










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Germination and seedling morphology of four South American Smilax (Smilacaceae)
Aline Redondo Martins1*, Anielca Nascimento Soares2*, Aline Bertolosi Bombo2, Alessandra Fidelis3*, Ana Dionisia da Luz Coelho Novembre4* & Beatriz Appezzato da Glória2
*Dirección para correspondencia
Abstract
Species of Smilax, also known as greenbrier, are widely distributed in Brazil and their commercial trades are carried out by the extractivism of native species. We the aim to provide information about the germination and development of seedlings in four Smilax species, different experiments were developed under controlled conditions. We evaluated two germination treatments: temperature (30ºC and 20-30ºC) and light (presence/ absence), and for few cases the tetrazolium treatment was applied. A different treatment response was observed among the studied species. Light had a significant influence in S. brasiliensis, with the highest germination rates at 20-30ºC in dark conditions. S. campestris showed significant differences among temperature treatments, but not to light; while S. cissoides showed high germination rates (66-78%), independently of treatment. However, S. polyantha had low germination rates (19-24%). After one year, the expanded leaves showed different characteristics among the studied species. Leaves of S. brasiliensis were ovate, coriaceous, three main veins and prickle-like structures only on the midrib on abaxial face. S. campestris leaves were oblong, coriaceous and prickle-like structures were located at the leaf midrib and margin. S. cissoides had ovate-elliptic, membranaceous leaves, with three main veins with prickle-like structures on the abaxial face. S. polyantha leaves showed ovateelliptic, coriaceous leaves, with three main veins, translucent secondary veins and no prickle-like structures. A seedling identification key was elaborated based on morphological characteristics.
Key words: greenbrier, leaf morphology, light, medicinal plant, temperature.
Smilax species, also known as “salsaparrilha” (Greenbrier), are usually used by people due to their anti-rheumatic properties attributed to infusions of their roots and rhizophores. These species are widely distributed in Brazil and their commercial trades are carried out by the extractivism of native species (Andreata 1997).
Studies about the germination of species are important tools to help solving taxonomical and phylogenetical questions and to help the understanding of morphology using ontongeny (Andreata & Pereira 1990, Oliveira 2001). Additionally, the establishment of conditions for germination is of crucial importance for the production and trade of viable seeds.
Several authors carried out studies about germination of Smilax species (Holm 1890, Andreata 1980, Pogge & Bearce 1989, Andreata & Pereira 1990, Barnea et al. 1991, Andreata & Menezes 1999, Rosa & Ferreira 1999, D’Antuono & Lovato 2003, Santos et al. 2003, Palhares et al. 2009, Martins et al. 2011). These studies tested different treatments and they found different results in relation to responses to light, dormancy break and germination rates. However, Smilax is a dioecious genus and therefore the plants are generally markedly present in the ecosystems, their seeds are difficult to germinate.
Seedlings of monocotyledons have a complex structure. Unfortunately, the literature contains many superficial seedling descriptions, together with an inexact and often incorrect or misleading terminology (Tillich 2007). There is a lack of studies about the morphology of seedlings of Smilacaceae family. One of the few studies described the morphology of seedlings of five different Brazilian species of Smilax considering the pos-germination period and the appearance of the first eophyll (Andreata & Pereira 1990). Martins et al. (2011) presented seed germination and seedling and young plant morphology of Smilax polyantha and discusses the terminology of the underground system.
Therefore, the aim of this study was to provide information about the germination and seedling morphology of four species of Smilax known as “salsaparrilha”: Smilax brasiliensis Sprengel, S. campestris Grisebach, S. cissoids Martius ex Grisebach and S. polyantha Grisebach. Moreover, a seedling identification key was elaborated based on morphological traits, which can help further researches in identifying Smilax seedlings.
Materials and methods
Fruit sampling: Fruits were sampled randomly from individuals in different plant populations and occurrence locations (Table 1). The plant samples were registered and added to the plant collection of the Herbarium (ESA) of the Escola Superior de Agricultura ‘‘Luiz de Queiroz’’ of the Universidade de São Paulo.
Germination experiments: We tested two different factors affecting germination rates: two temperature conditions (30°C and 20-30°C) and two light regimes (eight hours of light, and total absence of light). For each treatment, we used four replicates, with 25 seeds each. The seeds were previously selected in order to eliminate grossly damaged or malformed seeds. All seeds were transferred to germination box (11x3cm), with 292g of sterilized sand and 45mL of water. Observations were carried out weekly during 30 weeks. Germination was confirmed by the emission of aboveground parts, since seeds were buried (1.0cm deep) in sand.
We performed analysis of variance using randomization tests to test differences between treatments for each species. We chose this method, since there is no assumption of normal distribution of the data (Manly 2007). We also used Euclidean distance as resemblance measure (10 000 iterations). The statistical analyses were performed by the software MULTIV (Pillar 2005).
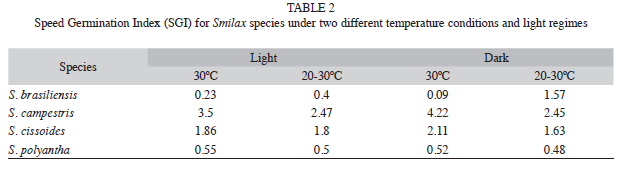
At the end of the experiments, the SGI (Speed Germination Index) was calculated according to Maguire (1962). The seeds that did not germinate had their viability tested by performing a modified tetrazolium test according to the methodology adopted by Cury et al. (2010). Seeds were also considered not viable when the tissue was damaged and/or altered by fungi, compromising the seedling development.
Seedling morphology: Ten seedlings of each study species were analysed concerning their morphology. They were photographed with a digital camera during the following development stages (Andreata & Pereira 1990): early seedling development, seedlings with the first expanded eophyll, young plants with five eophylls and one-year old plants. The following traits were observed (Andreata & Pereira 1990): number of hyaline cataphylls formed in the primary axis, characteristics of the cotyledonary sheath, number and shape of young leaves, aerial stem ramification, formation of underground stems and distinction among primary and adventitious roots, as well as their establishment.
Results
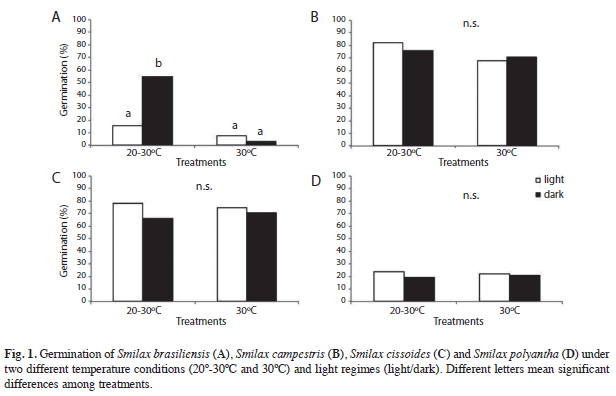
Germination experiments: The results showed that seeds of S. brasiliensis (Fig. 1A) showed an interaction between light and temperature (p=0.0002). The highest values of germination were reached under conditions of 20-30ºC with the absence of light (55%, p≤0.05). On the other hand, significant differences between treatments of temperature could be found for S. campestris (p=0.04, Fig. 1B), but not for light conditions (p=0.81). The 79% of seeds germinated in the treatment of 20-30ºC.
No significant differences between treatments for both light and temperature conditions were found for S. cissoides and S. polyantha (p=0.89 and p=0.06 for S. cisscoides, and p=0.99 and p=0.44 for S. polyantha Fig. 2C, D). The first species showed high germination rates (between 66 and 78%), whilst the second one had the lowest germination rates (19-24%) among all study species.
The first species to germinate was S. campestris (third week). This species also showed the highest values of SGI (2.45 to 4.22, Table 2). S. cissoides began to germinate at the fourth week and it had SGI values from 1.63 to 1.86. Finally, seeds of S. brasiliensis and S. polyantha had the lowest SGI values (0.09 to 1.57 and 0.5, respectively).
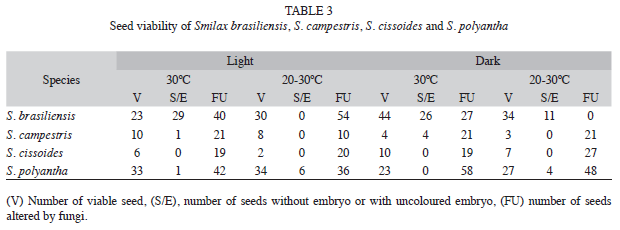
The tetrazolium tests showed that ca. 36-58% of the total number of seeds of S. polyantha had some kind of tissue alteration due to the presence of fungi at the end of the experiments (Table 3).
The other species showed lower percentages of fungi infection. S. campestris and S. cissoids had about 20% of infected seeds. Seeds of S. brasiliensis showed variation according to the treatment: 5% of all seeds were infected by fungi for the treatment 20-30ºC (dark), whilst this percentage increased to 50% for 20-30ºC (light). S. campestris and S. cissoides seeds had the lowest percentage of viable seeds at the end of the experiments (2-10%). S. brasiliensis had 23-44% of viable seeds and S. polyantha between 23-34% (Table 3).
Morphological analysis: The primary axis was formed by three (rarely four) translucent cataphylls before the formation of the first eophyll, which occurred 15-30 days after the germination. The cotyledonary sheath had at first a yellowish colour and a soft consistency (Fig. 2 A, C), surrounding completely the reduced epicotyl. Further at the development phase, the sheath became brownish and chaffy. The morphology of the first eophyll did not differ among the study species, showing ovateshape membranaceous leaves with convex apex, margin entire, rounded base, acrodrome leaf venation base and three main veins (Fig. 2 B, D). This first eophyll dried out when the plant had five leaves and thus, the eophyll was not visible in one-year old plants (Fig. 2 E, 3).
The completely expanded leaves in oneyear old plants had distinct characteristics among the study species. The largest leaf of S. brasiliensis was 8x7cm, ovate-shaped, with a coriaceous consistency and three main veins. The midrib had prickle-like structures on the abaxial face (Fig. 2 F, G). Leaves of S. campestris had 4.5x2cm, oblongate-elliptic shape, coriaceous consistency and only one midrib, also with prickle-like structures on the abaxial face. However, prickle-like structures were also observed on the leaf margins (Fig. 2 H, I).
S. cissoides showed ovate-elliptical shaped leaves of 7x4cm, with a membranaceous consistency and the presence of three main veins with prickle-like structures on the abaxial face (Fig. 2 J, K). Finally, S. polyantha had coriaceous leaves, 7x4.5cm, with ovate-elliptical shape, three main veins and the absence of pricklelike structures. On the margin of the leaves, the vein was translucent, as well as the secondary veins. This characteristic was not observed by the other study species (Fig. 2 L, M).
The aerial caulinar ramification began after the formation of the five first leaves (five months after germination).
The one-year old individuals of S. polyantha and S. cissoides showed ca. five aerial stems, whilst S. campestris and S. brasiliensis had ca. seven stems (Fig. 3). The length of the stems by S. cissoides and S. campestris could be up to 1m, whilst stems of S. polyantha and S. brasiliensis were not larger than 0.5m (0.3 and 0.4, respectively). The average width of stems did not vary among study species (1-2mm, Fig. 3). All study species showed prickle-like structures at the internode. However, they were more abundant by S. campestris and S. cissoides.
Each aerial branch was originated from an axillary bud present on the base of the early aerial stem. Therefore, the development of the underground system began from the fifth eophyll, which can be verified by the presence of reduced knotty from the early development phases of the underground systems on one-year old plants (Fig. 3).
The primary roots of S. cissoides and S. brasiliensis could not be differentiated from the adventitious roots in the early stages of seedling development (Fig. 2 C). The primary root was differentiated in S. polyantha until the formation of the first eophyll. In S. campestris, the primary root remained differentiated until the formation of the fifth eophyll. After one year, all study species showed well-developed adventitious radicular system, composed by white and brownish roots (Fig. 3).
Discussion
The germination dawn, as well as the percentage of germination are factors that can be related to the time needed by the seeds for embryo maturation, absence and presence of light and to scarification (Andreata & Pereira 1990). According to Rosa & Ferreira (1999), the highest germination rates and more rapid germination were verified in dark conditions. Pogge & Bearce (1989) indicated that the increase in light intensity as a factor can both enhance germination rates and reduce the time needed for germination to begin.
Light was an important factor affecting the germination of only one of the study species, S. brasiliensis, which usually occurs in open areas. On the other hand, the remaining study species, that occurs on forest edges, were not affected by this factor. Pogge & Bearce (1989) tested the different light conditions for the germination of two Smilax species. They observed that the species that occurred in open areas (S. glauca) needed more light to germinate than S. rotundifolia, which could be found in shaded areas. According to the authors, light seemed to be a limiting factor for the germination of S. glauca.
The high germination rates of treatments with temperatures 20-30º and 30ºC for S. campestris and S. cissoids, reinforce the idea that the low germination rates found for Smilax polyantha were not only related to temperature variation, substrate or light, but to an intrinsic factor of its metabolism and it thus can be an adaptation strategy of this species to the environment.
The germination rates of Smilax campestris, S. cissoides and S. brasiliensis found in this study are close to the ones obtained by Rosa & Ferreira (1999). Their results showed that 71% of total seeds of Smilax campestris germinated in the presence of light and temperatures of 30ºC, whilst 51% germinated using temperatures of 25-35°C. Pogge & Bearce (1989) found similar results for germination under constant temperatures (22°C) for Smilax rotundifolia (95%) and S. glauca (82%).
Rosa & Ferreira (1999) showed that germination began 30 days after experiments, in the presence of light, and after 15 days in dark conditions. Seeds of S. rotundifolia germinated after two months, whilst seeds of S. glauca began to germinate only after ten months (Pogge & Bearce 1989). The process of germination can vary between 39 and 93 days for seeds of Smilax rufescens, S. elastica, S. syphylitica, S. quinquenervia and S. syringoides (Andreata 1980). Our study showed the begin of germination after 22 days for S. campestris and 29 days for S. cissoides, confirming thus, the highest Speed Germination Indices for both species. Nevertheless, S. brasiliensis and S. polyantha showed the lowest values of Speed Germination Indices, beginning to germinate only after 36 days.
We conducted preliminary tests to break the dormancy of seeds of Smilax polyantha collected in 2004, as described by several authors (Andreata 1980, Pogge & Bearce 1989, D’Antuono & Lovato 2003, Santos et al. 2003, Shao et al. 2006). We used chemical scarification (sulfuric acid) and mechanical (sandpaper), heat shock and gibberillic acid (GA3), following the methodology described by Santos et al. (2003). However, no seeds germinated after the treatments described above. According to Shao et al. (2006) the main cause of dormancy of Smilax riparia was post-maturation of seed embryo. Additionally, we observed that seeds put to germinate using filter paper were more contaminated by fungi than the ones in vermiculite. These preliminary tests contributed to the further performance of experiments, because we did not scarified seeds before experiments and was used sand as substrate.
Seeds of study species showed hypogeal germination, the same one found in other species of Smilacaceae (Tillich 2007), and cryptocotylar, with the formation of hyaline cataphylls, as described by Duke (1965).
According to Andreata & Pereira (1990), the main root is difficult to be identified since the beginning of the development for Smilax elastica, S. quinquenervia, S. rufescens, S. syphylitica and S. syringoides because the adventitious roots can be confused with the primary ones. The presence of the membranaceous cotyledonar sheath and differentiated leaf traits among the species observed in the present study were also verified by these authors. However, they stated that the first eophyll can differentiate the seedlings and this fact was not observed in our study, since the eophyll of the study species did not show significant differences.
Guaglianone & Gattuso (1991) and Andreata (1997) discuss that the detailed characterization of the leaves showed great taxonomic value for distinguishing species of the genus Smilax. Martins et al. (2010) observing the anatomy of the vegetative underground organs of six species of Smilax L. concluded that the morphology of this organs do not allow to distinguish among the studied species. The authors suggested that species still need to be identified by leaf morphology as demonstrated in the present study.
Martins et al. (2011) showed that the horizontal growth and subsequent thickening of the underground organ of S. polyantha depend on the axillary buds development from the basal nodes of previous branches. According to them all adventitious roots originate from the underground stem called rhizophore. The same development of underground stem and root formation were observed in the present study.
Holm (1890) observed that the number of ramifications of Smilax rotundifolia and S. glauca could determine the age of plants, since the formation of a new stem could be observed every year. We did not find this relation for the study species, since after one year of development, individuals of S. polyantha and S. cissoids showed five aerial stems, whilst plants of S. campestris and S. brasiliensis had ca. seven stems.
Acknowledgments
We thank to FAPESP (São Paulo Council for Research) - BIOTA Program (Project Proc. nº 05/54984-5 and Proc. nº 05/58964-9), for the financial support of this research and doctoral scholarship to Aline Redondo Martins and CNPq for research grants. We also thank the technical laboratory support of Marli Kasue Misaki Soares and Helena Pescarin Chamma. This study is part of the PhD thesis of Aline Redondo Martins (Programa de Pós-graduação em Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Brazil).
Identification key for one-year old plants of Smilax brasiliensis, S. campestris, S. cissoides and S. polyantha
References
Andreata, R.H.P. 1980. Smilax Linnaeus (Smilacaceae). Ensaio para uma revisão das espécies brasileiras. Arq. Jard. Bot. Rio de Janeiro 24: 179-301. [ Links ]
Andreata, R.H.P. 1997. Revisão das espécies brasileiras do gênero Smilax Linnaeus (Smilacaceae). Pesqui. Bot. 47: 7-244. [ Links ]
Andreata, R.H.P. & T.S. Pereira. 1990. Morfologia das plântulas de algumas espécies de Smilax L. Pesqui. Bot. 41: 7-24. [ Links ]
Andreata, R.H.P. & N.L. Menezes. 1999. Morfoanatomia do embrião, desenvolvimento pós-seminal e origen do rizóforo de Smilax quinquenervia Vell. (Smilacaceae). Bol. Bot. Univ. São Paulo 18: 39-51. [ Links ]
Barnea, A., Y. Yom-Tov & J. Friedman. 1991. Does ingestion by birds affect seed germination? Funct. Ecol. 5: 394-402. [ Links ]
Cury, G., A.D.L.C. Novembre & B. Appezzato da Glória. 2010. Seed germination of Chresta sphaerocephala DC. and Lessingianthus bardanoides (Less.) H. Rob. (Asteraceae) from Cerrado. Braz. Arch. Biol. Technol. 53: 1299-1308. [ Links ]
D’Antuono, L.F. & A. Lovato. 2003. Germination trials and domestication potential of three native species with edible sprouts: Ruscus aculeatus L., Tamus communis L. and Smilax aspera L. Acta Hortic. 598: 211-218. [ Links ]
Duke, J.A. 1965. Keys for the identification of seedlings of some prominent woody species in eight forest types in Puerto Rico. Ann. Mo. Bot. Gard. 52: 314-350. [ Links ]
Guaglianone, E.R. & S. Gattuso. 1991. Estudios taxonómicos sobre el género Smilax (Smilacaceae). Bol. Soc. Argent. Bot. 27: 105-129. [ Links ]
Holm, T. 1890. Contributions to the knowledge of the germination of some North American plants. Mem. Torrey Bot. Club 2: 57-108. [ Links ]
Maguire, J.D. 1962. Speed of germination-aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 2: 176-177. [ Links ]
Manly, B.F.J. 2007. Randomization, bootstrap, and Monte Carlo methods in biology. Chapman & Hall/CRC, Boca Raton, Florida, USA. [ Links ]
Martins, A.R., N. Pütz, A. Soares, A.B. Bombo & B. Appezzato-da-Glória. 2010. New approaches to underground systems in Brazilian Smilax species (Smilacaceae). J. Torrey Bot. Soc. 137: 220-235. [ Links ]
Martins, A.R., N. Pütz, A.D.L.C. Novembre, S.M.S. Piedade & B. Appezzato-da-Glória. 2011. Seed germination and seedling morphology of Smilax polyantha (Smilacaceae). Biota Neotrop. 11. [ Links ]
Oliveira, D.M.T. 2001. Morfologia comparada de plántulas e plantas jovens de leguminosas arbóreas nativas: espécies de Phaseoleae, Sophoreae, Swartzieae e Tephrosieae. Rev. Bras. Bot. 24: 85-97. [ Links ]
Palhares D., M.A. Tiné, D. Vinha, C.E.S. Silveira & L.B.P. Zaidan. 2009. Studies on the seeds of Smilax goyazana A.DC. (Smilacaceae). Phyton (Horn, Austria) 49: 117-130. [ Links ]
Pillar, V.D. 2005. MULTIV: Multivariate exploratory analysis, randomization testing and bootstrap resampling. Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil. [ Links ]
Pogge, F.L. & B.C. Bearce. 1989. Germinating common and cat greenbrier. Tree Planters’ Notes 40: 34-37. [ Links ]
Rosa, S.G.T. & A.G. Ferreira. 1999. Germination of medicinal plant: Smilax campestris Griseb. (Salsaparrilha). Acta Hortic. 502: 105-111. [ Links ]
Santos, M.R.A., R. Paiva, G.A.C. Gomes, P.D.O. Paiva & L.V. Paiva. 2003. Estudos sobre superação de dormência em sementes de Smilax japecanga Grisebach. Cienc. Agrotecnol. 2: 319-324. [ Links ]
Shao, Mei-Ni, X.S. Jun, L. Zuo, Q. Bo & F.H. Yan. 2006. Preliminary study on the germination character of Smilax riparia seed. J. Anhui Agric. Sci. 23: 6166-6167. [ Links ]
Tillich, H.J. 2007. Seedling diversity and the homologies of seedlings organs in the order Poales (Monocotyledons). Ann. Bot. 100: 1413-1429. [ Links ]
*Correspondencia a: Aline Redondo Martins: Institute of Biology, Universidade Estadual de Campinas, 13083-970, Campinas, SP, Brazil; aline_martins_botanica@yahoo.com.br
Anielca Nascimento Soares, Aline Bertolosi Bombo & Beatriz Appezzato da Glória: Department of Biological Sciences, Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de São Paulo,13418-900, Piracicaba, SP, Brazil; anielcasoares@hotmail.com, aline_bbombo@hotmail.com, bagloria@esalq.usp.br
Alessandra Fidelis: Department of Ecology, Universidade de São Paulo, 05508-900, São Paulo, Brazil; fidelis@usp.br
Ana Dionisia da Luz Coelho Novembre: Department of Crop Science, Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de São Paulo,13418- 900, Piracicaba, SP, Brazil; adlcnove@esalq.usp.br
1. Institute of Biology, Universidade Estadual de Campinas, 13083-970, Campinas, SP, Brazil; aline_martins_botanica@yahoo.com.br
2. Department of Biological Sciences, Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de São Paulo,13418-900, Piracicaba, SP, Brazil; anielcasoares@hotmail.com, aline_bbombo@hotmail.com, bagloria@esalq.usp.br
3. Department of Ecology, Universidade de São Paulo, 05508-900, São Paulo, Brazil; fidelis@usp.br
4. Department of Crop Science, Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de São Paulo,13418- 900, Piracicaba, SP, Brazil; adlcnove@esalq.usp.br
Received 26-I-2011. Corrected 05-V-2011. Accepted 07-VI-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}