










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.2 San José Jun. 2012
Araneofauna (Arachnida: Araneae) en cultivos de algodón (Gossypium hirsutum) transgénicos y convencionales en el norte de Santa Fe, Argentina
Arachnofauna (Araneae: Araneae) in transgenic and conventional cotton crops (Gossypium hirsutum) in the North of Santa Fe, Argentina
Arachnofauna (Araneae: Araneae) in transgenic and conventional cotton crops (Gossypium hirsutum) in the North of Santa Fe, Argentina
*Dirección para correspondencia:
Abstract
Spiders have considerable potential importance for their role as predators to some pests in agricultural systems. The composition of spiders in transgenic and conventional cotton at the Research Station of INTA Reconquista (Santa Fe) was studied during the 2005-2006 season. The experiment was a complete randomized block design with three replications and three treatments: transgenic Bt cotton (ALBt), conventional cotton without chemical control (ALCSC), and conventional cotton with chemical control (ALCCC). Weekly, spiders were collected using nets, vertical cloth and pitfall-traps. A total of 1 255 specimens (16 families, and 32 species) were collected. Seven families were found in all the treatments, mainly Thomisidae (n=1 051, 84.04%) and Araneidae (n=83, 6.64%). The Hunting spiders guild ambushers (n=1 053, 83.91%), “Orb weavers” (n=85, 6.77%) and “Stalkers” (n=53, 4.22%) were more abundant. There were no significant differences in the indexes diversity between treatments. Spiders were presented during the whole crop season, with peaks about flowering and boll maturity, with the highest abundance in ALBt. This work is part of the first set of data registered in Argentina about spider’s community in cotton crops. Rev. Biol. Trop. 60 (2): 611-623. Epub 2012 June 01.
Key words: spiders, diversity, Bt crop, agroecosystems, Reconquista.
Resumen
Las arañas tienen un valor potencial considerable por su rol depredador de insectos, estas son plagas de la agricultura. Durante la campana agrícola 2005/06, en INTA Reconquista, Santa Fe (Argentina) se estudio la composición de arañas presentes en cultivos de algodón transgénico y convencional, mediante un diseño experimental de bloques completos al azar, con tres repeticiones y tres tratamientos: algodón transgénico Bt (ALBt), algodón convencional sin control químico (ALCSC) y con control químico (ALCCC). Semanalmente, se capturaron arañas, con una red entomológica de arrastre, paño vertical de 1m y trampas de caída. Asimismo se recolectaron 1 255 ejemplares (16 familias y 32 especies). Siete familias se presentaron en los tres tratamientos, donde predomino Thomisidae (n=1 051, 84.04%) y Araneidae (n=83, 6.64%). El gremio cazadoras por emboscada (n=1 053, 83.91%), “Tejedoras de telas orbiculares” (n=85, 6.77%) y “Cazadoras al acecho” (n=53, 4.22%) fueron las mas abundantes. No hubo diferencias significativas en los índices de diversidad entre tratamientos. Las arañas se presentaron durante todo el ciclo del cultivo, con picos en las semanas de floración y madurez de las capsulas, además la mayor abundancia la encontramos en el ALBt. Este trabajo constituye el primer registro sobre la comunidad de arañas en cultivos de algodón para Argentina.
Palabras claves: arañas, diversidad, cultivo Bt, agroecosistemas, Reconquista.
La desaparición de especies expresa la falta de estrategias adecuadas de manejo y acciones para desarrollar planes apropiados para la conservación de las mismas. Las actividades antrópicas han reducido significativamente las áreas naturales y en consecuencia han favorecido la perdida de la biodiversidad. El conocimiento de la taxocenosis de especies, la obtención de datos de las comunidades biológicas, el estudio de las relaciones ecológicas y funcionales a nivel ecosistémico, ayudan a encarar y poner en marcha acciones destinadas a la conservación de la biodiversidad (Patrick et al. 1999).
Los estudios sobre la comunidad de arañas en los últimos anos han cobrado gran importancia, debido a sus características ecológicas y biológicas. Las arañas son abundantes en ambientes naturales y cultivados, y son consideradas uno de los grupos de macroinvertebrados depredadores dominantes en el medio terrestre (Patrick et al. 1999). En los agroecosistemas, se las considera enemigos naturales de insectos perjudiciales, pero al mismo tiempo sirven de alimento para otros depredadores (Whitcomb 1980).
En los últimos anos se han incrementado los estudios de la biota del neotropico referidos a las comunidades de arañas en bosques (Hofer 1990, Silva 1996, Silva & Coddington 1996, Flórez 1999, Avalos et al. 2007, Grismado 2007, Rubio et al. 2008), en areas naturales protegidas (Corronca & Abdala 1994, Rico-G et al. 2005, Zvedeniuck 2009, López Lezama 2010) y en diferentes cultivos, como los realizados en cultivos de arroz (Rodrigues et al. 2008), de eucaliptos (Rodrigues et al. 2010) y especialmente los estudios realizados en Argentina sobre aranas asociadas a cultivos de trigo, soja y alfalfa (Minervino 1996, Liljesthrom et al. 2002, Beltramo et al. 2006, Armendano 2008, Gonzalez et al. 2009, Armendano & Gonzalez 2010, Benamu Pino 2010). En otros países, se ha documentado en diversos trabajos la incidencia de las arañas como controladoras de diversas plagas en cultivos de algodón, y se demostró la gran abundancia de presas capturadas por los diferentes gremios de aranas (Hayes & Lockley 1990, Nyffeler et al. 1987, 1988, 1989, 1992, Nyffeler & Sterling 1994, Perez-Guerrero et al. 2009), mientras que en Argentina no existen hasta la actualidad, estudios referidos a esta temática.
La estructura del hábitat y su complejidad tiene una relación directa con la diversidad de las especies, los cuales determinan la presencia de diferentes gremios de arañas, al presentar diferentes requerimientos (Wise 1993, Altieri 1995, Pearce et al. 2003). Por lo tanto, la identificación de los gremios de arañas, son esenciales para los estudios de artrópodos en los agroecosistemas (Uetz et al. 1999).
El algodón es un cultivo anual con una compleja arquitectura y desarrollo de canopia, además presenta una gran riqueza y diversidad de fauna, en la cual se incluye a las arañas (Luczak 1979, Heitholt 1999). En los últimos anos han surgido nuevas variedades, geneticamente modificadas de algodón, por ejemplo la variedad transgenica de algodón Bt, utilizada como herramienta adicional, junto a nuevas tecnologías (aplicación de diferentes alternativas de cosecha, sistemas de siembras, etc.) (Paytas 2010), que ayudan al crecimiento de la producción algodonera. Gomez Galvis & Florez Daza (2005) evaluaron el efecto de la variedad de algodón transgénico sobre la araneofauna, debido a que su aplicación esta muy cuestionada por su posible impacto en la biodiversidad.
El algodón es uno de los cultivos más importantes desde el punto de vista económico para Argentina y puesto que, no existen estudios referidos a la comunidad de arañas en este cultivo, el objetivo del presente trabajo es conocer la composición, diversidad, riqueza y abundancia de arañas, y los gremios presentes en cultivos de algodón transgénico y convencional con control químico y sin control químico en el norte de la provincia de Santa Fe, Argentina.
Materiales y métodos
Área de estudio: El trabajo se llevo a cabo durante la campana de algodón 2005- 2006 (noviembre a abril) en el campo de la Estación Experimental Agropecuaria de INTA Reconquista (29°11’ S - 59°52’ W), provincia de Santa Fe. La región pertenece al área Fitogeografica de bosques y esteros del Chaco Húmedo, con temperaturas medias anuales, superiores a los 20oC y precipitaciones que superan las 1 100-1 200mm anuales (Pereyra 2003).
El cultivo de algodón fue trabajado mediante labranza convencional (Rojas 2001). El diseño experimental utilizado fue de bloques completos al azar, con tres tratamientos y tres repeticiones (Cantatore de Frank 1980). Cada parcela fue de 30m de ancho x 30m de largo y estaba rodeada por cuatro surcos de sorgo a 2.8m y separadas entre si por calles de 10m libres de vegetación. La distancia entre los surcos del algodón dentro de cada parcela fue de 0.90m. El cultivo estuvo rodeado por zonas adyacentes de cultivos de soja y plantaciones de eucaliptus. Los tratamientos fueron: 1) algodón transgénico Bt (ALBt), 2) algodón convencional sin control químico (ALCSC) y 3) algodón convencional con control químico ALCCC). Los insecticidas aplicados al ALCCC, fueron: Cipermetrina EC 25%, 150cc/ha, Endosulfan EC 35%, 2lt/ha, Spinosad SC 48%, 40cc/ha y Novaluron SC 10%, 100cc/ha a los 30, 45 y 70 días después de la emergencia. Todos las parcelas el 15/02/06 (estado de madurez de capsulas) recibieron una aplicación de regulador de crecimiento, Cloromecuato, 150cc/ha.
Muestreo de arañas: Las capturas de arañas se realizaron semanalmente, a los 30 días después de la emergencia del cultivo, mediante tres técnicas de muestreos: 1) Pano vertical de un metro (Sosa et al. 1998) con la cual: se seleccionaron al azar cinco metros contiguos en cada parcela; 2) Red entomológica de arrastre (Kogan & Pitre 1980) en donde: se realizaron 50 golpes de red al azar en cada parcela; 3) Trampas de caída (pitfall-traps) (Melbourne 1999, Adis 2002) las cuales se dispusieron al azar, cuatro por parcela, con solución salina (sal (kg): agua (l) en proporción 1:8, con gotas de detergente) y se mantuvieron activas durante 48hr. Por parcela se registro el número de arañas por cinco metros de paño, por 50 golpes de red y por cuatro trampas de caída.
Las arañas recolectadas fueron fijadas con alcohol al 70% e identificadas mediante claves sistemáticas (Ramirez 1999, Dippenaar-Schoeman & Jocque 1997) y consulta de material de referencia de la colección existente en el Museo de Ciencias Naturales “Florentino Ameghino” (Provincia de Santa Fe). Cuando no fue posible la identificación especifica, los especímenes fueron separados en morfoespecie. Asimismo, se determino el sexo y estado de desarrollo de cada individuo.
El material recolectado fue donado a la Cátedra de Diversidad Animal I, Facultad de Humanidades y Ciencias, Universidad Nacional del Litoral y al Museo Provincial de Ciencias Naturales “Florentino Ameghino” de Santa Fe (Argentina).
Para el calculo de los índices en cada tratamiento, se considero la suma de arañas capturadas, mediante el programa PAST, versión 1.83 (Hammer et al. 2001) se calculo la riqueza de especies (S) y la diversidad mediante los índices de Shannon-Wienner (H’), de Simpson (λ) y de Equidad de Pielou (J’). También, se estimo el indice de similitud por presencia-ausencia de Jaccard (Moreno 2001). Las especies de arañas fueron agrupadas en gremios a partir de la clasificación propuesta por Uetz et al. (1999).
Los datos de abundancia, riqueza e índice de diversidad fueron analizados mediante un análisis de varianza (ANOVA one-way), donde se ingresaron las arañas identificadas a nivel de familia. Para la comparación de medias se utilizo el Test de Tukey (α=0.05) del software InfoStat/Profesional, versión: 2010 (Di Rienzo et al. 2010). Ademas se comparo la abundancia de aranas en cada tratamiento a lo largo del desarrollo del cultivo.
Resultados
Composición de la comunidad de arañas: Se recolecto un total de 1 255 arañas en los tres tratamientos. El algodón Bt fue el que presento el mayor numero de arañas (659), seguido por el ALCSC (354) y por ultimo el ALCCC (242).
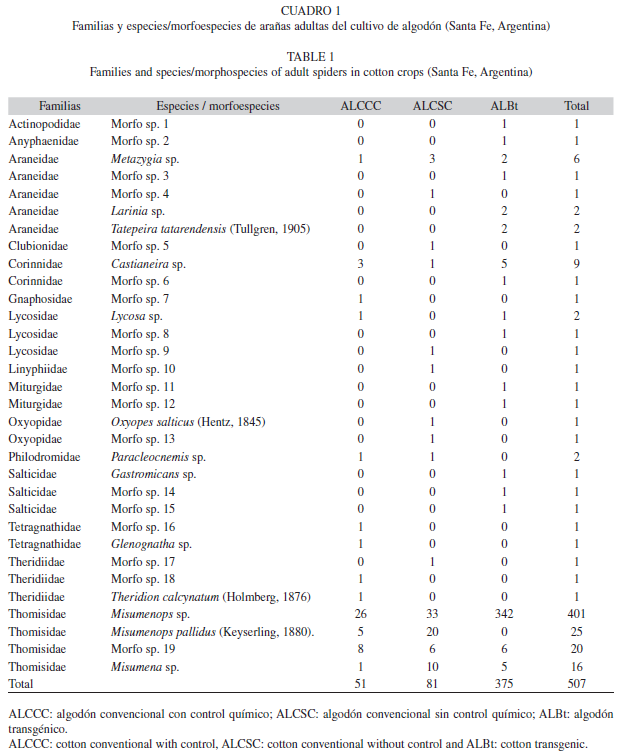
De tal manera que en total se identificaron 16 familias (que incluye juveniles y adultos) capturadas en los tres tratamientos, una de las cuales, Actinopodidae, pertenece al suborden Mygalomorphae, y las restantes 15 al suborden Araneomorphae. En los tres tratamientos la familia mas abundante fue Thomisidae (1 0 1 individuos; 83.74% del total de arañas recolectadas), seguida por Araneidae (83 individuos; 6.64%), Salticidae (34 individuos; 2.72%), Lycosidae (21 individuos; 1.68%), Oxyopidae (19 individuos; 1.52%), Corinnidae (10 individuos; 0.8%) y Theridiidae (9 individuos; 0.72%). Actinopodidae y Anyphaenidae (1 individuo; 0.08%) se registraron únicamente en el ALBt, Clubionidae y Linyphiidae (1 individuo; 0.08%) en el ALCSC, y Tetragnathidae (2 individuos; 0.16%) en el ALCCC. Las familias Gnaphosidae y Philodromidae, solo estuvieron presentes en el ALCSC y en el ALCCC, mientras que la familia Miturgidae y Sparassidae solo se detectaron en el ALCCC y ALBt.
De los individuos capturados, el 60% (n=748) correspondió a individuos juveniles y el resto (n=507) a individuos adultos, entre los que se identificaron 32 especies (Cuadro 1); y cabe destacar que la mas abundante en los tres tratamientos fue Misumenops sp. (401 individuos; 79.09%). El 84.63% de los individuos adultos correspondió a machos y el 15.37% a hembras. La especie mas abundante entre los machos fue Misumenops sp. y entre las hembras Misumenops pallidus (Keyserling, 1880).
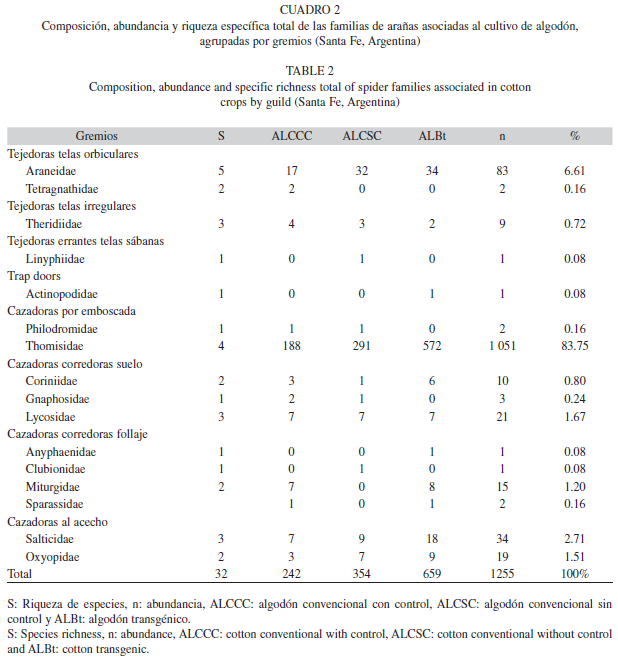
Composición de gremios: Las familias recolectadas en los tres tratamientos, en donde se toma en cuenta el total de aranas capturadas entre juveniles y adultos, se agruparon en ocho gremios (Cuadro 2). El mas representativo fue el de “Cazadoras por emboscada” con 1 053 individuos, siendo la familia Thomisidae la mas abundante. Seguido por el gremio “Tejedoras de telas orbiculares” con 85 individuos y “Cazadoras al acecho” con 53 individuos. En menor cantidad se presento el gremio “Cazadoras corredoras de suelo”, “Cazadoras corredoras follaje” y “Tejedoras telas irregulares”, con 34, 19 y nueve individuos respectivamente. El gremio exclusivo del ALCSC fue el de “Tejedoras errantes de telas de sabanas”, mientras que “Trap Doors” se presento solamente en el ALBt. Para los tres tipos de tratamientos, las abundancias de cada gremio fueron analizadas, donde unicamente el gremio “Cazadoras al acecho” difiere significativamente entre el ALBt y ALCSC (F=3.41, p=0.0345, gl=2), mientras que los demas gremios no presentaron diferencias (p≥0.05).
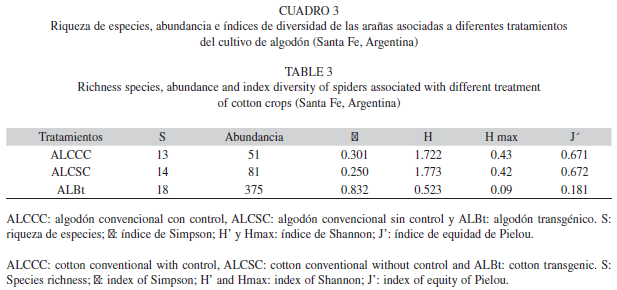
Composición específica e índices de diversidad: Los valores de los índices de diversidad de Shannon, de Simpson y de Equidad para los tres tratamientos estudiados se muestran en el Cuadro 3. El mayor valor del índice de diversidad (H’), correspondió al ALCSC (H’=1.773), seguido por ALCCC (H’=1.722) y por ALBt (H’=0.523), mientras que la mayor riqueza especifica correspondió al ALBt (S=18), seguido por el ALCSC (S=14) y por el ALCCC (S=13). A partir del análisis de varianza, la diversidad de familias analizadas mediante el Test de Tukey (α=0.05) no mostro diferencias significativas entre los tratamientos (λ: F=1.79, p=0.2791, gl=2; H’: F=4.14, p=0.1062, gl=2). Entonces al realizar el análisis de la riqueza de especies, el ALBt presento mayor riqueza que los otros dos tratamientos, diferenciándose significativamente (F=38,8, p=0.0024, gl=2).
Con respecto a la estructura de la comunidad de arañas, se puede observar que el índice de equidad de Pielou fue mayor en el ALCSC (J’=0.672), donde mas de la mitad de las especies están igualmente representadas. Luego sigue el ALCCC (J’=0.671) y por ultimo el ALBt (J’=0.181) (Cuadro 3). Sin embargo, el análisis de varianza entre los tratamientos no presentaron diferencias significativas (J’: F=1.15, p=0.404, gl=2).
Ahora bien, con lo que se refiere a la similitud de especies, el índice de Jaccard demostró que existe un 35% de especies compartidas entre los tratamientos ALCCC y ALCSC, seguido de un 24% entre ALCCC y ALBt y en menor proporción, 19% entre ALBt y ALCSC.
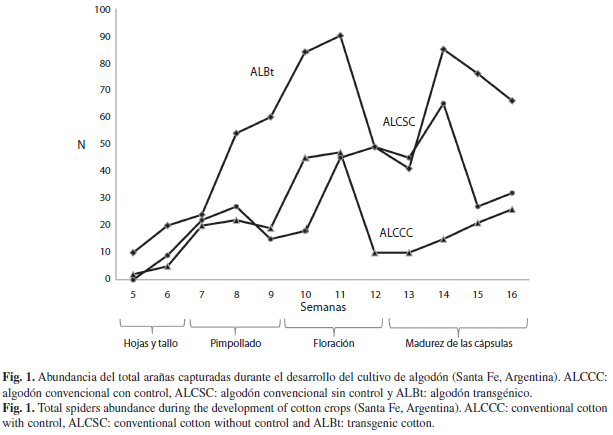
Análisis temporal: Durante todo el desarrollo del cultivo se encontraron arañas en los tres tratamientos. El mayor pico de arañas se registro en el estado de floración, mes de febrero (semana 11) (Fig. 1), el cual corresponde con pico mas alto de ALBt (n=90), donde dominaron las familias Thomisidae (4.91%) y Araneidae (0.85%). Las parcelas ALCSC y ALCCC alcanzaron una abundancia igual a 45 y 47 respectivamente, y Thomisidae fue la familia predominante (4.16% y 3.40%, respectivamente). Se observo un segundo pico de abundancia de la población de arañas que coincidió con el estado de madurez de las capsulas, mes de marzo (semana 14), en el ALBt (n=85) y en el ALCSC (n=65), mientras que en el ALCCC no se registro un segundo aumento en el numero de arañas. Nuevamente las familias Thomisidae (4.44%) y Araneidae (1.32%) fueron las dominantes. El registro del menor numero de arañas correspondió a la semana cinco en los tres tratamientos (n=9) y en la semana 13 del ALCCC (n=13).
El análisis de varianza para la abundancia de arañas en los tres tratamientos, demostró que existieron diferencias significativas en seis semanas del desarrollo del cultivo. En las semanas 5 y 9, el ALCSC obtuvo una abundancia menor al 1% del total, el cual difiere del ALBt y ALCCC (Semana 5: F=9.75, p=0.029, gl=2; semana 9: F=7.82, p=0.0415, gl=2). En la semana 10 el ALBt y ALCCC se diferenciaron del ALCSC (F=17.06, p= 0.011, gl=2), donde este ultimo presento una menor cantidad de arañas (n=18). En la semana 11 el ALBt se diferencio de los otros dos tratamientos (F=15.47, p=0.0131, gl=2). En la semana 12, el ALCSC y ALBt, se diferenciaron del ALCCC, ya que este ultimo presento una menor abundancia (n=10). Por ultimo, en la semana 13 se diferencio el ALCSC del ALCCC, pero no así el ALBt de los otros dos tratamientos (F=7.43, p=0.045, gl=2).
Discusión
Las familias de arañas registradas para los cultivos de algodón estudiados en el presente trabajo, representan el 25% de las familias de arañas citadas para Argentina (Pikelin & Schiapelli 1963, Platnick 2011), resultados similares con los planteados por Liljesthrom et al. (2002), Beltramo et al. (2006), Benamu Pino (2010), Armendano & González (2010). Esto es esperable por tratarse de ambientes perturbados en los cuales se reduce drásticamente la diversidad de la vegetación y por consiguiente disminuye la biodiversidad faunística, donde están incluidas las arañas (Altieri 1995).
De las 16 familias registradas, cuatro representaron el 95% del total capturado. Estos resultados son coincidentes con lo propuesto por Young & Edwards (1990), donde demostraron que solo cinco familias de arañas constituyen el 61% del total de la comunidad de arañas en cultivos norteamericanos.
La familia Thomisidae fue la que contribuyo con el mayor numero de arañas, la cual alcanzo el 84.08 % del total, seguida por las familias Araneidae, Salticidae y Lycosidae, aunque en proporciones mas bajas. El resto de las familias no supero el 2%. Al comparar nuestros resultados con otros estudios realizados sobre cultivos de algodón se observan otras familias dominantes. En Texas, Egipto y Australia, Mansour (1987), Dean & Sterling (1992) y Whitehouse et al. (2005) encontraron que las familias Oxyopidae, Tetragnathidae y Clubionidae fueron las mas representativas de los cultivos de algodón convencional. De igual forma, Gómez Galvis & Flórez Daza (2005), en un estudio realizado en Colombia sobre cultivos de algodón transgénico y convencional, observaron que Theridiidae y Linyphiidae fueron las familias que contribuyeron con mayor numero, mientras que Pérez-Guerrero et al. (2009), en España, sobre cultivos de algodón ecológico, señalan como familias mas abundantes a Miturgidae y Salticidae. Asimismo coinciden con nuestros resultados, Minervino (1996) y Liljesthrom et al. (2002) en cultivos de soja convencional y Armendano & Gonzalez (2010) en cultivos de alfalfa, y mostraron que Thomisidae fue la familia dominante representando el 50% del total de las arañas presentes. La gran abundancia para Thomisidae demuestra la capacidad de esta familia para habitar sitios perturbados, como también adaptarse a las distintas condiciones naturales, donde se comportaría como un importante integrante del grupo depredador generalista en el cultivo de algodón. En cambio, estudios realizados en cultivos de soja transgénica (Benamu Pino 2010) realizados en la provincia de Buenos Aires, Argentina, las familias Hahniidae y Lycosidae predominaron con un 40% y 26% respectivamente y en cultivos de arroz en Rio Grande do Sul, Brasil, fueron mas representativas Anyphaenidae, Oxyopidae y Tetragnathidae (Rodrigues et al. 2008).
En los tres tratamientos estudiados la especie mas abundante fue Misumenops sp. Esto no es coincidente con lo observado en otros trabajos realizados en cultivos de algodón. Gómez Galvis & Flórez Daza (2005) registraron para Colombia Erigoninae sp. 1, Theridion sp. 1 y Theridula sp. 2 como las principales especies del cultivo de algodón convencional y transgénico; Dean & Sterling (1992) observaron que Oxyopes salticus (Hentz, 1845) contribuyo mayoritariamente en los cultivos de algodón convencional de Texas; Mansour (1987) para algodón convencional y Pérez Guerrero et al. (2009) para algodón ecológico, registraron para Israel y España respectivamente, la especie Cheiracanthium mildei (L. Kock 1864) como la mas representativa. Al comparar las especies obtenidas en los cultivos de algodón estudiados con otros cultivos, se observan resultados coincidentes con los nuestros. Minervino (1996) registro sobre cultivos de soja convencional, que M. pallidus contribuyo en un 42.3% y Campos et al. (1999) en cultivos de sorgo de Brasil, estimaron que la especie M. pallidus (25.6%) fue la mas abundante, lo que es coincidente con lo manifestado por Liljesthrom et al. (2002) y Armendano & Gonzalez (2010) para cultivos de soja convencional y alfalfa de Argentina. El caso contrario se presenta en Benamu Pino (2010) que registro a las especies Antistea sp. (Hahniidae) y Diapontia sp. (Lycosidae) como las mas abundantes para cultivos de soja transgénica y Rodrigues et al. (2008) en cultivos de arroz sostienen que las especies predominantes fueron: Alpaida veniliae (Keyserling, 1865), Tetragnatha nitens (Audouin, 1826), Ashtabula sp. y Tetragnatha aff. jaculator (Tullgren, 1910) datos no coincidentes con lo hallado en el presente trabajo.
El alto numero de ejemplares juveniles obtenidos (n=753) coincide con lo registrado en otros trabajos de cultivos agrícolas (Liljesthrom et al. 2002, Gómez Galvis & Flórez Daza 2005, Rodrigues et al. 2008, Armendano & Gonzalez 2010, Benamu Pino 2010) y en trabajos realizados en áreas naturales (Rubio et al. 2008, Avalos et al. 2009), donde mas del 60% de los ejemplares son inmaduros. Al analizar las proporciones de juveniles y adultos en trabajos de ecología de comunidades hace suponer una constante colonización del cultivo desde las zonas adyacentes (Minervino 1996, Beltramo et al. 2006, Rodrigues et al. 2008).
En el presente estudio predominaron las especies cazadoras por emboscadas, en donde la familia Thomisidae fue la mas abundante en los tres tratamientos, seguidas por las tejedoras de telas orbiculares y cazadoras al acecho. Este predominio es semejante a lo observado por Minervino (1996) y Liljesthrom et al. (2002) en cultivos de soja, por Armendano & González (2010) en cultivos de alfalfa y a lo propuesto por Uetz et al. (1999), donde la comunidad de arañas en agroecosistemas se caracteriza por la presencia de especies cazadoras por emboscada, seguidas por cazadoras corredoras de suelo, corredoras sobre la vegetación y tejedoras tipo sabanas y orbiculares. El predominio de las cazadoras por emboscada tendría una justificación al momento de considerar su incidencia sobre larvas de lepidopteros, causantes de grandes danos en el cultivo. En cambio el gremio mejor representado en cultivos de algodón convencional y transgénico de Colombia (Gómez Galvis & Flórez Daza 2005) fueron las tejedoras en sabana, determinado por la extraordinaria abundancia de la familia Linyphiidae, seguido por las tejedoras de telas orbiculares, tejedoras de telas irregulares, tejedoras en embudo, errantes en suelo, errantes en follaje, cazadoras al acecho y cazadoras en emboscada.
Desde el punto de vista temporal la mayor abundancia de arañas se observo en verano, en el cual alcanzo su pico mas alto en la floración (febrero) y maduración de las capsulas (marzo). Estos estados del cultivo (estado reproductivo) corresponden a la llegada de insectos, donde las arañas dispondrían de una importante fuente de alimento. En la semana 12, entre la floración y la maduración de las capsulas, se observa una brusca disminución en la densidad de las arañas, la que estaría relacionada con cuestiones climáticas (periodo de sequia y altas temperaturas seguidas de intensas lluvias), que coinciden con lo sugerido por Whitmore et al. (2002) respecto a que diversos factores afectan la estabilidad ambiental y consecuentemente la diversidad de especies en un hábitat determinado. De igual modo, Rodrigues et al. (2008) sostienen una correlación significativa entre factores abióticos (temperatura y precipitaciones) y la abundancia y riqueza especifica de arañas en cultivos de arroz en Brasil. Otro factor a tener en cuenta a la hora de analizar esta disminución y que podría estar influenciando la abundancia de la población de arañas, es la aplicación del regulador de crecimiento, practica que se realiza habitualmente antes de la formación de capsulas en el norte de Santa Fe.
La mayor diversidad correspondió al ALCSC, mientras que la mayor riqueza especifica correspondió al ALBt. No obstante, la diversidad analizada en los tres tratamientos no mostro diferencias significativas, pero si la riqueza del ALBt, donde se diferencio de los otros tratamientos. Por lo tanto, el algodón transgénico, a nivel de diversidad no se diferencio del algodón convencional, datos similares a lo hallado por Gómez Galvis y Flórez Daza (2005). Esto permite estimar que el algodón transgénico no afecta a la población de arañas, alberga significativamente una mayor riqueza de familias y es una herramienta efectiva para incluirlo dentro del control biológico de plagas de importancia agrícola.
El algodón transgénico presento la mayor abundancia de arañas a lo largo del desarrollo del cultivo y el algodón tratado con agroquímicos la menor. Esto concuerda con lo que postulan algunos autores donde la aplicación de insecticidas ejerce un efecto negativo sobre la población de arañas (Mansour 1987, Minervino 1996, Patrick et al. 1999, Pérez-Guerreo et al. 2009 & Benamu Pino 2010). Novillo et al. (1999) y Duran et al. (2000) sostienen que la lucha contra las plagas en cultivos de algodón se basa en el uso de insecticidas con las consecuencias de aparición de resistencia, contaminación ambiental y disminución de la población de enemigos naturales.
El presente trabajo constituye el primer registro sobre la comunidad de arañas en cultivos de algodón para Argentina. El conocimiento de la biodiversidad es un principio fundamental en la agricultura sustentable. Los resultados arribados demuestran la necesidad de incrementar los conocimientos sobre el rol de la comunidad de arañas en diferentes cultivos y la importancia de lograr practicas de manejo adecuadas (ej. Manejo en el uso de insecticidas) si se quiere alcanzar la autorregulación biótica de los agroecosistemas.
Agradecimientos
A la Estación Experimental Agropecuaria INTA Reconquista y al personal auxiliar del Laboratorio de Entomología por su colaboración en los muestreos a campo. A Daniela Vitti, Sebastián Zuil y Marcelo Paytas por los aportes brindados. Al personal técnico del Centro de Estudios Parasicológicos y de Vectores (CEPAVE) por su colaboración.
Referencias
Adis, J. 2002. Recommended sampling techniques, p. 555-576. In. J. Adis (ed.). Amazonian Arachnida and Myriapoda. Pensoft, Moscu, Rusia. [ Links ]
Altieri, M.A. 1995. Agroecology: the Science of Sustainable Agriculture. Westview, Boulder, Colorado, EEUU. [ Links ]
Armendano, A. 2008. Estudio de la aracneofauna presente en agroecosistemas de importancia económica (Trigo y alfalfa). Tesis de Doctorado, Universidad Nacional de La Plata, Buenos Aires, Argentina. [ Links ]
Armendano, A. & A. González. 2010. Comunidad de arañas (Arachnida, Araneae) del cultivo de alfalfa (Medicago sativa) en Buenos Aires, Argentina. Rev. Biol. Trop. 58: 747-757. [ Links ]
Avalos, G., G.D. Rubio, M.E. Bar & A. González. 2007. Arañas (Arachnida, Araneae) asociadas a dos bosques degradados del Chaco húmedo en Corrientes, Argentina. Rev. Biol. Trop. 55: 899-909. [ Links ]
Avalos, G., M.P. Damborsky, M.E. Bar, E.B. Oscherov & E. Porcel. 2009. Composición de la fauna de Araneae (Arachnida) de la Reserva provincial Ibera, Corrientes, Argentina. Rev. Biol. Trop. 57: 339-351. [ Links ]
Beltramo, J., I. Bertolaccini & A. González. 2006. Spiders of soybean crops in Santa Fe province, Argentina: influence of surrounding spontaneous vegetation on lot colonization. Bras. J. Biol. 66: 29-41. [ Links ]
Benamu Pino, M.A. 2010. Composición y estructura de la comunidad de arañas en el sistema de cultivo de soja transgénica. Tesis de Doctorado, Universidad Nacional de La Plata, Buenos Aires, Argentina. [ Links ]
Cantatore de Frank, N.M. 1980. Manual de estadística aplicada. Hemisferio Sur, Buenos Aires, Argentina. [ Links ]
Campos, A.R., E.B. Filho, F.M. Lara & I.M.P. Rinaldi. 1999. Composicao da Artropodofauna Entomofoga Associada a Diferentes Genotipos de Sorgo Granifero no Cerrado do Sudeste do Brasil. An. Soc. Entomol. Brasil 28: 703-714. [ Links ]
Corronca, J.A. & C.S. Abdala. 1994. La fauna araneologica de la Reserva Ecologica “El Bagual”, Formosa, Argentina. Aracnologia Supl. 9: 1-6. [ Links ]
Dean, D.A. & W.L. Sterling 1992. Comparison of sampling methods to predict phenology of predaceous arthropods in a cotton agroecosystem. Miscellaneous Public. 1: 1-13. [ Links ]
Dippenaar-Schoeman, A.S. & R. Jocque. 1997. African Spiders: An Identification Manual. Plant Protection Research Institute Handbook N° 9. [ Links ]
Di Rienzo, J.A., F. Casanoves, M.G. Balzarini, L. González, M. Tablada & C.W. Robledo. 2010. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. (también disponible en línea: www.infostat. com.ar). [ Links ]
Duran, J.M., M. Alvarado, E. Ortiz, A. De La Rosa, J.A. Ruiz, A. Sánchez & A. Serrano. 2000. Contribución al conocimiento de Earias insulana (Boisduval, 1833) (Lepidoptera, Noctuidae), la oruga espinosa del algodonero, en Andalucia occidental. Bol. San. Veg. Plagas 26: 215-228. [ Links ]
Florez, E. 1999. Estructura y composición de una comunidad de aranas (Araneae) en un bosque muy seco tropical de Colombia. Bol. Entomol. Venez. 14: 37-51. [ Links ]
Gomez Galvis, L. & E. Flórez Daza. 2005. Estudio comparativo de las comunidades de Arañas (Araneae) en cultivares de algodón convencional y transgénico en el Departamento del Tolima, Colombia. Acta Biol. Colombiana 10: 79. [ Links ]
González, A., G. Liljesthrom, E. Minervino, D. Castro, S. Gonzalez & A. Armendano. 2009. Predation by Misumenops pallidus (Araneae: Thomisidae) on insect pests of soybean cultures in Buenos Aires Province, Argentina. J. Arachnol. 37: 282-286. [ Links ]
Grismado, C. 2007. Comunidades de aranas de la Reserva Natural Otamendi, Provincia de Buenos Aires. Riqueza especifica y Diversidad. Departamento de Ciencias Biológicas, Universidad CAECE. Buenos Aires, Argentina. [ Links ]
Hammer, O., D.A.T. Harper & P.D. Ryan. 2001. PAST: Paleontological Statistics software package for education and data analysis. Paleontologia Electronica 4: 1-9. (también disponible en línea: http://palaeoelectronica.org/2001_1/past/issue1_01.htm). [ Links ]
Hayes, J.L. & T.C. Lockley. 1990. Prey and Nocturnal activity of wolf spiders (Araneae: Lycosidae) in cotton Fields in the Delta Region of Mississippi. Environ. Entomol. 5: 1512-1518. [ Links ]
Heitholt, J.J. 1999. Cotton: Factors associated with assimilation capacity flower production, boll set, and yield, p. 235-269. In L.D. Smith & C. Hamel (eds.). Crop yield, Physiology and processes. Stoneville, EEUU. [ Links ]
Hofer, H. 1990. The spider community (Araneae) of a central Amazonian blackwater inundation forest (Igapo). Acta Zoologica Fennica 190: 173-179. [ Links ]
Kogan, M. & H.N. Pitre. 1980. General Sampling methods for aboveground population of soybean arthropods, p. 30-60. In M. Kogan & D.E. Herzag (eds.). Sampling methods in Soy bean Entomology. Springer, Nueva York, EEUU. [ Links ]
Liljesthrom, G., E. Minervino, D. Castro & A. González. 2002. La comunidad de arañas del cultivo de soja en la provincia de Buenos Aires, Argentina. Neotrop. Entomol. 31: 197-210. [ Links ]
López Lezama, D.M. 2010. Biodiversidad de Theridiidae (Arachnida: Araneae) de la Reserva Natural e Histórica Isla Martin García, Buenos Aires, Argentina. Facultad de Ciencias, Programa Biología, IBAGUE, Universidad Del Tolima, Colombia. [ Links ]
Luczak, J. 1979. Spiders in agrocoenoses. Pol. Ecol. Stud. 5: 151-200. [ Links ]
Mansour, F. 1987. Spiders in sprayed and unsprayed cotton fields in Israel, their interactions with cotton pests and their importance as predators of the Egyptian cotton leaf worm, Spodoptera littoralis. Phytoparasitica 15: 31-41. [ Links ]
Melbourne, B.A. 1999. Bias in the effect of habitat structure on pitfall traps: An experimental evaluation. Australian J. Ecol. 24: 228-239. [ Links ]
Minervino, E. 1996. Estudio biológico y ecobiologico de arañas depredadoras de plagas de soja. Tesis de Docotorado, Universidad Nacional de La Plata, Buenos Aires, Argentina. [ Links ]
Moreno, C.E. 2001. Métodos para medir la biodiversidad. M & T. Manuales y Tesis SEA. Vol. 1. Zaragoza, España. [ Links ]
Novillo, C., J. Soto & J. Costa. 1999. Resultados en España con variedades de algodón, protegidas genéticamente contra las orugas de las capsulas. Bol. San. Veg. Plagas 25: 383-393. [ Links ]
Nyffeler, M., D.A. Dean & W.L. Sterling. 1987. Predation by green lynx spider, Peucetia viridans (Araneae: Oxyopidae), inhabiting cotton and woolly croton plants in east Texas. Environ. Entomol. 16: 355-359. [ Links ]
Nyffeler, M., D.A. Dean & W.L. Sterling. 1988. The southern black widow spider, Latrodectus mactans (Araneae, Theridiidae), as a predator of the red imported fire ant, Solenopsis invicta (Hymenoptera, Formicidae), in Texas cotton fields. J. Appl. Entomol. 106: 52-57. [ Links ]
Nyffeler, M., D.A. Dean & W.L. Sterling. 1989. Prey selection and predatory importance of orb-weaving spiders (Araneae: Araneidae, Uloboridae) in Texas cotton. Environ. Entomol. 18: 373-380. [ Links ]
Nyffeler, M., D.A. Dean & W.L. Sterling. 1992. Impact of the striped lynx spider (Araneae: Oxyopidae) and other natural enemies on the cotton fleahopper (Hemiptera: Miridae) in Texas cotton. Environ. Entomol. 21: 1178-1188. [ Links ]
Nyffeler, M. & W.L. Sterling. 1994. Comparison of the feeding niche of polyphagous insectivores (Araneae) in a Texas cotton plantation: estimates of niche breadth and overlap. Environ. Entomol. 23: 1294-1303. [ Links ]
Patrick, M., C. Alain & Y. Frederic. 1999. Spiders (Araneae) useful for pest limitation and bioindication. Agric. Ecosyst. Environ. 74: 229-273. [ Links ]
Paytas, M. 2010. Improving cotton yield under water limiting conditions in Argentina. Update on cotton production research 2: 17-18. [ Links ]
Pearce, J.L., L.A. Venier, G. Eccles, J. Pedlar & D. McKenney. 2003. Influence of habitat and microhabitat on epigeal spider (Araneae) assemblages in four stand types. Biodivers. Conserv. 13: 1305-1334. [ Links ]
Pereyra, F. 2003. Ecoregiones de la Argentina. Servicio Geológico Minero Argentino, Buenos Aires, Argentina. [ Links ]
Pérez-Guerrero, S., R. Tamajon, H.K. Aldebis & E. Vargas- Osuna. 2009. Comunidad de arañas en cultivos de algodón ecológico en el sur de España. Rev. Colom. Entomol. 35: 168-172. [ Links ]
Pikelin, B.S.G. de & R.D. Schiapelli. 1963. Llave para la determinación de las familias de arañas argentinas. Physis 24: 43-72. [ Links ]
Platnick, N.I. 2011. The World Spider Catalog, Version 10.0. American Museum of Natural History (Consultado: 15 enero 2011, http://research.amnh.org/iz/ spiders/catalog). [ Links ]
Ramírez, M.J. 1999. Clave para el Orden Araneae, p. 39-59. In F.A. Crespo, M.S. Iglesias, A.A. Ojanguren, M.J. Ramírez & A.C. Valverde. (eds.). El ABC en la determinación de artrópodos. Claves para especímenes presentes en la Argentina I. CCC Educando, Buenos Aires, Argentina. [ Links ]
Rico-G, A., J. Beltrán., A. Álvarez & D.E. Flórez. 2005. Diversidad de arañas (Arachnida: Araneae) en el Parque Nacional Natural Isla Gorgona, Pacifico Colombiano. Biota Neotropica 1 A: 1-12. [ Links ]
Rodrigues, E.N.L., M.J. Mendonca & R. Ott. 2008. Fauna de aranhas (Arachnida, Araneae) em diferentes estagios do cultivo do arroz irrigado em Cachoeirinha, RS, Brasil. Iheringia, Ser. Zool. Porto Alegre 98: 362-371. [ Links ]
Rodrigues, E.N.L., M.J. Mendonca, J.L.O. Rosado & A.E. Loeck. 2010. Soil spiders in differing environments: Eucalyptus plantations and grasslands in the Pampa biome, southern Brazil. Rev. Colomb. Entomol. 2: 277-284. [ Links ]
Rojas, L. 2001. La labranza mínima como practica de producción sostenible en granos básicos. Agron. Mesoamericana 12: 209-212. [ Links ]
Rubio, G.D., J.A. Corronca & M.P. Damborsky. 2008. Do spider diversity and assemblages change in different contiguous habitats? A case study in the protected habitats of the Humid Chaco ecoregion, north-east Argentina. Environ. Entomol. 37: 419-430. [ Links ]
Silva, D. 1996. Species composition and community structure of Peruvian rainforest spiders: a case study from a seasonally inundated forest alond the Samiria river. Revue Suisse de Zoologie 597-610. [ Links ]
Silva, D. & J.A. Coddington. 1996. Spiders of Pakitza (Madre de Dios, Peru): species richness and notes in community structure, 241-299. In E. Florez (ed.).1998. Estructura de comunidades de aranas (Araneae) en el departamento Del Valle, suroccidente de Colombia. Caldasia 20: 173-186. [ Links ]
Sosa, M.A., J. Farina Núñez & S. Mazza. 1998. Comparison of vertical cloth and whole plant bag sampling methods for estimating predator populations on cotton. World Cotton Research 2, Athens, Greece. [ Links ]
Uetz, G.W., J. Halaj & A.B. Cady. 1999. Guild structure of spiders in major crops. J. Arachnol. 27: 270-280. [ Links ]
Whitcomb, W. 1980. Sampling Spiders in Soybean Fields, p. 544-551. In M. Kogan & D.C. Herzog. 1980. Sampling Methods in Soybean Entomology. Springer, Nueva York, EEUU. [ Links ]
Whitehouse, M.E.A., L.J. Wilson & G.P. Fitt. 2005. A Comparison of Arthropod Communities in Transgenic Bt and Conventional Cotton in Australia. Environ. Entomol. 34: 1224-1241. [ Links ]
Whitmore, C., R. Slotow, T.E. Crouch & A.S. Dippenaar-Schoeman. 2002. Diversity of spiders (Araneae) in a savanna Reserve, Northern Province, South Africa. J. Arachnol. 30: 344-356. [ Links ]
Wise, D.H. 1993. Spiders in ecological webs. Cambridge University, Cambridge, EEUU. [ Links ]
Young, O.P. & G.B. Edwards. 1990. Spiders in United States field crops and their potential effect on crop pests. J. Arachnol. 18: 1-27. [ Links ]
Zvedeniuck, I. 2009. Diversidad de Araneidae (Arachnida:Araneae) en la Reserva Natural e Histórica Isla Martin García. Departamento de Ciencias Biológicas, Universidad CAECE, Buenos Aires, Argentina. [ Links ]
*Correspondencia a: Melina Soledad Almada & Alda Gonzalez: Centro de Estudios Parasitologicos y de Vectores (CEPAVE) (CONICET LA PLATA- UNLP), Calle 2 No 584 (1900) La Plata, Argentina; melinalmada02@yahoo.com.ar, asgonzalez@cepave.edu.ar
María Ana Sosa: Estación Experimental Agropecuaria (EEA) INTA Reconquista, Santa Fe, Ruta 11 Km 773 (3560) Reconquista, Argentina; msosa@correo.inta.gov.ar
1. Centro de Estudios Parasitologicos y de Vectores (CEPAVE) (CONICET LA PLATA- UNLP), Calle 2 No 584 (1900) La Plata, Argentina; melinalmada02@yahoo.com.ar, asgonzalez@cepave.edu.ar
2. Estación Experimental Agropecuaria (EEA) INTA Reconquista, Santa Fe, Ruta 11 Km 773 (3560) Reconquista, Argentina; msosa@correo.inta.gov.ar
Recibido 07-IV-2011. Corregido 26-VIII-2011. Aceptado 30-IX-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}