










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.2 San José Jun. 2012
Isolation and characterization of infectious Vibrio sinaloensis strainsfrom the Pacific shrimp Litopenaeus vannamei (Decapoda: Penaeidae)
Ma. del Carmen Flores-Miranda1*, Antonio Luna-Gonzalez1, Angel I. Campa Cordova2*, Jesus A. Fierro-Coronado1, Blanca O. Partida-Arangure1, Jose Pintado3* & Hector A. Gonzalez-Ocampo1
*Dirección para correspondencia:
Abstract
Infectious diseases especially those caused by bacterial and viral pathogens are serious loss factors in shrimp farming. In this study, bacteria were isolated from the gut and hepatopancreas of stressed shrimps obtained from a commercial farm. The isolates were screened on Thiosulfate citrate bile salt sucrose (TCBS) agar plates for the selection of Vibrio species. Presumptive vibrios were characterized through tests for hemolytic and enzymatic activity, hydrophobicity, growth and molecular identification. Three experimental infections were conducted in order to confirm the pathogenicity of selected bacterial strains VHPC18, VHPC23, VHPC24 and VIC30. In the third experimental challenge the LD50 was obtained, it lasted 10 days with 10 shrimp, weighing 6.9±1.1g, per tank. The treatments in triplicate were: (1) saline solution (control group); (2) 2×105CFU/shrimp; (3) 4×105CFU/shrimp; (4) 2×106CFU/shrimp; (5) 4×106CFU/shrimp, and (6) 8×106CFU/shrimp. In all challenges, water parameters measured during the experimental period remained within optimum ranges. Pathogenicity tests confirmed that the mixture of four vibrio isolates, identified as Vibrio sinaloensis, was virulent for L. vannamei. The LD50 value was 1.178×105CFU/g body weight. V. sinaloensis may act as opportunistic pathogens for cultured L. vannamei. Rev. Biol. Trop. 60 (2): 567-576. Epub 2012 June 01.
Key words: Vibrio sinaloensis, vibriosis, hemolytic activity, enzymatic activity, Litopenaeus vannamei.
Resumen
Las enfermedades de etiologia infecciosa, especialmente las causadas por patogenos bacterianos y virales ocasionan graves perdidas en el cultivo de camaron blanco Litopenaeus vannamei. En este estudio se caracterizo: la actividad enzimatica y hemolitica; hidrofobicidad; crecimiento e identificacion molecular de vibrios aislados del intestino y hepatopancreas de camarones estresados, obtenidos de una granja comercial, en medio Agar Tiosulfato Citrato Bilis Sacarosa. Ademas, se realizaron tres infecciones experimentales para confirmar la patogenicidad de las cepas bacterianas seleccionadas VHPC18, VHPC23, VHPC24 y VIC30. En la tercera infeccion experimental se obtuvo la LD50, el reto duro 10 dias, con 10 camarones por tanque con un peso de 6.9±1.1g. Los tratamientos se realizaron por triplicado: (1) solucion salina (grupo control); (2) 2×105UFC/camaron; (3) 4×105UFC/camaron; (4) 2×106UFC/camaron; (5) 4×106UFC/camaron y (6) 8×106UFC/camaron. En todos los retos, los parámetros del agua permanecieron dentro de los intervalos optimos. Las pruebas de patogenicidad confirmaron que la mezcla de cuatro aislados de Vibrio, identificados como Vibrio sinaloensis, fue virulenta para L. vannamei. El valor de la LD50 fue de 1.178×105UFC/g de peso corporal. Los resultados permiten establecer que las cepas de V. sinaloensis pueden actuar como patogenos oportunistas en L. vannamei cultivado.
Palabras clave: Vibrio sinaloensis, vibriosis, actividad enzimatica, actividad hemolitica, Litopenaeus vannamei.
In shrimp farming, the Pacific whiteleg shrimp (Litopenaeus vannamei) is the primary penaeid shrimp currently being cultured in Central and South America (Burge et al. 2007). However, with the rapid development of shrimp aquaculture, infectious diseases especially caused by bacterial and viral pathogens are serious loss factors in shrimp farming (Lightner 1996, Lavilla-Pitogo et al. 1998, Primavera 1998, de la Pena et al. 2003).
Bacteria are etiological agents that generate diseases, and Vibrionaceae represent the most important group of pathogens for both larvae and juvenile shrimp. Vibrio species are microorganisms that live in the shrimp’s environment, often as part of the normal microflora inhabiting the surface of their cuticle or colonizing areas of the gut or hepatopancreas. Vibrio are Gram-negative, facultative anaerobes, comma-shaped rods found both in freshwater and marine ecosystems (Brock & Lightner 1990).
Vibrio act as opportunistic or secondary pathogens that can cause mortality from a few to 100% in affected populations under stress (Lightner 1988). Horowitz & Horowitz (2001) postulated that if shrimp are not suffering from primary infections, physical damage or stress, their resistance against vibrios is adequate to prevent disease. This idea was further supported by Alday-Sanz et al. (2002), who showed that shrimp, when exposed to ammonia prior to an immersion challenge with Vibrio, suffered more frequent and earlier pathological changes than shrimp exposed to the bacterium alone.
Variations in virulence between strains of the same Vibrio species are a common phenomenon (Soto-Rodriguez et al. 2003) but, perhaps, pathogenic strains are the exception rather than the rule. Therefore, it is important to know the normal Vibrio microbiota of a cultured marine organism in order to understand better the rol of a certain bacterial strain or species in a pathogenic process (Gomez-Gil et al. 2008).
Vibriosis is a serious threat to the aquaculture industry, responsible for massive mortality of cultured penaeids worldwide (Baticados et al. 1990). This disease is mainly caused by specific strains of Vibrio anguillarum, V. alginolyticus, V. parahaemolyticus, V. harveyi, V. penaeicida, V. campbellii, both in hatcheries and in grow-out cultures (Lightner 1988, Lavilla-Pitogo et al. 1990, Rattanama et al. 2009).
Vibrio sinaloensis was isolated and described for the first time from cultured spotted rose snapper, Lutjanus guttatus in Mazatlan, state of Sinaloa, Mexico (Gomez-Gil et al. 2008); however, this is the first time that V. sinaloensis is isolated from L. vannamei shrimp.
This study was undertaken to isolate and characterize Vibrio isolates, with pathogenic potential, from the gut and hepatopancreas of whiteleg shrimp. Experimental infections were performed to assess the potential pathogenic capability of the isolates.
Materials and methods
Isolation of Vibrio from gut and hepatopancreas: Bacteria were isolated from the gut and hepatopancreas of six juvenile shrimp (7.7±0.7g). Animals were collected in August (2008) from a commercial farm Acuicola Cuate Machado (Guasave, Sinaloa, Mexico, 25°18’42.83’’ N - 108°29’22.41’’ O) reported to have mortalities due to White-spot syndrome virus. In the laboratory, guts and hepatopáncreas of shrimp were aseptically removed and placed into Eppendorf tubes with 200µL of sterile saline solution (2.5% NaCl). Shrimp tissues were homogenized with a pestle. The homogenate (100μL) was inoculated into Thiosulphate Citrate Bile Sucrose (TCBS, BD Bioxon, Cat. No.265020) agar supplemented with 2.5% NaCl. The plates were kept at 37°C for 24h. Colonies were selected and streaked onto TCBS plates and incubated as above. The isolates maintained in pure culture were stored at -85°C in Trypticase Soy (TS, BD Bioxon, Cat. No. 211670) broth with 2.5% NaCl and 15% (v/v) glycerol.
Characterization of the isolates: The isolates were characterized using Gram stain and based on cellular morphology. In addition, hemolytic activity (HA), hydrophobicity, extracellular enzymatic activity, and kinetics of bacterial growth were studied to be used as criteria to select potential pathogens (see below). Furthermore, molecular identification was done as part of the characterization process.
Hemolysis assay: Hemolysis was determined according to Apun-Molina et al. (2009). The supernatant fraction of overnight cultures of each isolate in TS broth was obtained by centrifugation at 10 000g for 10min and tested for its hemolytic activity on blood agar (BA, BD Bioxon, Cat. No. 273300) supplemented with 5% (v/v) heparinized human blood. Wells of 6mm diameter were made on the BA-supplemented Petri plates. The wells were filled each with 50μL supernatant or TS broth (negative control) and incubated for 24h at 37°C. A clear zone surrounding the well indicated hemolytic activity (a or β). The isolates with β-hemolysis activity were selected as potential pathogens and were used for further analysis.
Congo red binding assay: The Congo red (CR, Sigma, Cat. No. C6277) binding assay was performed by adding 0.03% (w/v) Congo ed to the TS agar medium supplemented with 1.0% NaCl. Each isolate was streaked and incubated at 37°C for 24h. The deep red coloration of colonies was considered positive, indicating a hydrophobic response of the isolate (Sharma et al. 2006). The β-hemolytic isolates with larger lysis diameter and Congo red-positive were selected as potential pathogens and were used for further analysis.
Extracellular enzymatic activity: Extracellular protease and lipase activities were determined according to Leon et al. (2000). Basal medium (1.5% agar and 0.5% yeast extract) supplemented with 2% skim milk or 1% gelatin was used to test the proteolytic activity (PA) of supernatants of 24h cultures. Lipolytic activity (LA) was tested in basal medium plates supplemented with 1% Tween 80 (Sigma, Cat. No. P1754). Wells of 6mm diameter were made on plates, then filled with 50µL supernatant and incubated at 37°C for 24h. Sterile TS broth supplemented with 2.5% NaCl was used as negative control. A clear zone around the well was considered positive for proteolytic activity and a cloudy zone around the well was considered positive for lipolytic activity.
Kinetics of bacterial growth: Bacterial growth kinetics was assessed to determine the log growth phase of each isolate. From the stock at -85°C, 20μl were inoculated in 50mL TS broth, supplemented with 2.5% NaCl, and incubated at 37°C for 24h. Bacterial growth was measured by reading absorbance in a Thermo Spectronic Genesys 2 Spectrophotometer (Thermo Scientific, Waltham, MA, USA) at 580nm. Measurements were made at 3, 6, 9, 12, 24, and 48h.
Bacterial count: Strains were grown for 24h in TS broth to count the colony forming units (CFU). Bacteria were centrifuged at 10 000g during 20min at room temperature and the cellular pellet was washed two times with sterile saline water (2.5% NaCl) and resuspended in 1mL of the same water. The bacterial suspension was then adjusted to an optical density of one in a Thermo Spectronic Genesys 2 Spectrophotometer at 580nm. To determine the CFU/mL of bacterial suspension, we used the serial dilution method.
Molecular identification: DNA extraction was performed with Bactozol kit (MRC, Cincinnati, OH, USA), and a 1 500-bp fragment of the 16S rRNA gene was amplified by using primers 27f and 1 492r (Jensen et al. 2002). PCR products were cleaned with spin columns and quantified with Quant-iT™ dsDNA HS kit (Invitrogen, Carlsbad, CA, USA). PCR products were tested for DNA sequencing. Bacterial sequences were subjected to BLAST searches (Zhang et al. 2000) by using the National Center for Biotechnology Information GenBank database.
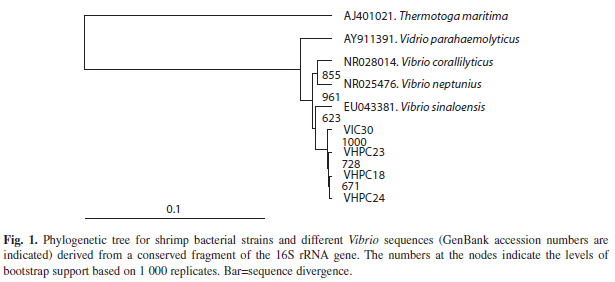
Phylogenetic analysis: Bacterial smallsubunit rRNA sequences were aligned with other Vibrio sequences by using the CLUSTAL X program (Larkin et al. 2007). Evolutionary relationships among the defined rRNA sequences were inferred by using the neighborjoining method (Saitou & Nei 1987) in the TreeView® 1.6 software program (Page 1996). The accuracy of the resulting tree was measured by bootstrap resampling of 1 000 replicates. The Thermotoga maritima sequence was used as out-group.
Experimental infections: In order to confirm the pathogenicity of the bacterial strains isolated from juvenile shrimp VHPC18, VHPC23, VHPC24, and VIC30, three experimental challenges were conducted. Healthy shrimp (L. vannamei) obtained from a local commercial farm (Acuicola Cuate Machado) in October (2008) and May (2009) were maintained in the laboratory in 1 000-L plastic tanks with 400L filtered (20μm) sea water with constant aeration. The healthy shrimp selection was done based on visible features.
In the experimental challenges, animals were maintained in an indoor culture system in 120-L plastic tanks with 80L filtered (20μm) sea water and constant aeration. Shrimp were acclimated to culture conditions for five days. Each treatment had two or three replicates, with 5-10shrimp/tank. Shrimp were fed with commercial feed (Purina, Ciudad Obregon, Sonora, Mexico, 40% protein) twice daily at 09:00 and 17:00 hours. Uneaten food and waste matter were removed daily, and every three days half of the water was changed. Values of pH (HI 98127 pHep, Hanna Instruments), salinity (Refractometer W/ATC 300011, Sper Scientific), and dissolved oxygen and temperature (YSI model 55 Oxygen meter, Yellow Spring Instruments) were determined every three days.Accumulated mortality was recorded daily over the culture period and the results were expressed in survival rate (%).
Overnight cultures (TS broth) of the bacterial strains to be tested were washed by centrifugation (10 000g for 10min) and suspended in sterile saline solution (2.5% NaCl). The bacterial suspensions were adjusted to an optical density of one. The experimental inoculation of bacteria was performed with a mixture containing isolates VHPC18, VHPC23, VHPC24, and VIC30 at the same proportion. Shrimp were injected into the first abdominal segment with 40μL of either this bacterial mixture or saline solution for the control group (2.5% NaCl) using a sterile 1mL syringe with a 25-gauge needle.
Experimental challenge I: This experiment was conducted for 10 days with five shrimp, weighing 9.8±2.5g, per tank (n=60). The experiment was conducted as a completely randomized design with six treatments in duplicate: (1) saline solution (control group); (2) 4×100CFU/shrimp; (3) 4×101CFU/shrimp; (4) 4×102CFU/shrimp; (5) 2×103CFU/shrimp, and (6) 4×103CFU/shrimp. Water temperature was maintained at 23.4±0.8oC, pH at 8.1±0.1, oxygen at 6.4±0.4mg/mL, and salinity at 35psu.
Experimental challenge II: This experiment was conducted for 10 days with five shrimps, weighing 10.5±2.6g, per tank (n=60) and completely randomized design with six treatments in duplicate: (1)saline solution (control group); (2) 2×105CFU/shrimp; (3)4×105CFU/shrimp; (4)2×106CFU/shrimp;(5) 4×106CFU/shrimp, and (6) 8×106CFU/shrimp. Water temperature was maintained at 23.38±0.2oC, pH at 8.2±0.1, oxygen at 6.2±0.2mg/mL, and salinity at 35psu.
Experimental challenge III: This experiment was conducted for 10 days with 10 shrimp, weighing 6.9±1.1g, per tank (n=180). Treatments were in triplicate. The rest of the experimental conditions were as in experimental challenge II. Water temperature was maintained at 25.8±0.4°C, pH at 8.1±0.1, oxygen at 6.1±0.4mg/mL, and salinity at 35psu.
In all challenges, water parameters measured during the experimental period remained within optimum ranges (Brock & Main 1994).
LD50: The results obtained in the third experiment were used to calculate the median lethal dose (LD50) by using Probit analysis (Finney 1952) with PASWR Statistics Ver. 18.
One-way analysis of variance (ANOVA) using the F test was applied to examine the differences in survival (%) among treatments. Survival data were arcsine transformed according to Daniel (1997). Where significant ANOVA differences were found, Tukey’s HSD test was used to identify the nature of differences at p<0.05.
Results
Isolation of Vibrio from gut and hepatopancreas: Thirty presumptive Vibrio strains were isolated from the hepatopancreas (25) and gut (five) of L. vannamei.
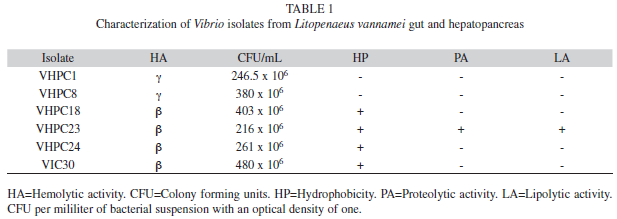
Characterization of the isolates: The selected isolates formed green colonies in TCBS medium, and were Gram-negative comma-shaped rods. Three isolates depicted γ-hemolysis, 11 α-hemolysis and 16 ß-hemolysis in BA. The ß-hemolytic isolates VHPC18, VHPC23, VHPC24, and VIC30 showed larger lysis diameter (8.0, 8.0, 13.0, and 13.5mm, respectively) and were selected for further characterization (Table 1). In addition, the γ-hemolytic isolates VHPC1 and VHPC8 were preserved to be used as comparison in further analyses. The β-hemolytic isolates were Congo red positive (hydrophobic) and showed pigmented colonies. The γ-hemolytic isolates showed non-pigmented colonies. The isolate VHPC23 showed proteolytic and lipolytic activities. The diameter of the halo in casein and gelatin hydrolysis tests was 16 and 22mm, respectively. In the hydrolysis with Tween 80, the diameter of the halo was 7mm (Table 1).
All isolates showed a log phase between three and nine hours. When bacterial suspensions with absorbance of one were counted, colony forming units of selected isolates were between 216×106 to 480×106CFU/mL of bacterial suspension (Table 1).
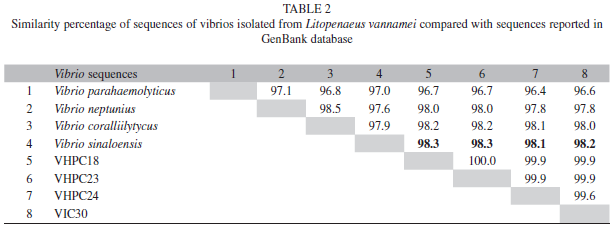
Molecular identification: The sequences of the conserved fragment of the 16S rRNA gene amplified by PCR were used for BLAST homology searches, and the results of bacterial strains identification revealed that isolates appeared to be related (98% homology) to Vibrio sinaloensis (accession number EU043381) (Gomez-Gil et al. 2008). Additionally, these sequences were also compared with the MegAlign program of DNASTAR® software (version 2.0 Madison, Wisconsin, USA). The observed homology between the sequences of isolates VHPC18 and VHPC23 was 100%, these isolates showed a 99.9% homology with the isolate VHPC24. The isolate VIC30 presented a homology of 99.9%, 99.9% and 99.6% with isolates VHPC18, VHPC23 and VHPC24, respectively (Table 2).
Phylogenetic analysis: The phylogenetic tree that was constructed mainly with sequences from four Vibrio strains clearly grouped the isolates with V. sinaloensis (Fig. 1).
Experimental challenge I: Challenge I revealed that a dose up to 103CFU/shrimp was insufficient to cause mortality, since survival was 100% in all treatments.
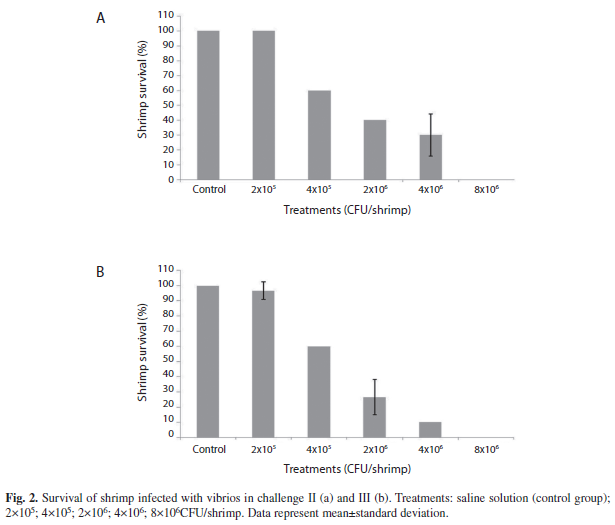
Experimental challenge II: The results obtained with challenge II showed that the mean survival of shrimp in the control treatment was 100±0.0%. In treatments with 2×105, 4×105, 2×106, 4×106 and 8×106CFU/shrimp, survival was 100±0.0, 60±0.0, 40±14.1, 35±0.0 and 0%, respectively (Fig. 2a). Signs of infection included brown or black spots on the shell, murky whitish muscle, and lack of food in the gut.
Experimental challenge III: The results obtained in experiment III showed that mean survival of shrimp in the control treatment was 100±0.0%. In treatments with 2×105, 4×105, 2×106, 4×106 and 8×106CFU/shrimp, survival was 97±5.7, 60±0.0, 27±11.5, 10±0.0 and 0%, respectively. Signs of infection were similar to those of experiment II (Fig. 2b).
LD50: Probit analysis yielded an LD50 value of 1.178×105 CFU/g body weight.
Discussion
Severe stress and injury to shrimp under poor environmental conditions lower their resistance, rendering them susceptible to viral as well as bacterial infection (Liu 1990). Vibriosis is known to affect a wide range of fish and shellfish organisms (Brock & LeaMaster 1992, Aguirre-Guzman 2004). In this study, 30 isolates of presumptive vibrios were obtained from the hepatopancreas and gut of L. vannamei. Isolates with pathogenic potential were selected according to their ability to lyse erythrocytes (Joseph et al. 1982, Zamora-Rodriguez 2003), positive hydrophobicity (Khuntia et al. 2008), and production of extracellular enzymes such as proteases and lipases (Farzanfar 2006, Balcazar et al. 2006).
Hemolytic bacteria are able to synthesize exotoxins that cause partial or total lysis of blood erythrocytes of different animals (Zamora-Rodriguez 2003), which was a desirable feature in our isolation. Joseph et al. (1982) mentioned that hemolytic strains are pathogenic in nature. However, it is important to note that although hemolytic activity is considered one of the pathogenic characteristics of bacteria, it is not always useful in determining pathogenicity. For example, both hemolytic and non-hemolytic strains of Streptococcus are important human pathogens (Michael et al. 1988). This suggests the need for further tests such as hydrophobicity.
The β-hemolytic isolates were Congo red positive (hydrophobic) showing pigmented colonies. A positive result indicates that the bacteria have the ability to bind nonspecifically to the epithelium of the shrimp intestine by hydrophobic interactions. In the absence of hydrophobic molecules on the surface of the bacteria and the epithelium, they would repel, as both have negative charge. Thus, hydrophobic interactions favor the adhesion and colonization ability of a pathogen (An & Friedman 2000, Rinkinen et al. 2003).
In addition to hemolysis, bacterial pathogenic factors such as enterotoxin, protease and hemaglutinin excretions have been reported for aquatic organisms (Inamura et al. 1984). In this study, only the selected isolate VHPC23 showed proteolytic and lipolytic activities. Some authors claim that the production of extracellular enzymes such as proteases and lipases help the nutrition of the host (Farzanfar 2006, Balcazar et al. 2006), whereas others believe that the overproduction of these enzymes is a virulence factor, since pathogenic strains have high proteolytic, extracellular lipolytic and hemolytic activity (Quesada-Herrera et al. 2004).
The four isolates with pathogenic potential were classified by using only molecular techniques. Despite the fact the amplified sequence of the 16S rRNA gene was quite conserved, the bacterial identity determined by homology searches and by using the generated phylogenetic tree corresponded with V. sinaloensis. Isolates VHPC18 and VHPC23 are genotypically identical for the sequenced 16S rRNA gene fragment, even though different phenotypical behavior, related with proteolytic and lipolytic activities, was observed. V. sinaloensis was firstly isolated from cultured spotted rose snapper (Lutjanus guttatus) in the same geographical area (state of Sinaloa, Mexico) (Gomez-Gil et al. 2008).
The results obtained in the experimental infection of shrimp with intramuscular injection of pooled strains induced high mortality at higher doses. Albeit the most works, colleagues use one strain for challenges, it is important to remark that interactions between microorganisms and with the host are very complex. Vibrio may act as primary and secondary/opportunistic pathogens of shrimp, and synergistic effects may occur among them (Austin & Austin 1993). Therefore, in this study, we decided to infect shrimp with a mixture of Vibrio isolates with pathogenic potential rather than with a single one. Moreover, a synergic effect could be expected between the strain with proteolytic and lipolytic activity (VHPC23) and the strains without them (VPC18, VHPC24, VIC30).
In shrimp, the different natural routes of infection by virulent bacterial isolates are, theoretically: oral, trans-cuticular, or caused by wounds, by an imbalance in the natural bacterial flora, or by vertical transmission of the pathogen (Saulnier et al. 2000). However, we used intramuscular infection to force the disease (vibriosis) and to obtain an intermédiate mortality (LD50), suitable to test the effect of feed additives (probiotics, prebiotics, immunostimulants) in future works with pathogens. The LD50 value (1.178×105CFU/g shrimp) in L. vannamei was higher than the LD50 (2.5×104CFU/g shrimp) obtained by Jayasree et al. (2006) for V. harveyi isolated and tested in Penaeus monodon, but similar to the LD50 (1.13×105CFU/g shrimp) obtained by Lee et al. (1996), who challenged P. monodon with V. alginolyticus. However, our LD50 was lower than the LD50 (2.5×105CFU/g shrimp) reported by Song et al. (1993), who challenged P. monodon with V. damsela or the LD50 (2.46×105CFU/g shrimp) found by Lee et al. (1996), who challenged P. japonicus with V. alginolyticus.
Based on the results obtained in this study, we established that strains of V. sinaloensis may act as opportunistic pathogens in cultured L. vannamei.
Acknowledgments
Authors are grateful to Consejo Estatal de Ciencia y Tecnologia del Estado de Sinaloa (CECyT-Sinaloa) and to the Secretaria de Investigacion y Posgrado del Instituto Politecnico Nacional (SIP-IPN) for financial support. Ma. del Carmen Flores Miranda acknowledges CONACYT-Mexico and SIP-IPN for the M.Sc. grants.
References
Aguirre-Guzman, G. 2004. ¿Los Vibrio sp. son agentes patogenos importantes para el cultivo de camaron? Programa Nacional de Sanidad Acuicola y la Red de Diagnostico 1: 1-9. [ Links ]
Alday-Sanz, V., A. Roque & J.F. Turnbull. 2002. Clearing mechanisms of Vibrio vulnificus biotype I in the black tiger shrimp Penaeus monodon. Dis. Aquat. Organ. 48: 91-99. [ Links ]
An, Y.H. & R.J. Friedman. 2000. Handbook of bacterial adhesion: principles, methods and applications. Humana, Inc. New Jersey, USA. [ Links ]
Apun-Molina, J.P., A. Santamaria-Miranda, A. Luna-Gonzalez, S.F. Martinez-Diaz & M. Rojas-Contreras. 2009. Effect of potential probiotic bacteria on growth and survival of tilapia Oreochromis niloticus L., cultured in the laboratory under high density and suboptimum temperature. Aquac. Res. 40: 887-894. [ Links ]
Austin, B. & D.A. Austin. 1993. Bacterial fish pathogens: disease in farmed and wild fish. Ellis Horwood, New York, USA. [ Links ]
Balcazar, J.L., I. De Blas, I. Ruiz-Zarzuela, D. Cunningham, D. Vendrell & J.L. Muzquiz. 2006. The role of probiotics in aquaculture. Vet. Microbiol. 114: 173-186. [ Links ]
Baticados, M.C.L., C.R. Lavilla-Pitogo, E.R. Cruz-Lacierda, L.D. de la Pena & N.A. Sunaz. 1990. Studies on the chemical control of luminous bacteria Vibrio harveyi and V. splendidus isolated from diseased Penaeus monodon larvae and rearing water. Dis. Aquat. Organ. 9: 133-139. [ Links ]
Brock, J.A. & D.V. Lightner. 1990. Diseases of Crustacea, p. 245-424. In O. Kinne (ed.). Diseases of marine animals, vol 3. Biologische Anstalt Helgoland, Hamburg, Germany. [ Links ]
Brock, J.A. & B. LeaMaster. 1992. A look at the principal bacterial, fungal and parasitic diseases of farmed shrimp, p. 212-226. In J. Wyban (ed.). Proceedings of the special session on shrimp farming. World Aquaculture Society, Baton Rouge, Louisiana, USA. [ Links ]
Brock, J. & K.L. Main. 1994. A guide to the common problems and diseases of cultured Penaeus vannamei. World Aquaculture Society, Baton Rouge, Louisiana, USA. [ Links ]
Burge, E.J., D.J. Madigan, L.E. Burnett & K.G. Burnett. 2007. Lysozyme gene expression by hemocytes of Pacific white shrimp, Litopenaeus vannamei, after injection with Vibrio. Fish Shellfish Immunol. 22: 327-339. [ Links ]
Daniel, W.W. 1997. Bioestadistica. Base para el analisis de las ciencias de la salud. Limusa, Mexico, D.F. [ Links ]
de la Pena, L.D., C.R. Lavilla-Pitogo, A. Namikoshi, T. Nishizawa, Y. Inui & K. Muroga. 2003. Mortality in pond-cultured shrimp Penaeus monodon in the Philippines associated with Vibrio harveyi and white spot syndrome virus. Fish Pathol. 38: 59-61. [ Links ]
Farzanfar, A. 2006. The use of probiotics in shrimp aquaculture. FEMS Immunol. Med. Microbiol. 48: 149-158. [ Links ]
Finney, D.J. 1952. Probit Analysis. Cambridge University, Cambridge, England. [ Links ]
Gomez-Gil, B., E. Fajer-Avila, J. Pascual, M.C. Macian, M.J. Pujalte, E. Garay & A. Roque. 2008. Vibrio sinaloensis sp. nov., isolated from the spotted rose snapper, Lutjanus guttatus Steindachner, 1869. Int. J. Syst. Evol. Micr. 58: 1621-1624. [ Links ]
Horowitz, A. & S. Horowitz. 2001. Disease control in shrimp aquaculture from a microbial ecology perspective, p. 199-218. In C.L. Browdy & D.E. Jory (eds.). The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture 2001. The World Aquaculture Society, Baton Rouge, Louisiana, USA. [ Links ]
Inamura, H., K. Muroga & T. Nakai. 1984. Toxicity of extracellular products of Vibrio anguillarum. Fish Pathol. 19: 89-96. [ Links ]
Jayasree, L., P. Janakiram & R. Madhavi. 2006. Characterization of Vibrio spp. Associated with Diseased Shrimp from Culture Ponds of Andhra Pradesh (India). J. World Aquac. Soc. 37: 523-532. [ Links ]
Jensen, S., O. Bergh, O. Enger & B. Hjeltnes. 2002. Use of PCR-RFLP for genotyping 16S rRNA and characterizing bacteria cultured from halibut fry. Can. J. Microbiol. 5: 379-386. [ Links ]
Joseph, S.W., R.R. Colwell & J.B. Kaper. 1982. Vibrio parahaemolyticus and related hallophilic Vibrios. Crit. Rev. Microbiol. 10: 73-124. [ Links ]
Khuntia, C.P., B.K. Das, B.R. Samantaray, S.K. Samal & B.K. Mishra. 2008. Characterization and pathogenicity studies of Vibrio parahaemolyticus isolated from diseased freshwater prawn, Macrobrachium rosenbergii (de Man). Aquac. Res. 39: 301-310. [ Links ]
Larkin, M.A., G. Blackshields, N.P. Brown, R. Chenna, P. McGettigan, H. McWilliam, F. Valentin, I.M. Wallace, A. Wilm, R. Lopez, J.D. Thompson, T.J. Gibson & D.G. Higgins. 2007. Clustal W and Clustal X version 2.0. Bioinformatics 23: 2947-2948. [ Links ]
Lavilla-Pitogo, C.R., E.M. Leano & M.G. Paner. 1998. Mortalities of pond-cultured juvenile shrimp, Penaeus monodon, associated with dominance of luminescent vibrios in the rearing environment. Aquaculture 164: 337-349. [ Links ]
Lavilla-Pitogo, C.R., C.L. Baticados, E.R. Cruz Lacierda & L.D. de La Pena. 1990. Occurrence of luminous bacterial disease of Penaeus monodon larvae in the Philippines. Aquaculture 91: 1-13. [ Links ]
Lee, K.K., S.R. Yu, T.I. Yand, P.C. Liu & F.R. Chen. 1996. Isolation and characterization of Vibrio alginolyticus from diseased kuruma prawn, Penaeus japonicus. Lett. Appl. Microbiol. 22: 111-114. [ Links ]
Leon, J., F. Pellon, V. Unda, J. David, C. Anaya & V. Mendoza. 2000. Produccion de enzimas extracelulares por bacterias aisladas de invertebrados marinos. Rev. Per. Biol. 7: 202-210. [ Links ]
Lightner, D.V. 1988. Vibrio disease in penaeid shrimp, p. 42-47. In C.J. Sinderman & D.V. Ligntner (eds.). Disease Diagnosis and Control in North American Marine Aquaculture. Elsevier, Amsterdam, Holland. [ Links ]
Lightner, D.V. 1996. A handbook of pathology and diagnostic procedures for diseases of cultured penaeid shrimp. World Aquaculture Society, Baton Rouge, Lousiana, USA. [ Links ]
Liu, C.I. 1990. The diseases of cultured Penaeus monodon with emphasis on recent discoveries in Taiwan, p. 180-201. In G.H. Kou, H. Wakabayashi, I.C. Liao, S.N. Chen & C.F. Lo (eds.). Fish diseases Proc. R. Jpn. Symp. Taipei, Taiwan. [ Links ]
Michael, J.P., E.C.S Chan & N.R. Krieg. 1988. Microbiology. McGraw-Hill, New York, USA. [ Links ]
Page, R.D.M. 1996. Treeview: an application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 12: 357-358. [ Links ]
Quesada-Herrera, A., M. Pozo & J. Rosa-Placencia. 2004. Seleccion de probioticos bacterianos para su uso en el cultivo de camaron. III Congreso Iberoamericano Virtual de Acuicultura. Comunicacion Tecnica CIVA 2004. 97-100 (tambien disponible en linea: http://www.civa2004.org). [ Links ]
Primavera, J.H. 1998. Tropical shrimp farming and its sustainability, p. 257-289. In S. de Silva (ed.). Tropical Mariculture. Academic, New York, USA. [ Links ]
Rattanama, P., K. Srinitiwarawong, J.R. Thompson, R. Pomwised, K. Supamattaya & V. Vuddhakul. 2009. Shrimp pathogenicity, hemolysis, and the presence of hemolysin and TTSS genes in Vibrio harveyi isolated from Thailand. Dis. Aquat. Organ. 86: 113-122. [ Links ]
Rinkinen, M., E. Westermarck, S. Salminen & A.C. Ouwehand. 2003. Absence of host specificity for in vitro adhesion of probiotic lactic acid bacteria to intestinal mucus. Vet. Microbiol. 97: 55-61. [ Links ]
Saitou, N. & M. Nei. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4: 406-425. [ Links ]
Saulnier, D., P. Haffner, C. Goarant, P. Levy & D. Ansquer. 2000. Experimental infection models for shrimp vibriosis studies: a review. Aquaculture 191: 133-144. [ Links ]
Sharma, K.K., S.S. Soni & S. Meharchandani. 2006. Congo red dye agar test as an indicator test for detection of invasive bovine Escherichia coli. Veterinarski Arhiv. 76: 363-366. [ Links ]
Sindermann, C.J. 1990. Bacteria, p. 41-74. Principal diseases of marine fish and shellfish, Vol 2. Academic, San Diego, California, USA. [ Links ]
Song, Y.L., W. Cheng & H.S. Wang. 1993. Isolation and characterization of Vibrio damsela infections for cultured shrimp in Taiwan. J. Invertebr. Pathol. 61: 24-31. [ Links ]
Soto-Rodriguez, S.A., A. Roque, M.L. Lizarraga-Partida, A.L.Guerra-Flores & B. Gomez-Gil. 2003. Virulence of luminous vibrios to Artemia franciscana nauplii. Dis. Aquat. Organ. 53: 231-240. [ Links ]
Zamora-Rodriguez, L.M. 2003. Aislamiento, identificación y conservación de cultivos de bacterias lácticas antagonistas de microbiota contaminante de sangre de matadero. Tesis de Doctorado, Universidad de Girona, Girona, España. [ Links ]
Zhang, Z., S. Schwartz, L. Wagner & W. Miller. 2000. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 7: 203-14. [ Links ]
*Correspondencia a: Ma. del Carmen Flores-Miranda, Antonio Luna-Gonzalez, Jesus A. Fierro-Coronado, Blanca O. Partida-Arangure & Hector A. Gonzalez-Ocampo: Centro Interdisciplinario de Investigacion para el Desarrollo Integral Regional (Unidad Sinaloa). Instituto Politecnico Nacional. Boulevard Juan de Dios Batiz Paredes 250, Guasave, Sinaloa 81101, Mexico; pamela_mya@hotmail.com, aluna@ipn.mx, arturofierrojr@hotmail.com, blanka_opa@hotmail.com, hgocampo@yahoo.com
Ángel I. Campa Córdova: Centro de Investigaciones Biologicas del Noroeste (CIBNOR), Mar Bermejo 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, Mexico; angcamp04@cibnor.mx
Jose Pintado: Instituto de Investigacions Marinas (CSIC), Eduardo Cabello No. 6, Vigo, Galicia, Espana; pintado@iim.csic.es
1. Centro Interdisciplinario de Investigacion para el Desarrollo Integral Regional (Unidad Sinaloa). Instituto Politecnico Nacional. Boulevard Juan de Dios Batiz Paredes 250, Guasave, Sinaloa 81101, Mexico; pamela_mya@hotmail.com, aluna@ipn.mx, arturofierrojr@hotmail.com, blanka_opa@hotmail.com, hgocampo@yahoo.com
2. Centro de Investigaciones Biologicas del Noroeste (CIBNOR), Mar Bermejo 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, Mexico; angcamp04@cibnor.mx
3. Instituto de Investigacions Marinas (CSIC), Eduardo Cabello No. 6, Vigo, Galicia, Espana; pintado@iim.csic.es
Received 29-IV-2011. Corrected 18-IX-2011. Accepted 19-X-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}