boschmae, con 4 325 individuos de 4 630 capturados. Y hay que tomar en cuenta que los valores de la diversidad están relacionados directamente con la equitabilidad e inversamente con la dominancia.
Por su parte, los resultados del índice biológico revelaron cuales fueron las especies típicas y características de estos sistemas de pastos marinos. El análisis de IB, realizado para las 10 especies más abundantes del total, reveló que en Manzanillo, por su abundancia y constancia, tres especies tipifican esa comunidad íctica, estas son: Haemulon boschmae, Nicholsina usta y Haemulon steindachneri, estas especies obtuvieron un IB superior al 50% del valor máximo teórico (120), mientras que para La Brea, cuatro especies están por encima del 50% del máximo valor calculado (240), estas son: Nicholsina usta, Halichoeres bivittatus, Haemulon boschmae y Sparisoma radians. Cabe destacar que H. boschmae y N. usta, obtuvieron el máximo valor de cada estación (79.6% y 95.4%, respectivamente), en consecuencia estas especies deben nominar la comunidad íctica de las áreas estudiadas.
Los valores de similaridad entre las dos localidades estudiadas, consideran la ausencia o presencia de las especies y su abundancia, según el coeficiente de afinidad cuantitativo, Morisita-Horn arrojó un valor de 0.29 el cual se considera un valor bajo, en donde se toma en cuenta que el intervalo de valores para este índice va de cero, cuando no hay especies compartidas entre ambos sitios, hasta uno, cuando los dos sitios tienen una similar composición de especies, de allí que el resultado obtenido permite inferir que las dos comunidades ícticas son disímiles.
Discusión
El número de especies capturadas puede considerarse elevado si se compara con otros estudios de comunidades de peces asociados a pastos marinos, en la región oriental (Ruiz 1992, Méndez 1995, Allen et al. 2004, Silva 2009) y occidental del país (Villamizar 1993, Díaz 1997), otras regiones del Caribe (Rosales-Casián 2004), e inclusive de otras latitudes (Gullström & Dahlberg 2004, Unsworth 2007).
La riqueza de especies de un área en particular va a estar determinada por múltiples factores como lo son: la extensión del área, la densidad de la vegetación, la duración del periodo de muestreo, la proximidad a otros hábitats como arrecifes de coral, manglares, parches arenosos y fangosos, la realización de muestreos diurnos y nocturnos, así como los tipos de artes de pesca utilizados (Acero 1980, Ruiz 1992).
Los resultados obtenidos del número de especies para las estaciones estudiadas mostraron poca variabilidad temporal, generalmente la variación del número de especies suele asociarse a los cambios temporales de los factores ambientales; sin embargo, a pesar de que la temperatura y la salinidad registrados presentaron dos periodos de altas y bajas temperaturas producto de la surgencia y las épocas de lluvia-sequia, no se observó un patrón definido en el número de especies.
Es de gran importancia señalar que de las especies capturadas, 14 no habían sido reportadas para el Golfo de Cariaco: Chaenopsis resh (Robins & Randall 1965), Doratonotus megalepis (Günther 1862), Xyrichtys novacula (Linnaeus 1758), Xyrichtys splendens (Castelnau 1855), (Cryptotomus roseus Cope 1871), Sparisoma atomarium (Poey 1861), Synodus synodus (Linnaeus 1758), Hemiramphus balao (Lesueur 1821), Decapterus punctatus (Cuvier 1829), Bothus lunatus (Linnaeus 1758), B. maculiferus (Poey 1860), Scorpaena brasiliensis (Cuvier 1829), Monacanthus ciliatus (Mitchill 1818) y Aluterus schoepfii (Walbaum 1792). Con esto aumenta a 247 el total de especies ícticas dentro del Golfo de Cariaco.
Los pastos marinos son utilizados por muchas especies de peces, de diferentes clases y edades, aunque, generalmente están constituidos por individuos juveniles e inmaduros, el elevado número de ejemplares juveniles obtenidos en este estudio lo ratifica. Un gran número de especies, tanto de interés ecológico como pesquero, dependen al menos en una parte de su ciclo de vida de estos ambientes, y son muchas las hipótesis que se han propuesto para explicar la alta abundancia de los peces juveniles en los pastos marinos, basadas en la protección ante predadores y la abundancia de alimento, y utilizan estas áreas como zonas de reproducción, criadero y alimentación (Nagelkerken et al. 2000, Aguilar 2004, Gullström & Dahlberg 2004, Acosta et al. 2007).
Con respecto a la abundancia y a la biomasa íctica, se tiene para las dos estaciones valores elevados. Sanders (1960), resalta que las abundancias son un criterio más valido que la biomasa para representar la estructura de una comunidad. Esto, se debe a que la presencia de un animal grande que aparece en una muestra pero que no es típico de la misma, puede alterar la distribución de la biomasa hasta en dos órdenes de magnitud; lo cual produce estimaciones de la estructura comunitaria con un sesgo asociado que será significativamente mayor que el permitido en este tipo de investigaciones.
La estacionalidad de las variables ambientales parece estar influyendo en la abundancia y biomasa capturada, dado que los menores valores fueron obtenidos en la época de surgencia y los valores más altos corresponden a los meses donde el proceso de surgencia se encontraba en época de calma, aún y cuando estadísticamente no existe relación entre este factor con las variables ecológicas (p>0.05). Y como es sabido en las áreas de surgencia costera, influenciadas por los vientos alisios, se da el fenómeno de floración de fitoplancton, el cual sirve de sustento a una abundante comunidad de diversos grupos zoológicos, de los cuales los peces se alimentan.
Haemulon boschmae es una especie muy abundante en algunas localidades de la región oriental de Venezuela, donde se le puede observar a unos pocos metros de la superficie, habitan sobre fondos someros en áreas protegidas de aguas claras, las cuales forman grandes asociaciones o ensamblajes en toda la columna de agua, esta especie constituye un alimento importante para pargos, cunas, meros y otras especies de importancia económica; mientras que H. steindachneri, es otro Haemulidae de los más abundantes, que forman grandes agregaciones en fondos arenosos y con praderas de Thalassia (Cervigón 1993). Por su parte los Scaridae Nicholsina usta y Sparisoma radians son peces sumamente abundantes en todas las playas de la región nororiental, que se encuentran, preferiblemente, en zonas de aguas someras, también con praderas de T. testudinum (Cervigón 1994).
Cabe destacar que la mayor riqueza la presentó la familia Carangidae, con especies de gran interés pesquero, aunque con baja abundancia, por lo que se presume que estos hábitats de Thalassia son utilizados por estas especies para fines particulares, por lo que fueron catalogadas como especies accidentales, puesto que no tienen un patrón definido de utilización del área y pueden entrar y salir a la pradera solamente para protegerse y/o alimentarse.
Los visitantes ocasionales o accidentales predominaron en ambas localidades, según su estatus, aunque la mayoría de las especies fueron consideradas como ocasionales, estas nunca lograron desplazar a las residentes permanentes que alcanzaron los mayores valores en abundancia numérica y biomasa durante el tiempo de estudio. La mayoría de las especies de peces marinos acuden a las zonas de pastos marinos, manglares y áreas estuarinas, para la alimentación o refugio en alguna etapa de su ciclo de vida y se mantiene un patrón estacional en un hábitat particular cuando las condiciones de productividad son adecuadas (Unsworth 2007), lo cual explicaría las moderadas diversidades de peces en las localidades de Manzanillo y La Brea.
Menares & Sepúlveda (2005), consideran núcleos de alta diversidad, aquellas zonas con valores iguales o superiores a 2bits/ind., y núcleos de baja diversidad las zonas con valores menores a 2bits/ind. A pesar de esto, los valores obtenidos de diversidad total para Manzanillo y La Brea podrían considerarse bajos, al compararlos con otros estudios del nororiente de Venezuela (Valecillos 1993, De Grado et al. 2000). La variabilidad de la diversidad se debe principalmente a la dominancia de especies durante los muestreos, los cuales indican que cuando los valores son mínimos, se debe a la presencia de especies dominantes, por lo que no existe una distribución equitativa del número de organismos por especie.
Finalmente, el análisis de similaridad de Morista-Horn estableció una baja afinidad entre las dos localidades estudiadas, entonces se tiene dos áreas de pastos marinos relativamente cercanas pero distintas en su composición íctica, por lo que se puede inferir que esto se debe a las diferencias en la morfología de fondo y forma de la playa (protegida o expuesta), también la estructura y frondosidad de las Thalassia puede ser uno de los factores determinantes en la estructuración de las comunidades de peces presentes. Estos sistemas costeros varían ampliamente en función de la heterogeneidad estructural o complejidad. En el caso de los pastos marinos la complejidad está determinada principalmente por la talla de las hojas y la cobertura vegetal, factores que han sido relacionados con la riqueza y abundancia de especies (Aliaume et al. 1990, Nagelkerken et al. 2001, Unsworth 2007). Un medio de mayor heterogeneidad proporciona un mayor número y variedad de refugios, además de una gama de alimentos más amplia la cual permite sostener un mayor número de individuos. De manera particular en la estación La Brea el pasto marino fue más frondoso y con mayor amplitud, y en este sitio se encontraron los mayores valores de riqueza, abundancia y biomasa, como era de esperarse.
Otro factor no descartable en la composición íctica de estos pastos marinos es la posible conectividad con otros ecosistemas, tal es el caso de La Brea la cual está muy cercana a un gran complejo arrecifal en un sector denominado El Toro, conexión que se evidencia por la continua presencia de especies características de zonas arrecifales como el sargento Abudefduf saxatilis, entre otros, lo cual puede variar e inclusive incrementar la estructura íctica del sistema (De Grado et al. 2000). La proximidad a arrecifes de coral u otros hábitats más complejos efectivamente pueden influenciar la distribución y abundancia de ciertas especies ícticas en los pastos marinos (Nagelkerken et al. 2002, Nagelkerken & Van der Velde 2004). Baelde (1990), señala que las praderas son usadas como áreas de forraje por especies de peces coralinos y zonas de cría para peces de manglares cercanos. Aunque esta conectividad generalmente se evidencia sólo en determinadas especies (Acosta et al. 2007).
Estos resultados son una valiosa información sobre el potencial de los recursos actuales de la región y con los cuales se pueden desarrollar mejores alternativas de conservación y manejo para la comunidad de peces costeros. Asimismo, se demuestra que las praderas de Thalassia testudinum, en la costa noroccidental del Golfo de Cariaco, sostienen una gran abundancia y diversidad de peces juveniles; las cuales funcionan como áreas de cría, refugio y alimentación. Por lo tanto, es fundamental y oportuna la protección de este tipo de hábitat para la conservación de la diversidad de peces marinos en la región nororiental de Venezuela.
Agradecimientos
A Luis Nicolás Suarez y a su familia, por la dedicación invaluable y el conocimiento brindado durante los muestreos. A José Acosta por su colaboración en el procesamiento de las muestras.
Referencias
Acero, A. 1980. Observaciones ecológicas de la ictiofauna de una pradera de Thalassia en la Bahía de Nenguange (Parque Nacional Tayrona, Colombia). Bol. Inst. Oceanogr. São Paulo 29: 5-8. [ Links ]
Acero, A. & J. Garzón-Ferreira. 1986. Peces de las Islas del Rosario y de San Bernardo (Colombia) II. Tres nuevos registros para el Caribe Sur y 16 más para la costa norte continental colombiana. An. Inst. Inv. Mar. Punta Betín 15: 3-29. [ Links ]
Acosta, A., C. Bartels, J. Colcoveresses & M. Greenwood. 2007. Fish assemblages in seagrass habitats of the Florida Keys, Florida: Spatial and temporal characteristics. Bull. Mar. Sci. 81: 1-19. [ Links ]
Aguilar, J. 2004. Coastal habitat connectivity of reef fishes from Southwestern Puerto Rico. Tesis de Doctorado en Ciencias Marinas, Universidad de Puerto Rico,
Mayagüez, Puerto Rico. [ Links ]
Aliaume, C., G. Lasserre & M. Louis. 1990. Organisation spatiale des peuplements ichtyologiques des herbies á Thalassia du Grand Cul-de-Sac Marin en Guadeloupe. Rev. Hydrobiol. Trop. 23: 231-250. [ Links ]
Allen, T., M. Jiménez & S. Villafranca. 2004. Abundancia y riqueza específica de la ictiofauna asociada con Thalassia testudinum en el Golfo de Cariaco, Venezuela. Rev. Biol. Trop. 52: 973-980. [ Links ]
Allen, T., M. Jiménez, B. Márquez & Y. Figueroa. 2007. Ictiofauna de tres praderas de Thalassia testudinum de la costa norte del Golfo de Santa Fé, Estado Sucre Venezuela. Bol. Inst. Oceanogr. Venezuela 46: 67-78. [ Links ]
Amezcua-Linares, F. & A. Yáñez-Arancibia. 1980. Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El hábitat y estructura de las comunidades de peces. An. Centro Cien. Mar. Limnol. Univ. Nac. Autón. México 7: 69-118. [ Links ]
Baelde, P. 1990. Difference in the structure of fish assemblages in Thalassia testudinum beds in Guadeloupe, French West Indies, and their ecological significance. Mar. Biol. 105: 163-173. [ Links ]
Boström, C. & E. Bonsdorff. 2000. Zoobenthic community establishment and habitat complexity-the importance of seagrass shoot-density, morphology and physical disturbance for faunal recruitment. Mar. Ecol. Progr. 205: 123-138. [ Links ]
Bouchon-Navarro, Y., C. Bouchon & M. Louis. 1992. L’ichtyofaune des herbiers de phanérogames marines de la Baie de Fort-de-France (Martinique, Antilles Francaises). Cybium 16: 307-330. [ Links ]
Cervigón, F. 1991. Los peces marinos de Venezuela. Vol. I. Fundación Científica Los Roques, Cromotip, Caracas, Venezuela. [ Links ]
Cervigón, F. 1993. Los peces marinos de Venezuela. Vol. II. Fundación Científica Los Roques, Cromotip, Caracas, Venezuela. [ Links ]
Cervigón, F. 1994. Los peces marinos de Venezuela. Vol. III. Fundación Científica Los Roques, ExLibris, Caracas, Venezuela. [ Links ]
Cervigón, F. 1996. Los peces marinos de Venezuela. Vol. IV. Fundación Científica Los Roques, ExLibris, Caracas, Venezuela. [ Links ]
De Grado, A., A. Bashirullah & A. Prieto. 2000. Variación espacio-temporal de la comunidad de peces en Laguna Grande de Obispo, Golfo de Cariaco, Estado Sucre, Venezuela. Acta Cient. Venez. 51: 96-103. [ Links ]
Den Hartog, C. & J. Kuo. 2006. Taxonomy and biogeography of seagrasses, p. 1-23. In A.W.D Larkum, R.J. Orth & C.M. Duarte (eds.). Seagrasses: Biology, Ecology and Conservation. Springer Netherlands, Dordrecht, Holanda. [ Links ]
Díaz, Y. 1997. Relaciones tróficas en la ictiofauna asociada a praderas de Thalassia testudinum en el Parque Nacional Morrocoy. Tesis de Grado, Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Gómez, A. 1987. Estructura de la taxocenosis de peces en praderas de Thalassia testudinum de la Bahía de Charagato, Isla de Cubagua, Venezuela. Bol. Inst. Oceanogr. Venezuela 26: 125-146. [ Links ]
González-Gándara, C., S. Trinidad-Martínez & V. Chávez- Morales. 2006. Peces ligados a Thalassia testudinum en el Arrecife Lobos, Veracruz, México: diversidad y abundancia. Rev. Biol. Trop. 54: 18-194. [ Links ]
Gullström, M. & Dahlberg, M. 2004. Fish community structure of seagrass meadows around Inhaca Island, southern Mozambique. Tesis de Doctorado, Uppsala University, Uppsala, Suecia. [ Links ]
Johnson, R.A. & D.W. Wichern. 1992. Applied multivariate statistical analysis. Prentice-Hall, Englewood Cliffs, Nueva Jersey, EEUU. [ Links ]
Krebs, C. 1989. Ecological Methodology: The Experimental Analysis of Distribution and Abundance. Harper & Row, Nueva York, EEUU. [ Links ]
López-Ordaz, A., M. Ortaz & J. Rodríguez-Quintal. 2009a. Trama trófica de una comunidad de peces en una pradera marina en el Caribe Venezolano. Rev. Biol. Trop. 57: 963-975. [ Links ]
López, M., L. Ruíz & A. Prieto. 2009b. Abundancia y riqueza de peces en dos praderas de Thalassia testudinum en la zona costera de Cumana, Estado Sucre, Venezuela. Bol. Centro Invest. Biol. 43: 29-46. [ Links ]
Martínez, L. 1971. Comparación de la fauna ictiológica de la Laguna Grande del Obispo y la Bahía de Mochima, Sucre, Venezuela. Trabajo para ascender a la categoría de Profesor Asociado, Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Menares, B. & J.I. Sepúlveda. 2005. Grupos recurrentes de peces y crustáceos demersales en la zona centro-sur de Chile. Inv. Mar. Valparaíso 33: 91-100. [ Links ]
Méndez, E. 1995. Ictiofauna de una pradera de Thalassia testudinum Saco del Golfo de Cariaco. Edo. Sucre. Venezuela: Análisis de comunidad. Trabajo para ascender a la categoría de Profesor Titular, Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Nagelkerken, I. & G. Van der Velde. 2004. Relative importance of interlinked mangroves and seagrass beds as feeding habitats for juvenile reef fish on a Caribbean Island. Mar. Ecol. Progr. 274: 153-159. [ Links ]
Nagelkerken, I., G. Van der Velde, M. Gorissen, G. Meijer, T. Van’t Hof & C. Den Hartog. 2000. Importance of Mangroves, Seagrass Beds and the hollow coral reef as a Nursery for Important Coral Reef Fishes, Using a Visual Census Technique. Estuar. Coast. Shelf Sci. 51: 31-44. [ Links ]
Nagelkerken, I., S. Kleijnen, T. Klop, R. van den Brand, E. Cocheret & G. Van der Velde. 2001. Dependence of Caribbean reef fishes on mangroves and seagrass beds as nursery habitats: a comparison of fish faunas between bays with and without mangroves/seagrass beds. Mar. Ecol. Progr. 214: 225-235. [ Links ]
Nagelkerken, I., C. Roberts, G. Van der Velde, M. Dorenbosch, M. Van Riel, E. Cocheret & P. Nienhuis. 2002. How important are mangroves and seagrass beds for coral-reef fish? The nursery hypothesis tested on an island scale. Mar. Ecol. Prog. 244: 299-305. [ Links ]
Pollard, D. 1984. A review of ecological studies on seagrass-fish communities, with particular reference to recent studies in Australia. Aquat. Bot. 18: 3-42. [ Links ]
Rosales-Casián, J. 2004. Composición, importancia y movimientos de los peces de Bahía de San Quintín, Baja California, México. Ciencias Marinas 30: 109-117. [ Links ]
Ruiz, L. 1992. Estructura de las comunidades de peces en dos localidades del Saco del Golfo de Cariaco, Venezuela. Trabajo para ascender a la categoría de Profesor Titular, Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
San Cristóbal, C. 1984. Estructura de la comunidad ictiológica de dos praderas de Thalassia, en la Bahía de Mochima, Estado Sucre. Tesis de Grado en Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Sanders, H. 1960. Benthic studies in Buzzards Bay. III. The structure of the soft bottom community. Limnol. Oceanogr. 5: 138-153. [ Links ]
Silva, F. 2009. Estructura de la comunidad íctica de una pradera de Thalassia en el Saco del Golfo de Cariaco, Estado Sucre, Venezuela. Tesis de Grado en Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Soberón-Chávez, G. 1984. Mecanismos de producción de las poblaciones de peces demersales de la plataforma continental del sur del Golfo de México: Variables de interacción ecológica. Tesis de Maestría en Ciencias del Mar. Instituto de Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México, Ciudad de México, México. [ Links ]
Unsworth, R. 2007. Aspects of the ecology of Indo-Pacific seagrass systems. Tesis de Doctorado, Essex University, Colchester, Inglaterra. [ Links ]
Valecillos, Y. 1993. Estructura ecológica de la comunidad de peces del sistema Chacopata-Bocaripo, Península de Araya, Edo. Sucre. Venezuela. Tesis de Grado en Biología, Universidad de Oriente, Cumaná, Venezuela. [ Links ]
Vargas-Maldonado, I. & A. Yáñez-Arancibia. 1987. Estructura de las comunidades de peces en pastos marinos (Thalassia testudinum) de la Laguna de Términos, Campeche, México. An. Centro Cien. Mar. Limnol. Univ. Nac. Autón. México 14: 181-196. [ Links ]
Victoria, P. & D. Gómez. 1984. Nuevos registros de peces para la isla de San Andrés (Mar Caribe de Colombia). An. Inst. Mar. Punta Betín 14: 115-132. [ Links ]
Villamizar, E. 1993. Evaluación de la comunidad de peces en praderas de fanerógamas marinas del Parque Nacional Archipiélago de los Roques. Tesis de Doctorado, Universidad Central de Venezuela, Dtto. Capital, Venezuela. [ Links ]
Wilhm, J. 1968. Use of biomass units in Shannon’s formula. Ecology 49: 153-156. [ Links ]
Zar, J. 1996. Biostatistical analysis. Prentice-Hall International, Englewood Cliffs, Nueva Jersey, EEUU. [ Links ]
*Correspondencia a: Luis Alejandro Ariza A. & José Gregorio Núñez P.: Postgrado en Ciencias Marinas, Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná, Venezuela; luisalejandroariza@gmail.com, jgnp31@gmail.com
Lilia Ruiz & Elizabeth Méndez de E.: Departamento de Biología, Escuela de Ciencias, Universidad de Oriente, Apartado postal 245, Cumaná, Venezuela; liliaruiz@cantv.net, ibaiondo2006@gmail.com
1. Postgrado en Ciencias Marinas, Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná, Venezuela; luisalejandroariza@gmail.com, jgnp31@gmail.com
2. Departamento de Biología, Escuela de Ciencias, Universidad de Oriente, Apartado postal 245, Cumaná, Venezuela; liliaruiz@cantv.net, ibaiondo2006@gmail.com
Recibido 07-IV-2011. Corregido 20-VIII-2011. Aceptado 26-IX-2011.


{kind=link}
{kind=link}
{kind=link}