










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Ovarian folliculogenesis in collared peccary, Pecari tajacu (Artiodactyla: Tayassuidae)
Diva Anelie Guimarães1*, Sylvia Cristina Garcia de Garcia1, Maria Auxiliadora Pantoja Ferreira1, Suleima do Socorro Bastos da Silva1, Natália Inagaki de Albuquerque2* & Yvonnick Le Pendu3*
*Dirección para correspondencia
Abstract
The sustainability and production of collared peccary (Pecari tajacu) has been studied in the last few years; however, further information on its reproduction is necessary for breeding systems success. Understanding folliculogenesis aspects will contribute to effective reproductive biotechniques, which are useful in the preservation and production of wildlife. The aim of this study was to evaluate the ovarian folliculogenesis in collared peccary. Ovaries from six adult females of collared peccary were obtained through ovariectomy and analyzed. These were fixed in aqueous Bouin’s solution and sectioned into 7μm slices, stained with hematoxilin-eosin and analyzed by light microscopy. The number of pre-antral and antral follicles per ovary was estimated using the Fractionator Method. The follicles, oocytes and oocyte nuclei were measured using an ocular micrometer. Results showed that the length, width, thickness, weight, and the gross anatomy of the right and left ovaries were not significantly different. However, the mean number of corpora lutea was different between the phases of the estrous cycle (p<0.05), with the highest mean in the luteal phase. Primordial follicles were found in the cortex; the oocytes were enveloped by a single layer of flattened follicular cells. In the primary follicles, proliferation of the follicular cells gave rise to cuboidal cells (granulosa cells). The secondary follicle was characterized by two or more concentric layers of cuboidal cells (granulosa), beginning of antrum formation, and the presence of pellucid zone and theca cells. Antral follicles were characterized by a central cavity (antrum), the presence of cumulus oophorus and theca layers (interna and externa). In the right ovary, the values of the primordial and primary follicles were similar, but significantly different from the secondary ones (p<0.05). In the left ovary, significant differences were observed between all follicles in the follicular phase (p<0.05); the mean number of primordial and primary follicles was similar in the luteal phase. The mean number of pre-antral follicles and antral follicles in the follicular phase was higher in the left ovary (p<0.05). The mean number of antral follicles in the luteal phase was similar in both ovaries. We also found significant differences in mean diameter of preantral follicles, oocyte, granulosa layer and oocyte nucleus during the estrous cycle. In the antral follicles a significant difference was observed only in follicular diameter (p<0.05). The predominance of active primordial and primary follicles was found in both phases; otherwise the secondary follicles and antral follicles showed a high degree of degeneration. The results obtained in the present work will strengthen the development of biotechnology programs to improve the productive potential and conservation of the collared peccary.
Key words: ovary, morphology, histology, antral follicles, pre-antral follicles, collared peccary.
Resumen
La sustentabilidad y la producción de pecarí de collar (Pecari tajacu) han sido estudiados en los últimos años, sin embargo, más información sobre su reproducción es necesaria para el éxito de los sistemas de crianza . La comprensión de los aspectos relacionados con la foliculogénesis contribuirá con la aplicación de biotécnicas de reproducción, las cuales son útiles en la preservación y la producción de la vida silvestre. El objetivo de este estudio fue obtener datos sobre la población folicular del ovario de pecarí de collar. En relación con la población folicular en el ovario derecho, los valores de los folículos primordiales y primarios fueron similares, pero se observó que había una diferencia significativa (p<0.05) con el secundario. En el ovario izquierdo, la fase folicular presentó diferencias significativas (p<0.05) entre todos los folículos, y en la fase lútea el número medio de folículos primordiales y primarios fueron similares. Ahora bien, con respecto a la población de folículos antrales, en la fase folicular, se observaron diferencias significativas entre los ovarios (p<0.05), y de forma similar en la fase lútea. También se encontraron diferencias significativas en el diámetro medio de los folículos preantrales, oocitos, la capa granulosa y el núcleo del oocito durante las fases del ciclo estral. Asimismo, en los folículos antrales se observó diferencia estadísticamente significativa (p<0.05) en el diámetro folicular. En ambas fases del ciclo estral, se encontró el predominio de folículos primordiales y primarios en desarrollo, por otro lado, los folículos secundarios y los folículos antrales mostraron un alto grado de degeneración. Los resultados aquí presentes son necesarios para el desarrollo de los programas de mejoramiento y conservación.
Palabras clave: ovario, morfología, histología, folículos antrales, folículos preantrales, pecarí de collar.
The collared peccary (Pecari tajacu) is a neotropical mammal that has biological characteristics different from other suids. They inhabit different ecosystems from the South of the United States of America to the North of Argentina (Sowls 1997). The collared peccary is extensively hunted for its meat and leather in some countries of Latin America and mainly in the Amazon region (Bodmer & Lozano 2001, Mayor et al. 2007, Fang et al. 2008). Presently, it is considered a wild animal with potential to be raised commercially (Nogueira-Filho & Nogueira 2004).
The sustainability and production handling of the species has been studied in the last years (Barbella 1993, Sowls 1997, Gottdenker & Bodmer 1998, Guimarães et al. 2004, Nogueira-Filho & Nogueira 2004, Le Pendu et al. 2006, Mayor et al. 2006, Albuquerque et al. 2010, Costa et al. 2010, Mayor et al. 2010). Nevertheless, further information on its reproduction is necessary for the success of breeding programs.
The cryopreservation of gametes, associated with assisted reproduction, is uncommon in wild animals (Fickel et al. 2007) due to the lack of studies on the reproductive morphophysiological mechanisms of this species.
Folliculogenesis, the process of formation, activation, growth, and maturation of follicles, occurs in the ovary cortex, starting with the formation of the primordial follicle and continuing to the pre-ovulation stage (Hafez 2004). Nevertheless, an in-depth understanding of the aspects related to this process will contribute to the application of biotechniques like in vitro fecundation and the manipulation of oocytes from pre-antral follicles, which are useful in the preservation of wildlife, as well as for animal production. The main goal of this study was to investigate the basic aspects of folliculogenesis in collared peccary.
Materials and methods
This experiment was performed in the Senador Álvaro Adolpho Animal Research Unit (Embrapa Eastern Amazon) in Belém, Pará State, Brazil, (1º28’ S - 48º27’ O). The animals were kept in family groups in 36m2 paddocks and fed with commercial swine feed (calorific supply of 2 500kcal per kg and protein level of 14%). Collared peccary females aged from 12 to 24 months were used; three females were subjected to ovariectomy in the follicular phase and the other three in the luteal phase. They were anesthetized with ketamine chloridate (5mg/kg) and 0.2% acepromazine (0.2mg/kg), administered intramuscularly before ovariectomy. The experiments performed in this study were approved by the Ethical Committee for Animal Research, Federal University of Pará (Authorization number CEPAE-ZOO001-08), and were conducted in compliance with current Brazilian and international laws on the manipulation of animals for research purposes. The ovaries were measured with a pachimeter and weighed in an analytical balance. The hemorrhagic body, corpora lutea, corpus albicans and antral follicles were counted with the aid of a magnifying glass.
All the samples were fixed in aqueous Bouin’s solution, dehydrated with increasing ethanol concentration, infiltrated and embedded in paraffin, cut in 7μm slices, stained with hematoxilin-eosin, and analyzed by light microscopy. The ovary follicles observed in different stages of maturation (pre-antral: primordial, primary and secondary; antral: tertiary) were classified as active or atretic, according to Schillo (2009).
Ovary follicles were counted in each phase of the cycle (follicular and luteal). The preantral follicles were counted in every 30th mounted section, and the antral follicles, in every 40th section. Only follicles which presented visible oocyte nuclei were considered. Follicle populations were estimated by the Fractionator Method (Gundersen et al. 1988) using the formula: number of follicles x number of sections. The results of the counting of the analyzed sections were added, giving an estimate of the follicle population of each ovary.
The ovary follicles were measured using an ocular micrometer coupled to a light microscope (10x and 40x). Five each of pre-antral and antral follicles were selected in each phase of the oestrous cycle for measurement of the follicle, oocyte, and oocyte nucleus diameter. The size of the germinal cell layer (GCs) was measured for all pre-antral follicles by subtracting the oocyte diameter from the follicle diameter.
The results are given as means (±standard deviation). Tukey’s test was used to compare the follicle dimensions and population estimates of the right and the left ovaries, according to the follicular development. Kruskal- Wallis’ test was used to compare the diameters of the oocytes, oocyte nuclei, the follicles and the layer of GCs of the different categories of pre-antral follicles. All the analyses were performed at 5% level of confidence using the software BioEstat 5.0 (Ayres et al. 2007).
Results
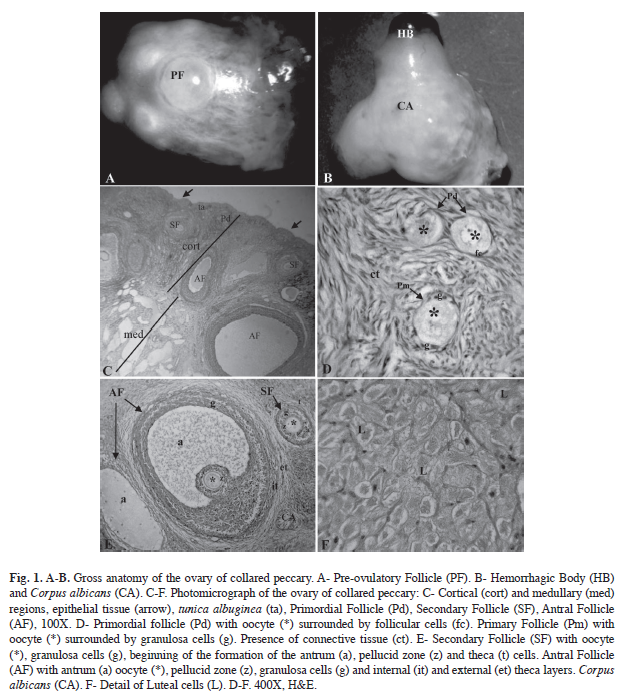
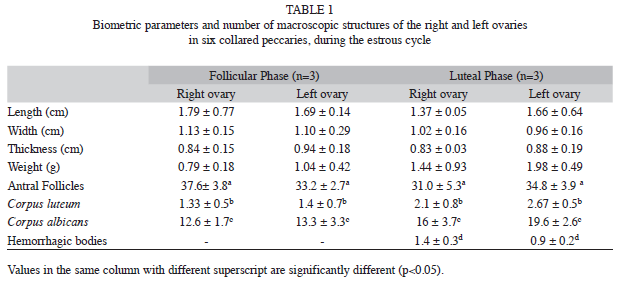
The analyzed collared peccary ovaries were partially covered by a bursa ovarica, had an oval shape, and an irregular surface (Fig. 1A, 1B). The length, width, thickness, weight, and number of macroscopic ovarian structures (antral follicles, corpora lutea, corpus albicans and hemorrhagic body) of both ovaries (right and left) were not statistically different. However, the mean number of each of the ovary structures was different (Table 1). The mean number of corpora lutea was significantly different between the oestrous cycle phases, being highest in the luteal phase (2.67±0.52).
The ovary surface had a cover of simple cuboidal epithelium, which cells, in the presence of pre-ovulatory follicles, corpora lutea or hemorrhagic body, became pavimentous cells. Right bellow the epithelium, there was a layer of dense connective tissue, the tunica albuginea. In the cortical region, follicles in different phase of development were observed, as well as corpora lutea, hemorrhagic body, corpus albicans and loose connective tissue. The medullar region consisted of densely vascularized loose connective tissue (Fig. 1C).
The primordial follicles (Fig. 1D) were formed by oocytes surrounded by a layer of follicular cells (pavimentous cells), and the primary follicles, by a single layer of granulosa cells (pavimentous and/or cuboidal). The secondary follicles (Fig. 1E) had from two-four layers of granulosa cells, the beginning of the formation of the antrum, pellucid zone, and thecas (external and internal). The antral follicle oocytes (Fig. 1E) were surrounded by four or more layers of granulosa cells, antrum, and internal (cellular) and external (fibrous) thecas.
The statistical data for pre-antral ovary follicles of the right and left ovaries in the follicular and luteal phases are shown in Table 2. During the oestrous cycle, the right ovary mean values were similar to those of the primary follicles, and statistically different (p<0.05) from the secondary follicles. The left ovary mean values were all statistically different in the follicular phase while the mean numbers of primordial and primary follicles were similar in the luteal phase. In both phases, the mean numbers of primordial and secondary follicles were higher for the left ovary than for the right ovary. The mean number of antral follicles was higher in the left than in the right ovary in the follicular phase (115.3±43.1 and 61.80±45, respectively). In the luteal phase, a mean of 54.1±19.1 antral follicles was observed for the right ovary, which was similar to that of the left ovary, with 65.3±30.5 antral follicles.
There was no difference (p<0.05) in the average number of primary follicles (303.6±122.1 and 237.0±68.8, respectively) in the follicular and luteal phases; however, the average number of secondary follicles (155.4±55.9 and 78.3±23.3, respectively) and antral follicles (103.5±45.6 and 60.0±26.1, respectively) was different (p<0.05) in both phases.
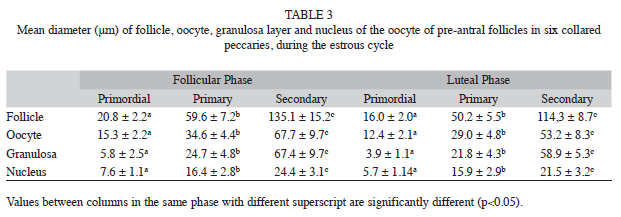
Statistical differences were also observed in the mean increase in diameter of pre- antral follicles, oocyte, the oocyte nucleus and the granulosa layer (Table 3) during the oestrous cycle phases. Only the mean follicle diameter (mm) was statistically different (p<0.05) in the antral phase (821±110.6 for follicular phase and 626.0±161.4 for luteal phase); the mean oocyte diameter (mm) of antral follicles in the follicular and luteal phases was 75.3±10.0 and 62.3±6.4, respectively; and the mean nucleus diameter (mm) was 37.8±2.7 and 30.4±4.4, respectively, for both phases.
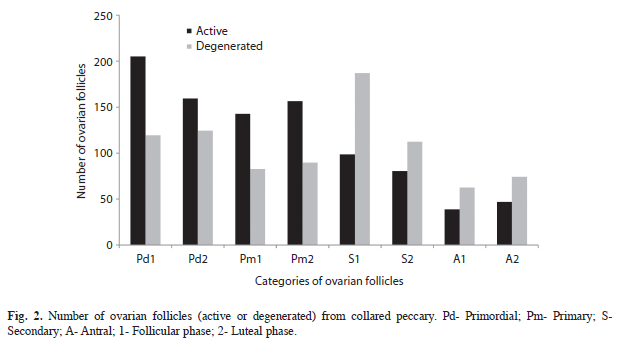
In both phases of the oestrous cycle, primordial and developing primary follicles predominated, while secondary and antral follicles had a high degree of degeneration (Fig. 2). The corpus albicans (Fig. 1E) was characterized by extensive fibrosis, while the hemorrhagic body was characterized by the formation of a blood clot. The corpora lutea were observed in both phases of the oestrous cycle, and with regard to the cell morphology, they were functional (Fig. 1F) or remanent. Accessory corpora lutea were also observed, being characterized by degenerative oocytes.
Discussion
These results show that the ovarian histological constitution of collared peccary is similar to that of most mammals. The ovary length, width, and thickness measurements corroborated the reports of Santos et al. (2000). However, the ovary weight found in this work was smaller than that described by Paula et al. (2003), which may be due to factors like age, reproductive phase, and nutritional state of the females used.
The mean number of corpora lutea was similar to that reported by Mayor et al. (2006) in hunted wild females, thus indicating that captivity conditions apparently do not interfere with the ovulation rate of this species. Most of the functional structures were observed in the cortical region of the ovary. Developing and degenerating follicles were identified, suggesting that follicle genesis is a dynamic process, with continuous cell growth and death. In this study, the primordial and primary follicles were agglomerated in the peripheral regions of the cortex, which agrees with Barbella (1993).
Ovarian follicular quantification varies highly among species. In buffaloes (Bubalus bubalis), the average number of primordial, primary, and secondary follicles is 3 518.98±1 988.37, 14 672.03±12 263.76 and 1 628.29±1 182.24, respectively (Mondadori et al. 2010), while in mice (100 days of age) it is 2 227±101, 265±32 and 79±5.6, respectively (Myers et al. 2004). Gosden & Telfer (1987) observed that in some species the number of primordial follicles varied allometrically with both body weight and maximum life expectation, and the difference in the follicular number between species could be a strategy, which guarantees fecundity throughout most of the lifespan. The average number of primordial follicles on peccaries is very low compared to other suiformes, such as the domestic pig (Sus scrofa), which has close to 400 000 (Greenwald & Moor 1989, Hafez 2004). So far, there are no references on similar species to allow comparisons.
Significant differences were observed in the number of secondary and antral follicles between the oestrous phases in collared peccaries. The means numbers were higher in the follicular phase, when the follicle is recruited, selected and becomes dominant. The mean number of primordial follicles was close to that of primary follicles, in contrast to what is generally observed in mammals. This result can be due to the follicle classification used in this work, which included in primary follicles category those with granulosa pavimentous cells or cuboidal shape, thus increasing the mean values in this category.
In this work, during the follicular phase, 103.5±45.6 antral follicles were observed in the ovary of collared peccary. The averages were higher than that observed (30.4±1.7) by Mayor et al. (2006) in the same species; this difference may be related to several factors including the age of the females, nutritional state and the methodology used in both experiments. According to Myers et al. (2004), there may be greater differences in follicular numbers per ovary in mice at similar ages using various methods of study.
Throughout folliculogenesis, over 99% of growing ovarian follicles become atretic (Monniaux et al. 2009). Degeneration predominated in secondary and antral follicles of collared peccaries, which may be related to the follicle growth wave, that is, this species seems to have a first atresia wave and follicle selection, followed by a new atresia and dominance of some follicles. These observations are in agreement with Mayor et al. (2006), who suggested the existence of follicular waves in this species, based on the size and number of antral follicles and corpus luteum. The follicle recruiting, selection, and dominance phases are observed in other species of artiodactyls, such as buffaloes and South American camelids (Baruselli et al. 1997, Riveros et al. 2010, Vaughan 2011); however, their morpho-physiological mechanism is yet little understood (Fortune 1994, Rodgers et al. 1999, Gougeon & Busso 2000, Monniaux et al. 2009).
The statistical difference in the diameter of the follicle structures observed in this study indicates that it can be a valuable parameter for the classification of the distinct follicle categories. Based on the changes in the diameter of the oocyte and nucleus, the granulosa layer, and of the follicle, we can infer that the follicle growth initially occurs by alteration of granulosa cells from pavimentous to cuboidal. Next, there is an increase in the oocyte size, the proliferation of the granulosa layer, and consequently, an increase in the follicle diameter.
According to Monniaux et al. 2009, folliculogenesis is divided into two stages (basal and terminal), and the follicular size is characteristic of the species. Primordial follicles range from 0.03 to 0.05mm, and when they reach 0.2mm in size, the antrum is formed. For these authors, in the terminal stage of folliculogenesis, follicles range from 0.2mm in rodents to 10mm in the ass, the oocyte size increases from 20 to 30 microns; and the mature follicle size ranges from 0.5mm in rodents to 15mm in the ass.
The comparison of the values found in the present work for the mean diameter of primordial follicle (16.0±2.0μm and 20.8±2.2μm), primary follicle (50.2±5.5μm and 59.6±7.2μm), and antral follicle (626.0±161.4μm and 821±110.6μm) are within the same range of values with those reported by Mayor et al. (2006) (12-32μm, 34-190μm and 530-1620μm, respectively); however, the variation found for the mean diameter of secondary follicle (114.3±8.7μm and 135.1±15.2μm) was smaller than that of the small pre-antral follicle (190- 490μm), observed by these same authors. Carvalho et al. (2003) reported the presence of an ovulation fossa; however, in this work the hemorrhagic body was not present in the same location, which indicates the absence of ovulatory fossa in collared peccary. Both cyclic and accessory corpora lutea were observed. The presence of the latter shows the probable need for elevation of plasma progesterone level in the beginning of gestation in this species, thus avoiding embryo death. In conclusion, the results obtained in the present work can be used as parameters in more detailed studies of folliculogenesis in biotechnology programs with the goal of improving the productive potential and conservation of the collared peccary.
Acknowledgments
We thank CNPq (Tayataja Project 471705/03-9) and SECTAM/FUNTEC/Governo do Estado do Pará Project 1025/2004 for supporting this project.
References
Albuquerque, N., D.A. Guimarães, H.L.T. Dias, Y. Le Pendu, P. Kahwage & A.R. Garcia. 2010. Intensive production system of collared peccary (Pecari tajacu) in Brazilian Amazon. Adv. Anim. Biosci. 1: 480-481. [ Links ]
Ayres, M., D. Ayres Jr., L. Ayres & A.A.S. Santos. 2007. BioEstat versão 5.0. Aplicações estatísticas nas áreas das ciências biomédicas. Sociedade Civil Mamirauá, Belém, Pará, Brazil. [ Links ]
Barbella, S.L. 1993. Cambios morfométricos en el aparato reproductivo del báquiro de collar (Tayassu tajacu) entre el nacimiento y los doce meses de edad. Rev. Fac. Agron. 19: 153-165. [ Links ]
Baruselli, P.S., R.G. Mucciolo, J.A. Visintin, W.G. Viana, R.P. Arruda, E.H. Madureira, C.A. Oliveira & J.R. Molero-Filho. 1997. Ovarian follicular dynamics during the estrous cycle in buffalo (Bubalus bubalis). Theriogenology 47: 1531-1547. [ Links ]
Bodmer, R. & E.P. Lozano. 2001. Rural development and sustainable wildlife use in Peru. Conserv. Biol. 15: 1163-1170. [ Links ]
Carvalho, A.F., M.C. de Lima, T.C. dos Santos, M. Bonatelli, M.A. Miglino, V.Y. Samoto, M.F. Oliveira, C.E. Ambrósio, F.T. Pereira & J.F. Martins. 2003. Comparação morfológica dos ovários de catetos e porca (Tayassu tajacu e Sus scrofa). In 4º Encontro de produção acadêmica da FIFEOB. São João da Boa Vista, São Paulo, Brazil. [ Links ]
Costa, G.M.J., M.C. Leal, J.V. Silva, A.C.S. Ferreira, D.A. Guimarães & L.R. França. 2010. Spermatogenic cycle length and sperm production in a feral pig species (collared peccary, Tayassu tajacu). J. Androl. 31: 221-230. [ Links ]
Fang, T.G., R.E. Bodmer, P.E. Puertas, P.A. Mayor, P.P. Peña, R.A. Villanes & D.T.S. Hayman. 2008. Certificación de pieles de pecaríes en la Amazonía Peruana: Una estrategia para la conservación y manejo de fauna silvestre en la Amazonía Peruana. Walter H. Wust, Lima, Perú [ Links ].
Fickel, J., A. Wagcher & A. Ludwig. 2007. Semen cryopreservation and the conservation of endangered species. Eur. J. Wildlife Res. 53: 81-89. [ Links ]
Fortune, J.E. 1994. Ovarian follicular growth and development in mammals. Biol. Reprod. 50: 225-232. [ Links ]
Gosden, R.G. & E. Telfer. 1987. Numbers of follicles and oocytes in mammalian ovaries and their allometric relationships. J. Zool. 211: 169-175. [ Links ]
Gottdenker, N. & R.E. Bodmer. 1998. Reproduction and productivity of white-lipped and collared peccaries in the Peruvian Amazon. J. Zool. London 245: 423-430. [ Links ]
Gougeon, A. & D. Busso. 2000. Morphologic and functional determinants of primordial and primary follicles in the monkey ovary. Mol. Cell. Endocrinol. 163: 33-41. [ Links ]
Greenwald, G.S. & R.M. Moor. 1989. Isolation and preliminary characterization of pig primordial follicles. J. Reprod. Fert. 87: 561-571. [ Links ]
Guimarães, D.A., J.V. da Silva, P. Mayor, Y. Le Pendu, N. Albuquerque & S.L.G. Nogueira-Filho. 2004. Reproductive biology of female collared peccaries (Tayassu tajacu) raised in captivity in Amazon region. In 6º Symposium International sur l’utilisation durable de la faune sauvage - 6th International Wildlife Ranching Symposium. Paris, France. [ Links ]
Gundersen, H., P. Bagge, T.F. Bendtsen, S.M. Evans, L. Korbo, N. Marcussen, A. Moller, K. Nielsen, J.R. Nyengaard, B. Pakkenberg, F.B. Sorensen, A. Vesterby & M.J. West. 1988. The new histology tools: dissector, fractionator, nucleator and point sampled and their use in pathological research and diagnosis. AMPIS 96: 857-881. [ Links ]
Hafez, B. 2004. Reprodução Animal. Manole Ltda, São Paulo, Brazil. [ Links ]
Le Pendu, Y., M.A.M. Lessa & D.A. Guimarães. 2006. Social relationships of collared peccaries (Tayassu tajacu) in an intensive production system. In XXIV Encontro Anual de Etologia. Brasília-DF, Brazil. [ Links ]
Mayor, P., D. Santos-Fita & M. Lopez-Bejar. 2007. Sostenibilidad en la Amazonía y Cría de Animales Silvestres. Centro de Estudios Teológicos de la Amazonía, Iquitos, Perú [ Links ].
Mayor, P., M. Fenech, R.E. Bodmer & M. Lopez-Bejar. 2006. Ovarian features of the wild collared peccary (Tayassu tajacu) from the northeastern Peruvian Amazon. Gen. Comp. Endocr. 147: 268-275. [ Links ]
Mayor, P., R.E. Bodmer, C. Cornejo, H. Gálvez & M. López-Bejar. 2010. Comparative reproductive biology of collared peccary (Tayassu tajacu) populations in the wild and in captivity under semi-extensive conditions in the Northeastern Peruvian Amazon. Suiform Soundings 10: 7-21. [ Links ]
Mondadori, R.G., T.R. Santin, A.A.G. Fidelis, E.P. Porfírio & S.N. Báo. 2010. Buffalo (Bubalus bubalis) pre-antral follicle population and ultrastructural characterization of antral follicle oocyte. Reprod. Dom. Anim. 45: 33-37. [ Links ]
Monniaux, D., A. Caraty, F. Clément, R. Dalbiès-Tran, J. Dupont, S. Fabre, N. Gérard, P. Mermillod, P. Monget & S. Uzbekova. 2009. Développement folliculaire ovarien et ovulation chez les mammifères. Inra Prod. Anim. 22: 59-76. [ Links ]
Myers, M., K.L. Britt, N.G.M. Wreford, F.J.P. Ebling & J.B. Kerr. 2004. Methods for quantifying follicular numbers within the mouse ovary. Reproduction 127: 569-580. [ Links ]
Nogueira-Filho, S.L.G. & S.S.C. Nogueira. 2004. Captive breeding programs as an alternative for wildlife conservation in Brazil, p. 171-190. In K.M. Silvins, R. Bodmer & J.M.V. Fragoso (eds.). People and nature: wildlife conservation in South and Central America. Columbia University, New York, USA. [ Links ]
Bodmer & J.M.V. Fragoso (eds.). People and nature: wildlife conservation in South and Central America. Columbia University, New York, USA.
Paula, V.V., C.E.B. Moura, K.D. Filgueira, R.C. Carvalho, W.P. Costa, J.F.G. Albuquerque, M.F. Oliveira, H.M. Pereira & J.S. Batista. 2003. Morfologia dos órgãos genitais femininos de catetos criados em cativeiro no semi-árido nordestino. Rev. Bras. Reprod. Anim. 27: 276-277. [ Links ]
Riveros, J.L., C. Bonacic, G. Schuler, B. Hoffmann, M.G. Chaves & B. Urquieta. 2010. Ovarian follicular dynamics and hormonal secretory profiles in guanacos (Lama guanicoe). Anim. Reprod. Sci. 119: 63-67. [ Links ]
Rodgers, R.J., T.C. Lavranos, I.L. Wezel & H.F. Irving- Rodgers. 1999. Development of the ovarian follicular epithelium. Mol. Cell. Endocrinol. 151: 171-179. [ Links ]
Santos, T.C., M.A. Miglino, G.V. Machado & W.M. Souza. 2000. Morfologia dos ovários, tubas uterinas e útero em catetos (Tayassu tajacu, Linnaeus, 1758) e queixadas (Tayassu pecari, Link, 1795). Braz. J. Vet. Res. Anim. Sci. 37: 203-209. [ Links ]
Schillo, K.K. 2009. Reproductive physiology of mammals: from farm to field and beyond. Delmar, New York, USA. [ Links ]
Sowls, L.K. 1997. Javelinas and other peccaries: their biology, management and use. University of Arizona, Tucson, Arizona, USA. [ Links ]
Vaughan, J. 2011. Ovarian function in South American camelids (alpacas, llamas, vicunas, guanacos). Anim. Reprod. Sci. 124: 237-243. [ Links ]
*Correspondencia:
Diva Anelie Guimarães: Institute of Biological Sciences, Federal University of Pará, Rua Augusto Corrêa nº01, Guamá, CEP66075-110, Belém, Pará, Brazil; diva@ufpa.br
Sylvia Cristina Garcia de Garcia: Institute of Biological Sciences, Federal University of Pará, Rua Augusto Corrêa nº01, Guamá, CEP66075-110, Belém, Pará, Brazil; sylviacristinabio@yahoo.com.br
Maria Auxiliadora Pantoja Ferreira: Institute of Biological Sciences, Federal University of Pará, Rua Augusto Corrêa nº01, Guamá, CEP66075-110, Belém, Pará, Brazil; auxi@ufpa.br
Suleima do Socorro Bastos da Silva: Institute of Biological Sciences, Federal University of Pará, Rua Augusto Corrêa nº01, Guamá, CEP66075-110, Belém, Pará, Brazil;, suleima_silva@yahoo.com.br
Natália Inagaki de Albuquerque: Brazilian Agricultural Research Corporation-Embrapa Eastern Amazon Region, Travessa Dr. Enéas Pinheiro s/nº, CEP66095-100, Belém, Pará, Brazil; natalia@cpatu.embrapa.br
Yvonnick Le Pendu: Department of Biological Sciences, State University of Santa Cruz, km 16, Rodovia Ilhéus/Itabuna, Salobrinho, CEP45662-900, Ilhéus, Bahia, Brazil; yvonnickuesc@gmail.com
1. Institute of Biological Sciences, Federal University of Pará, Rua Augusto Corrêa nº01, Guamá, CEP66075-110, Belém, Pará, Brazil; diva@ufpa.br, sylviacristinabio@yahoo.com.br, auxi@ufpa.br, suleima_silva@yahoo.com.br
2. Brazilian Agricultural Research Corporation-Embrapa Eastern Amazon Region, Travessa Dr. Enéas Pinheiro s/nº, CEP66095-100, Belém, Pará, Brazil; natalia@cpatu.embrapa.br
3. Department of Biological Sciences, State University of Santa Cruz, km 16, Rodovia Ilhéus/Itabuna, Salobrinho, CEP45662-900, Ilhéus, Bahia, Brazil; yvonnickuesc@gmail.com
Received 10-III-2011. Corrected 20-VI-2011. Accepted 22-VII-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}