










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Glandular trichomes in Connarus suberosus (Connaraceae): distribution, structural organization and probable functions
*Dirección para correspondencia
Abstract
Connarus suberosus is a typical species of the Brazilian Cerrado biome, and its inflorescences and young vegetative branches are densely covered by dendritic trichomes. The objective of this study was to report the occurrence of a previously undescribed glandular trichome of this species. The localization, origin and structure of these trichomes were investigated under light, transmission and scanning electron microscopy. Collections were made throughout the year, from five adult specimens of Connarus suberosus near Botucatu, São Paulo, Brazil, including vegetative and reproductive apices, leaves and fruits in different developmental stages, as well as floral buds and flowers at anthesis. Glandular trichomes (GTs) occurred on vegetative and reproductive organs during their juvenile stages. The GTs consisted of a uniseriate, multicellular peduncle, whose cells contain phenolic compounds, as well as a multicellular glandular portion that accumulates lipids. The glandular cell has thin wall, dense cytoplasm (with many mitochondria, plastids and dictyosomes), and a large nucleus with a visible nucleolus. The starch present in the plastids was hydrolyzed during the synthesis phase, reducing the density of the plastid stroma. Some plastids were fused to vacuoles, and some evidence suggested the conversion of plastids into vacuoles. During the final activity stages of the GTs, a darkening of the protoplasm was observed in some of the glandular cells, as a programmed cell death; afterwards, became caducous. The GTs in C. suberosus had a temporal restriction, being limited to the juvenile phase of the organs. Their presence on the exposed surfaces of developing organs and the chemical nature of the reserve products, suggest that these structures are food bodies. Field observations and detailed studies of plant-environment interactions, as well as chemical analysis of the reserve compounds, are still necessary to confirm the role of these GTs as feeding rewards.
Key words: ant-plant interactions, cell ultrastructure, savanna, food body, trichome.
Resumen
Connarus suberosus es una especie típica del bioma Cerrado brasileño, y sus inflorescencias junto con los ramos vegetativos jóvenes están densamente cubiertos por tricomas dendríticas. El objetivo de este estudio es presentar la ocurrencia de un tricoma glandular sin descripción previa para esta especie. La localización, origen y estructura de estos tricomas se observó mediante microscopia óptica, así como electrónica de transmisión y de barrido. Los tricomas glandulares (TGs) se producen en los órganos vegetativos y reproductivos, en sus fases jóvenes. Los TGs consisten en un pedúnculo uniseriado y multicelular, cuyas células contienen compuestos fenólicos, así como un sitio en el cual las células glandulares acumulan compuestos pécticos, almidón y lípidos. La acumulación de aceite en el citosol es impresionante, pues ocupa una gran parte del volumen celular. La porción glandular presenta células con paredes delgadas, un denso citoplasma y un núcleo grande con nucleolo visible. El citoplasma contiene gran cantidad de mitocondrias, plástidos y dictiosomas. El almidón presente en los plástidos se hidroliza durante la fase de síntesis, de tal modo que reduce la densidad del estroma plastidial. Algunos plástidos se fusionan con las vacuolas, además hay pruebas de la conversión de los plástidos en vacuolas. Durante las etapas finales de la actividad de los TGs, se observó un oscurecimiento del protoplasma en algunas de las células glandulares lo que sugiere la existencia de la muerte celular programada; después de lo anterior, los TGs se convierten en caducas. Por lo tanto la aparición de TGs en las superficies expuestas de los órganos en desarrollo en C. suberosus y la naturaleza química de los productos de reserva sugieren que estas estructuras son cuerpos alimenticios. Hay evidencias sobre el papel de estos tricomas en las interacciones planta-hormiga, especialmente en el bioma Cerrado.
Palabras clave: Sabana, cuerpos alimenticios, interacciones planta-hormiga, tricoma, ultraestructura celular.
The family Connaraceae comprises 12 genera and from 110 to 200 arboreal, shrub, or vine species that are widely distributed in tropical regions. The majority of the genera occur in Africa, although the greatest species diversity is found in the Americas (Breteler 1989). Brazil is considered the center of the diversity of Connaraceae in the Neotropics with approximately 64 species, 30 of which belong to Connarus (Forero 1983, Forero & Costa 2002). According to Ratter et al. (2001), Connarus suberosus Planch. is one of the most frequent woody plant species encountered in the Cerrado (savannas) of Brazil.
Forero (1983) reported that C. suberosus demonstrates characteristics commonly associated with Cerrado plants, including a thick and corky bark, tortuous branches and dense pilosity. According to this author, young vegetative branches, inflorescences and fruits in their early stages of development show dense rustcolored tomentose indumentum, and glandular trichomes (GTs) may be present on their petals, stamens and styles.
Jongkind (1989) identified six types of trichomes in paleotropical members of the Connaraceae, of which two were glandular. According to this author, glandular trichomes, consisting of an uniseriate multicellular peduncle and a multicellular secretory portion that is generally very colorful, are found in Cnestis, Connarus, Jollydora, Manotes, and Rourea; a second type of unicellular glandular trichome is characteristic of Manotes. In these genera, the distribution of GTs is essentially limited to the stem axes and floral organs.
Studies involving neotropical taxon of Connaraceae have demonstrated the presence of GTs on the abaxial surfaces of leaflets of Pseudoconnarus subtriplinervis (Schellenberg 1938), on the leaves of some species of Rourea, and on the axes and floral organs of Connarus and Rourea (Forero 1976, 1983). Glandular trichomes have also been observed on the endocarp of paleo and neotropical species of Connarus (Dickison 1971, Corner 1976, Forero 1983, Jongkind 1989).
In spite of the reports mentioned above, little is known about the structural organization of GTs in Connaraceae, and their ultrastructural aspects, biological functions and the secretions chemical composition have not yet been well examined; furthermore, their possible roles in plant interactions are completely unknown.
Glandular trichomes are involved in the synthesis of diverse chemical compounds such as oils, sugars, salts, digestive enzymes and other similar compounds, and have important roles in plant interactions (Fahn 1979, Werker 2000, Wagner et al. 2004). These trichomes, whether isolated or in groups, are often secretory structures such as nectaries, oil producing or salt excreting glands (Fahn 1979). Other structures involved in plant interactions, such as colleters (Renobales et al. 2001, Paiva & Machado 2006, Paiva 2009), food bodies, and similar structures (Rickson & Risch 1984) are also considered trichomes in some species.
In the present study, a new type of glandular trichome is reported for Connarus suberosus, and provide information about their localization, development, structure, and ultrastructural and histochemical aspects, to support discussions concerning their functions.
Materials and methods
Material collection for morphological studies: Collections were made from adult specimens of Connarus suberosus Planch. Near Botucatu, São Paulo, Brazil (22º53’11.4” S - 48º26’07.8” W), including vegetative and reproductive apices, leaves and fruits, in different developmental stages, as well as floral buds and flowers at anthesis. The samples were examined using a stereoscopic microscope to determine the patterns of distribution and to verify the occurrence of trichomes. A voucher specimen was deposited in the “BOTU” Herbarium, Department of Botany, UNESP, Botucatu, SP, number 24415.
Light microscopy: In order to examine the ontogeny and anatomy of trichomes, samples of stem apices and leaves, in different developmental stages, were fixed in Karnovsky solution (Karnovsky 1965) for 24 hours, dehydrated in an ethanol series (Johansen 1940), and submitted to pre-infiltration and infiltration in (2-hydroxyethyl)-methacrylate (HEMA, Leica™), following standard methods. Transversal and longitudinal sections (5μm thickness) were made using a rotary microtome, and the sections were stained with 0.05% toluidine blue at pH 4.7 (O’Brien et al. 1964).
Histochemical tests: Histochemical characterizations were performed using the following tests, with their respective controls: Sudan black B for lipids in general (Pearse 1980); Lugol to identify starch (Johansen 1940); 10% ferrous chloride aqueous solution with the addition of a small portion of calcium carbonate to detect phenolic substances (Johansen 1940), and 0.02% aqueous solution of ruthenium red to detect pectic substances (Jensen 1962). These tests were performed on both free-hand sections of fresh material and microtome sections of samples embedded in HEMA.
Transmission electron microscopy: The samples were fixed in Karnovsky solution (Karnovsky 1965) for 24 hours, post-fixed in osmium tetroxide (1%, 0.1M phosphate buffer, pH 7.2) and processed according to standard methodologies (Roland 1978). The ultrathin sections were contrasted with uranyl acetate and lead citrate and examined using a Philips CM 100 transmission electron microscope at 60kV.
Scanning electron microscopy: Samples were fixed in 2.5% glutaraldehyde in 0.1M phosphate buffer, pH 7.2, dehydrated in an ethanol series, and dried to their critical point using CO2 (Robards 1978). These dried samples were glued on aluminum stubs, coated with a 10nm layer of gold, and examined using a Quanta 200 (FEI Company) scanning electron microscope at 15 to 20kV; all images were processed digitally.
Results
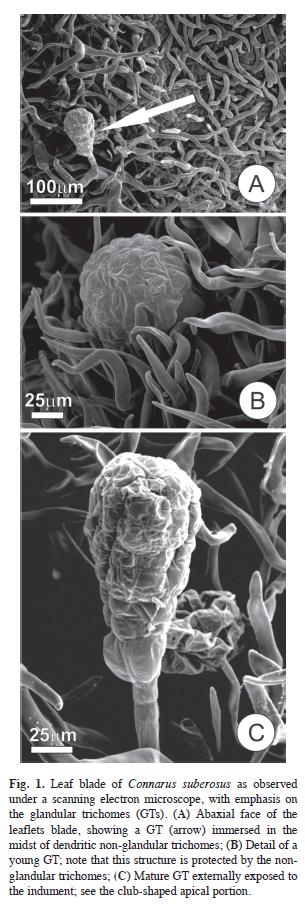
Distribution and ontogeny: Connarus suberosus has compound leaves, and has conduplicate leaflets when young. Glandular trichomes (GTs) occur on leaflets, bracteols and sepals, as well as on the surface of the stem and the axes of the inflorescences, and fruits during their juvenile stages. Young leaves show numerous GTs on the abaxial faces of the leaflets, especially along the midrib; although in lesser number, these structures also occur on the surfaces of the petioles and the rachis, mainly on their abaxial faces. GTs appear only occasionally on young stems and along the axes of the inflorescences. GTs are encountered in the midst of dendritic trichomes (Fig. 1A-C) that lend a rusty-tomentose aspect to the indument.
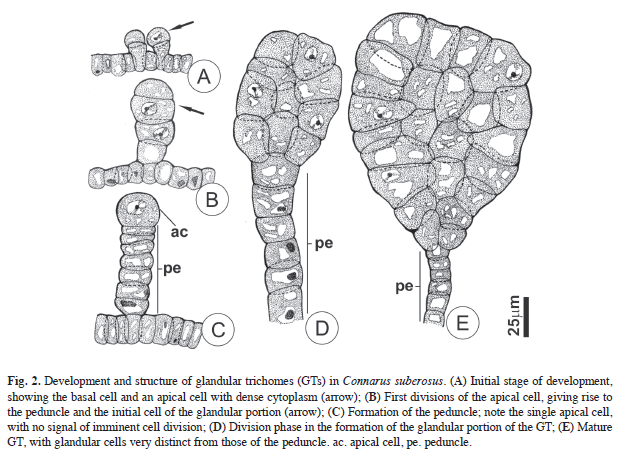
The development of glandular trichomes initiates with the distal expansion of a protodermal cell, producing a globe-shaped salience on the organ surface. Both a basal cell and an apical cell become defined (Fig. 2A) as a result of a periclinal division of this GT cell initial. The basal cell is funnel-shaped, and completes its differentiation in this stage. A periclinal division of the apical cell forms the peduncle cell initial, and an apical glandular initial (Fig. 2B). Successive periclinal divisions of the peduncle cell initial forms a multicellular and uniseriate peduncle (Fig. 2C). At this stage, the initial cell of the glandular portion remains in interphase, and only reinitiates division after the formation of the peduncle. This glandular initial cell then undergoes divisions in many planes, resulting in a glandular, multicellular club-shaped apical portion (Fig. 2D-E).
Structural and histochemical aspects: The GTs peduncle cells are short and cylindrical (Fig. 2D-E); the anticlinal faces of their cell walls are impregnated with lipidic substances. The peduncle cells of mature GTs are highly vacuolated, with the remaining cytoplasm being restricted to a thin layer compressed against the cell wall. No reserve compounds are accumulated in these cells, and the vacuoles contain large quantities of phenolic substances.
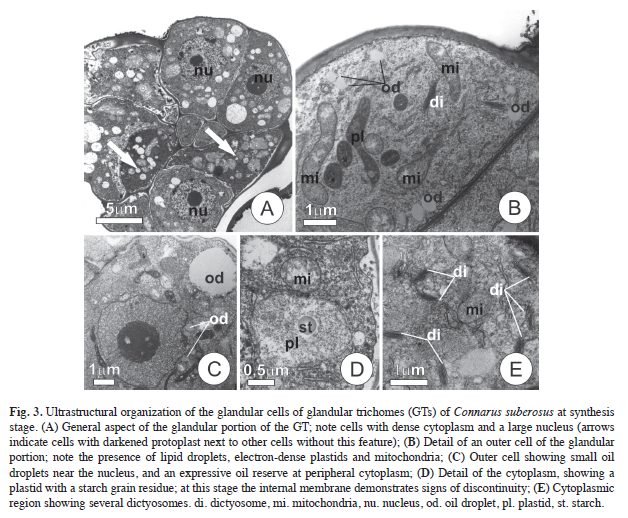
The cells of the glandular apical portion are compact and have thin cell walls, dense cytoplasm, a large nucleus, and a visible nucleolus (Fig. 3A, Fig. 3C). Pectic compounds, starch, and several lipid droplets can be seen in the cytoplasm of mature GTs. During the GTs final developmental stages, the vacuoles in some of the glandular cells increase in size and their protoplasts darkens; these changes are soon seen in neighboring cells, culminating in the cessation of synthesis activities and in GTs death that are caducous.
There is an expressive accumulation of lipids in the cells composing the apical portions of mature GTs in the form of droplets dispersed in the cytosol (Fig. 3B-C). Histochemical tests also indicated the presence of pectic secretions in the cytosol and subcuticular spaces, although in relatively small quantities. No evidence of any leakage of these secretions was seen under field or laboratory conditions.
Ultrastructural aspects and reserve accumulation: The cytoplasm of the apical cells of the GTs is rich in organelles; mitochondria, dictyosomes, plastids, and ribosomes are prevalent; the ribosomes are either free or associated with segments of the endoplasmic reticulum (Fig. 3A-D). The mitochondria have well-developed cristae and sometimes are juxtaposed with plastids (Fig. 3B-D). The endoplasmic reticulum appears to be closely associated with the plastids (Fig. 3D).
The plastids take on several shapes, but have only inconspicuous internal membrane systems, electron-lucent stroma, and starch grains. There is evidence for starch hydrolysis in mature GTs as the stroma becomes more electron-lucent and flocculated (Fig. 3D). The inner membranes of the plastids seem to rupture and this process is associated with the reduction of stroma density and the hydrolysis of the starch grains.
The dictyosomes have from four-seven cisternae, and their peripheral regions liberate small and not very dense vesicles, that become dispersed in the cytoplasmic matrix (Fig. 3E). Other interesting ultrastructural aspects include the presence of lipidic bodies in the cytoplasm, a reduced vacuole component and well-developed periplasmatic spaces (Fig. 3C-D).
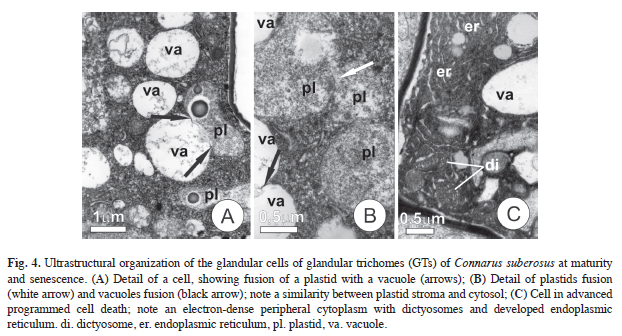
At the end of the synthesis stage, vacuoles had membranous inclusions and some fusions of plastids with vacuoles were seen (Fig. 4A-B). The vacuoles, which are about the same size as the plastids, have an amorphous content that is similar to the plastid stroma (Fig. 4A). Plastids fusion seems to occur only at this stage and appears to be similar to the fusion of these organelles with vacuoles. During this stage many GTs have cells with dense and dark cytoplasm, showing increased amounts of rough endoplasmic reticulum and active dictyosomes (Fig. 4C).
GTs remain immersed and protected in a dense mat of indument composed of nonglandular trichomes during their initial stages of development, but gradually emerge due to the lengthening of the peduncle cells. In this moment, lipidic reserves can be recognized.
Discussion
The dense indument of Connarus suberosus is composed by dendritic trichomes and it seems to confer an adaptative advantage to these plants by protecting them from desiccation (Fahn & Cutler 1992). Xylem transportation of water to developing and not yet fully photosynthetic organs is usually inefficient (Nobel et al. 1994) and makes them especially vulnerable to desiccation. In C. suberosus, a typical species of Cerrado vegetation, this vulnerability is particularly relevant, because Cerrado’s climate is markedly seasonal, with periods of very low relative humidity, high temperatures, and high solar irradiation (Franco 2002). Such conditions tend to favor the development of defensive structures against desiccation.
Previous studies with Connaraceae had demonstrated the presence of GTs principally on the floral axes and reproductive organs; only a few species develop these structures on vegetative organs. Forero (1983) and Forero & Costa (2002) reported the presence of GTs on the petal margins, on the surfaces of stamens and styles, and on the endocarp of C. suberosus; however, the characteristics referred by these authors differed markedly from the GTs described on vegetative organs, since those GTs showed short peduncle and smaller glandular head.
The presence of lipidic compounds on the anticlinal face of peduncle cells of the GTs of C. suberosus is typical of trichomes involved in secretory processes (see Fahn 1979), and apparently help to avoid the reflux of secretions. Considering the structural and histochemical aspects of the peduncle cells and the hydrolysis of starch registered in the glandular cells, it is possible to infer that starch constitutes an important energy source for both synthetic and secretory processes. Starch has been found to serve as the predominant energy reserve in other well-studied secretory structures (Fahn 1979, Paiva & Machado 2008), and the juxtaposition of mitochondria and amyloplasts indicate the energy need for starch hydrolysis, as discussed by Paiva & Machado (2008). For the species studied here, this process seems to furnish precursors for the synthesis of polysaccharides in the dictyosomes, as was observed by Andreeva et al. (1998). The vesicles that bud off from these organelles apparently transport secretion products to the plasmatic membrane, even though these compounds apparently do not accumulate in the periplasmatic and subcuticular spaces.
The observations of plastids fusing with vacuoles in the apical cells of the GTs, and the similarity between the contents of the vacuoles and the plastid stroma, strongly suggest a conversion of plastids into vacuoles, as has been reported in other types of secretory cells (Paiva & Machado 2008). It can also be inferred that this process culminates with the amplification of the vacuolar space in the final stages of the synthesis (Fahn 1979).
The modifications that were observed in the plastids occur during the final stages of synthesis activity and appear to mark the start of programmed cell death (PCD). Likewise, the darkening of the protoplast observed in some of the glandular cells of the GTs constitutes additional evidence of PCD, as described by Mohan & Inamdar (1986), Zer & Fahn (1992), and Paiva & Machado (2006). The co-existence of cells in different stages of cell death with fully viable and apparently healthy cells, reduces the possibility that these observations are due to technical artifacts or problems related to the sample fixing, reinforcing the idea that PCD is part of the normal developmental process of GTs.
The young, unexpanded leaflets of C. suberosus have a conduplicate blade that leaves only the abaxial face exposed, which seems to explain the distribution pattern of the glandular trichomes in those organs. Likewise, the bracteols and sepals that constitute the protective involucre of the floral buds during their different ontogenic stages also have their abaxial faces exposed. As such, the localization of the GTs and dendritic non-glandular trichomes in this species, seems to corroborate the interpretation that the GTs aid in the protection of developing organs (Mueller 1985). Some functions attributed to non-glandular and glandular trichomes include temperature regulation, reflectivity, and the reduction of transpiration (Wagner et al. 2004).
The occurrence of GTs in C. suberosus is restricted to the juvenile phase of the organs on which they appear and this, in association with the chemical nature of the reserve compounds that they contain, constitute evidence that these structures have a role in mediating symbiotic relationships that protect those developing organs; this function is attributed to food bodies in diverse plant species (O’Dowd 1980). Lipidic reserves are predominantly found in food bodies, although other compounds rich in energy or proteins can also be offered (O’Dowd 1982). It is worth pointing out that these GT structures are themselves protected in the midst of a dense indument of non-glandular trichomes, and that they emerge from this mat only when they have completed their development and lipidic reserves are present.
The lipids accumulated in the GTs of C. suberosus make these structures appealing food sources, and it is quite probable that they act as mediators of interactions between plants and ants, as observed in other species producing food bodies (Rickson 1980, Heil et al. 1998, Buono et al. 2008, Paiva et al. 2009). It is well established that food bodies help maintain mutualistic relationships between plants and ants, with the plants being benefited by protection against herbivores (O’Dowd 1980, 1982).
The GTs in C. suberosus have a temporal restriction, being limited to the juvenile phase of the organs. This is a common characteristic of situations in which young organs are protected by ants when their chemical and structural defenses have not yet been fully developed and they are most vulnerable to herbivory (Paiva et al. 2009, Possobom et al. 2010). As the GTs of C. suberosus are restricted to young organs and abscise before these organs mature, their role is most likely very similar to other food bodies described by the authors mentioned above.
Field observations and systematic studies of plant-environment interactions as well as the chemical analysis of the reserve compounds will still be necessary to confirm the role of the GTs of C. suberosus as feeding rewards. The ample occurrence of this species in the Cerrado biome confers a special relevance to a better understanding of the interactions mediated by these structures. According to Machado et al. (2008) ant-plant interactions in Brazilian Cerrado can reduce herbivore damage to leaves, buds or flowers in several plant species.
Acknowledgments
The authors would like to thank Luciano Milhomens for identifying the species, and FAPESP (Programa BIOTA, proc. no 00/12469- 3) for the financial support. João Donizete Denardi thanks CAPES for the PICDT grant, and Denise Maria Trombert Oliveira and Elder Antônio Sousa Paiva thank CNPq for the research productivity grant.
References
Andreeva, A.V., M.A. Kutuzov, D.E. Evans & C.R. Hawes. 1998. The structure and function of Golgi apparatus: a hundred years of questions. J. Exp. Bot. 49: 1281-1291. [ Links ]
Breteler, F.J. 1989. Connaraceae: a taxonomic study with emphasis on Africa. Agric. Univ. Wageningen Papers, Wageningen, Netherlands. [ Links ]
Buono, R.A., A.B. Oliveira & E.A.S. Paiva. 2008. Anatomy, ultrastructure, and chemical composition of food bodies of Hovenia dulcis (Rhamnaceae). Ann. Bot. 101: 1341-1348. [ Links ]
Corner, E.J.H. 1976. The seeds of dicotyledons. Cambridge University, England. [ Links ]
Dickison, W.C. 1971. Anatomical studies in the Connaraceae. I. Carpels. J. Elisha Mitchell Sci. Soc. 87: 77-86. [ Links ]
Fahn, A. 1979. Secretory tissues in plants. Academic, New York, USA. [ Links ]
Fahn, A. & D.F. Cutler. 1992. Xerophytes. Gebrüder Borntraeger, Berlin, Germany. [ Links ]
Forero, E. 1976. A revision of the american species of Rourea subgenus Rourea (Connaraceae). Mem. New York Bot. Garden 26: 1-119. [ Links ]
Forero, E. 1983. Connaraceae. Flora Neotropica 36: 1-208. [ Links ]
Forero, E. & C.B. Costa. 2002. Connaraceae, p. 85-92. In M.G.L. Wanderley, G.J. Shepherd & A.M. Giulietti (eds.). Flora fanerogâmica do Estado de São Paulo. v.2. FAPESP/ HUCITEC, São Paulo, Brazil. [ Links ]
Franco, A.C. 2002. Ecophysiology of woody plants, p. 178-197. In P.S. Oliveira & R.J. Marquis (eds.). The cerrados of Brazil: ecology and nature history of a neotropical savanna. Columbia University, New York, USA. [ Links ]
Heil, M., B. Fiala, W. Kaiser & K.E. Linsenmair. 1998. Chemical contents of Macaranga food bodies: adaptations to their role in ant attraction and nutrition. Funct. Ecol. 12: 117-122. [ Links ]
Jensen, W.A. 1962. Botanical histochemistry: principles and practice. W.H. Freeman, San Francisco, USA. [ Links ]
Johansen, D.A. 1940. Plant microtechnique. McGraw-Hill, New York, USA. [ Links ]
Jongkind, C.C.H. 1989. Indumentum, p. 19-21. In F.J. Breteler (ed.). The Connaraceae: a taxonomic study with emphasis on Africa. Agric. Univ. Wageningen Papers, Wageningen, Netherlands. [ Links ]
Karnovsky, M.J. 1965. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 27: 137A-138A. [ Links ]
Machado, S.R., L.P.C. Morellato, M.G. Sajo & P.S. Oliveira. 2008. Morphological patterns of extrafloral nectarines in woody plant species of the Brazilian cerrado. Plant Biology 10: 660-673. [ Links ]
Mohan, J.S.S. & J.A. Inamdar. 1986. Ultrastructure and secretion of extrafloral nectaries of Plumeria rubra L. Ann. Bot. 57: 389-401. [ Links ]
Mueller, R.J. 1985. Determinate branch development in Alstonia scholaris (Apocynaceae) the plagiotropic module. Am. J. Bot. 72: 1435-1444. [ Links ]
Nobel, P.S., J.L. Andrade, N. Wang & G.B. North. 1994. Water potentials for developing cladodes and fruits of a succulent plant including xylem-versus-phloem implications for water movements. J. Exp. Bot. 45: 1801-1807. [ Links ]
O’Brien, T.P., N. Feder & M.E. McCully. 1964. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 59: 368-373. [ Links ]
O’Dowd, D.J. 1980. Pearl bodies of a neotropical tree, Ochroma pyramidale: ecological implications. Am. J. Bot. 67: 543-549. [ Links ]
O’Dowd, D.J. 1982. Pearl bodies as ant food: an ecological role of some leaf emergences of tropical plants. Biotropica 14: 40-49. [ Links ]
Paiva, E.A.S., R.A. Buono & J.A. Lombardi. 2009. Food Bodies in Cissus verticillata (Vitaceae): ontogenesis, structure and functional aspects. Ann. Bot. 103: 517-524. [ Links ]
Paiva, E.A.S. 2009. Occurrence, structure and functional aspects of the colleters of Copaifera langsdorffii Desf. (Fabaceae, Caesalpinioideae). C. R. Biol. 332: 1078-1084. [ Links ]
Paiva, E.A.S. & S.R. Machado. 2006. Ontogenesis, structure and ultrastructure of Hymenaea stigonocarpa (Fabaceae–Caesalpinioideae) colleters. Rev. Biol. Trop. 54: 943-950. [ Links ]
Paiva, E.A.S. & S.R. Machado. 2008. The floral nectar of Hymenaea stigonocarpa (Fabaceae: Caesalpinioideae): structural aspects during floral development. Ann. Bot. 101: 125-133. [ Links ]
Pearse, A.G.E. 1980. Histochemistry theoretical and applied. v.2. Longman, London, England. [ Links ]
Possobom, C.C.F., E. Guimarães & S.R. Machado. 2010. Leaf glands act as nectaries in Diplopterys pubipetala (Malpighiaceae). Plant Biol. 12: 863-870. [ Links ]
Ratter, J.A., S. Bridgewater & J.F. Ribeiro. 2001. Espécies lenhosas da fitofisionomia cerrado sentido amplo em 170 localidades do bioma cerrado. Bol. Herb. Ezechias Paulo Heringer 7: 7-112. [ Links ]
Renobales, G., E. De Diego, B. Urcelay & A. López-Quintana. 2001. Secretory hairs in Gentiana and allied genera (Gentianaceae, subtribe Gentianinae) from the Iberian Peninsula. Bot. J. Linn. Soc. 136: 119-129. [ Links ]
Rickson, F.R. 1980. Developmental anatomy and ultrastructure of the ant–food bodies (Beccariian bodies) of Macaranga triloba and M. hypoleuca (Euphorbiaceae). Am. J. Bot. 67: 285-292. [ Links ]
Rickson, F.R. & S.J. Risch. 1984. Anatomical and ultrastructural aspects of the ant-food cell of Piper cenocladum C.DC. (Piperaceae). Am. J. Bot. 67: 543-549. [ Links ]
Robards, A.W. 1978. An introduction to techniques for scanning electron microscopy of plant cells, p. 343-403. In J.L. Hall (ed.). Electron Microscopy and Cytochemistry of Plant Cells. Elsevier, New York, USA. [ Links ]
Roland, A.M. 1978. General preparations and staining of thin sections, p. 1-62. In J.L. Hall (ed.). Electron Microscopy and Cytochemistry of Plant Cells. Elsevier, New York, USA. [ Links ]
Schellenberg, G. 1938. Connaraceae, p. 1-326. In A. Engler (ed.). Das Pflanzenreich, IV, 127. Engelmann, Leipzig, Germany. [ Links ]
Wagner, G.J., E. Wang & R.W. Shepherd. 2004. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 93: 3-11. [ Links ]
Werker, E. 2000. Trichome diversity and development. Adv. Bot. Res. 31: 1-35. [ Links ]
Zer, H. & A. Fahn. 1992. Floral nectaries of Rosmarinus officinalis L. structure, ultrastructure and nectar secretion. Ann. Bot. 70: 391-397. [ Links ]
*Correspondencia a: João Donizete Denardi. Universidade Estadual de Mato Grosso do Sul, Unidade Universitária de Coxim, Rua Pereira Gomes, 355, Vila Santa
Maria, 79.400-000, Coxim, MS, Brazil; joaodenardi@terra.com.br
Denise Maria Trombert Oliveira. Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Botânica, Avenida Antonio Carlos, 6627, Pampulha, Caixa Postal 486, 31.270-901, Belo Horizonte, MG, Brazil; dmtoliveira@icb.ufmg.br, epaiva@icb.ufmg.br
Elder Antônio Sousa Paiva. Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Botânica, Avenida Antonio Carlos, 6627, Pampulha, Caixa Postal 486, 31.270-901, Belo Horizonte, MG, Brazil; dmtoliveira@icb.ufmg.br, epaiva@icb.ufmg.br
1. Universidade Estadual de Mato Grosso do Sul, Unidade Universitária de Coxim, Rua Pereira Gomes, 355, Vila Santa
Maria, 79.400-000, Coxim, MS, Brazil; joaodenardi@terra.com.br
2. Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Botânica, Avenida Antonio Carlos, 6627, Pampulha, Caixa Postal 486, 31.270-901, Belo Horizonte, MG, Brazil; dmtoliveira@icb.ufmg.br, epaiva@icb.ufmg.br
Received 03-III-2011. Corrected 03-VI-2011. Accepted 05-VII-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}