










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Bloom of Gymnodinium catenatum in Bahía Santiago and Bahía Manzanillo, Colima, Mexico
Sonia Quijano-Scheggia1*, Aramis Olivos-Ortiz1, José J. Bustillos-Guzmán2*, Esther Garcés3*, Juan H. Gaviño-Rodríguez1, Marco A. Galicia-Pérez1, Manuel Patiño-Barragan1, Christine J. Band-Schmidt4*, Francisco J. Hernández-Sandoval2 & David J. López-Cortés2
*Dirección para correspondencia
Abstract
Gymnodinium bloom events are of concern, since they produce toxins, which have unfavorable consequences to marine ecosystems, human health and the economy. This report describes the physico-chemical conditions that were present during the algal bloom event on May 2010 in Bahía Manzanillo and Bahía Santiago, Colima, Mexico. For this, seawater nutrient analysis, phytoplankton counts, identification, and toxicity tests were undertaken. Nutrients in seawater were determined using colorimetric techniques, the higher concentrations (8.88μM DIN, 0.78μM PO4 and 24.34μM SiO2) were related with upwelling waters that promoted the algal bloom that began after registering the year lowest sea-surface temperature, favoring the rapid growth of G. catenatum (up to 1.02 x107cells/L). Phytoplankton counting was carried out using sedimentation chambers and cells enumerated on appropriated area. The bloom persisted in the bays for approximately two weeks and was associated with toxicity (determined with HPLC) in local oysters (1525.8μg STXeq/100g), and in phytoplankton (10.9pg STXeq/cells) samples. Strong variations in cell toxicity (1.4 to 10.9pg STXeq/cells), most likely reflected the availability of inorganic nutrients. The toxin profile of the phytoplankton samples consisted of 11 toxins and resembled those recorded for several strains of G. catenatum isolated from other coastal areas of Mexico.
Key words: Gymnodinium catenatum, algal bloom, toxicity, upwelling.
Resumen
La proliferación de Gymnodinium son motivo de preocupación, debido a que en algunas circunstancias producen toxinas, que tienen consecuencias desfavorables para los ecosistemas marinos, la salud humana y la economía. Este trabajo describe las condiciones fisicoquímicas presentes durante una proliferación algal detectado en mayo de 2010 en la Bahía de Santiago y Bahía Manzanillo (Colima, México). La proliferación algal inició poco tiempo después de registrarse las temperaturas oceánicas superficiales más bajas del año, las cuales permitieron un aumento de las concentraciones de nutrientes (8.88μM DIN, 0.78μM PO4 and 24.34μM SiO2) que favorecieron el desarrollo de G. catenatum (hasta 1.02 x107cel/L). Esta proliferación se detectó en las bahías durante dos semanas y fue relacionada con toxicidad en ostiones de la localidad (1525.8μg STXeq/100g) y en muestras de fitoplancton (10.9pg STXeq/cel). Fuertes variaciones en la toxicidad de G. catenatum (1.4 a 10.9pg STXeq/cel) pudieron reflejar la disponibilidad de nutrientes inorgánicos. El perfil de toxinas de las muestras del fitoplancton consistieron en 11 toxinas semejantes a las de varias cepas de G. catenatum aisladas de otras áreas de las costas de México.
Palabras clave: Gymnodinium catenatum, proliferaciones algales, toxicidad, surgencias.
Algal blooms occur in coastal waters worldwide and have been mainly ascribed to upwelling systems, oceans fronts, and anthropogenic discharges, among other factors (Hallegraeff 1993, Kudela et al. 2008, Trainer et al. 2009). Bloom events are a source of concern, since under certain circumstances bloom-forming algae produce toxins, with adverse consequences to marine ecosystems, human health, and the economy (Kudela et al. 2008, Shipe et al. 2008).
In coastal areas of the North Pacific, harmful algal blooms (HABs) have become more frequent, most likely associated with biological, physical, and chemical processes related to the California current (Kudela et al. 2010, Trainer et al. 2010). In events dominated by the naked dinoflagellate Gymnodinium catenatum Graham, increases in nitrogen compounds during winter-spring transitions in the water column and periods of upwelling in North Pacific coastal waters, stimulate the production of toxins, including those causing paralytic shellfish poisoning (PSP) toxin (Kudela et al. 2010).
The first record of G. catenatum in Mexican waters was in 1939, in the Gulf of California (Graham 1943). An historical review of the presence of blooms and toxins of G. catenatum in the Mexican Pacific has been done by Band-Schmidt et al. (2010). These authors reported many scarcely recognized relevant studies. Along the central Mexican Pacific coastline, G. catenatum has been found in a 700km area stretching from Bahía Banderas (21°N) to the port of Lázaro Cárdenas (~18°N). Despite that, abundances of this dinoflagellate have been as high as 2.1×105cells/L, there is only one report relating these species with toxicity in bivalves in Bahía Manzanillo, with a high toxicity value of 235.28μg saxitoxin/100g wet weight, in the oyster Crassostrea iridescens during that bloom (González-Chan et al. 2008). Species of dinoflagellates, diatoms and ciliates have been identified as the main phytoplankton components of bloom events. In this case, the species responsible for high-abundance proliferations were: Ceratium divaricatum (4.5×108cells/L), Myrionecta rubra (3×106cells/L), Dictyocha fibula (2.8 ×105cells/L). The presence of G. catenatum in Bahía Manzanillo was reported lately in August 1989 and April 2002 (Ortiz- Lira & Jiménez-Quiroz 2006). Furthermore, Figueroa-Torres & Zepeda-Esquivel (2001) have described G. catenatum blooms at the Internal Port of Manzanillo on April and December 1999 and in March 2000.
The characteristics and toxicity of HABs and the environmental and hydrodynamic conditions that favor these events are largely unknown, due to the lack of a systematic monitoring program in the affected areas. A better understanding of the hydrodynamic conditions present at the time of the bloom will improve our ability to predict these events, and thereby potentially limit their negative consequences. Therefore, the aim of this study was to evaluate the 2010 bloom of G. catenatum in the Bahía Manzanillo and Bahía Santiago.
Material and methods
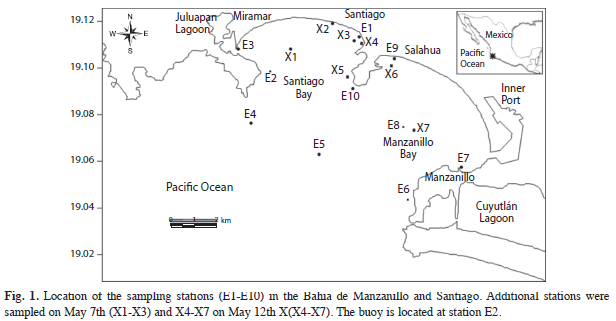
Sampling: The study area included Bahía Manzanillo and Bahía Santiago, in the central zone of the Mexican tropical Pacific (19°05’ N - 104°23’ W) (Fig. 1). Both bays are semicircular in shape and open to the ocean, with a distance of 15km between Punta Carrizal to the North and Punta Ventanas to the South. The bays, from the mouth to the shore, are about 6.5km long and have an area of 120km2. The bathymetry of Bahía Santiago indicates an average depth of about 6m; it is connected with the Juluapan lagoon. The Bahía Manzanillo has an average depth of 43m, with a maximum of 86m and, near the coastline, a minimum depth of 10m (Secretaría de Marina 1973, Galicia-Pérez 1987, Galicia-Pérez & Gaviño-Rodríguez 1996).
The monitoring program area included a routine monthly sampling carried out since 2009, which consisted in ten coastal and offshore stations; a phytoplankton bloom was detected between May 3rd to 20th of 2010 (Fig. 1). During this event, samples were taken at seven additional stations X1-X3 on May 7th and X4-X7 on May 12th.
Temperature and salinity: Both parameters were measured in situ with an YSI (model 85) probe. Additionally, data was recorded every 10min at a hydrographic buoy anchored at station two. The buoy contained a CTD with temperature and conductivity sensors, and an antenna for real-time data transmission via the Argos satellite system. Therefore, this data reflected daily changes in the variables of interest and verified the equipment optimum functioning.
Nutrients: Seawater samples for nutrient analysis (DIN [NO3-, NO2-, NH4+], PO43- and SiO2) were collected with a Niskin bottle and analyzed in a Skalar San Plus autoanalyzer as described in Grasshoff et al. (1983).
Phytoplankton counting: In the course of this routine monitoring program a bloom of G. catenatum was detected, lasting from May 7th -May 17th, 2010. During this period, sampling was increased, with samples collected every fifth day at 11 stations, and visual examination of sea-surface color changes were performed every hour between 8:00 and 14:30h. Water samples were collected using a 3m long silicone hosepipe (Stations E3 and E7) or 10m (the remaining Stations), depending on the depth.
The contents of the hosepipe were mixed in a bucket, with two 500mL subsamples transferred to plastic bottles and preserved with Lugol’s solutions. Phytoplankton taxonomic identification subsamples of the integrated water samples were examined using sedimentation chambers, and an area with a representative number of cells enumerated (20mL settled and more than 300cells enumerated) under an inverted bright-field light microscope (Motic AE31), at 200-400× magnification (Throndsen 1995). The abundance distribution data was transformed using the natural logarithm, and graphs were written. The total phytoplankton population was counted on 28 samples.
Toxin sampling and analysis: Water for toxin analysis was sampled at stations E7, X1, and X3 on May 7th 2010 and at stations X4, X5, X6, and X7 on May 12th. Samples were kept in plastic bottles on ice until filtration in the laboratory. Approximately 400mL of water was filtered, or until the filters (GF/F type) were saturated; these were kept at -20oC until analyzed. The extraction process begun with the addition of 2mL of acetic acid, followed by five min of sonication in an ice bath, and by centrifugation (3 000rpm for five min). The supernatant was filtered through a single-use syringe filter (0.45μm) and an aliquot (150μL) hydrolyzed with 1M HCl. A total of 10μL of each extract (hydrolyzed and non-hydrolyzed) was injected into a HPLC system (model HP 1100) equipped with a fluorescence detector (HP 1200) and previously described methods were followed (Hummert et al. 1997, Yu et al. 1998). Paralytic toxins contained in the extracts were separated on an ion-pair buffer gradient consisting of a solution made up of octansulfonic acid, ammonia phosphate (pH 6.9) and acetonitrile. After post-column oxidation with alkaline periodic acid, the resulting products were identified with a fluorescence detector (HP 1116) set to a wavelength of 330nm for excitation and 395nm for emission. Paralytic toxins were identified by comparing chromatograms obtained from sample extracts with those resulting from an injection of standard solutions (National Research Council Canada). Toxin content was quantified by comparing the peak areas in the chromatograms of sample extracts with those of the corresponding response factor.
Additionally, a sample of a local oyster, Chama echinata Broderip, known as red oyster, was collected on May 12th 2010 at station X5. Approximately 100g of fresh tissue were homogenized with a tissue grinder, from which 2.0g were subsequently treated with 4mL of acetic acid (1M) for toxin extraction. The sample was homogenized and then centrifuged (3000rpm for five min), with the resulting supernatant treated as the phytoplankton filter for extraction and HPLC analysis.
Results
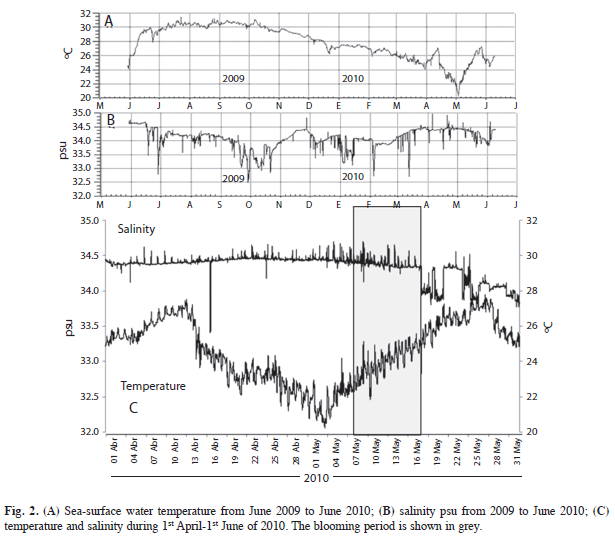
Temperature and salinity: The buoy seasurface registered temperatures from June 2009- June 2010. Results showed a rapid increase in June 2009, from to 24ºC-30ºC, this latter value persisted for the following few months. In November 2009, temperatures began to decrease gradually, reaching a minimum of 21ºC in the beginning of May 2010, when the annual cycle started again. The corresponding salinity records showed variations between 32.5 and 34.6, with an association between the low values and the area’s rainy season [average of 963.8mm (INEGI 2010)] (Fig. 2).
During the studied bloom event, salinity remained almost constant (34.4psu), but temperatures varied widely (from 20.5 to 27.5°C). Five days after a minimal temperature, a water discoloration was observed (May 7th at 22.5°C), and when the temperature reached 25.5°C on May 17th, the event was not perceived (Fig. 2).
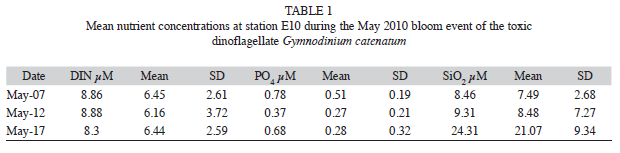
Nutrients: In general, phosphate and DIN varied only slightly, both at the surface and at 5m depth. On the contrary, silicates concentrations varied greatly in surface samples (as seen by the standard deviation); and by the end of the bloom, when a clear increase was observed at 5m depth (Table 1).
DIN values were initially low and uniform at the surface (3.1 to 7.4μM), while at 5m depth high values were consistently recorded; these values increased from the ocean towards the coast, with the maximum at E10 (12.2μM). On May 12th, DIN values were low at Bahía Santiago, with a slight increase towards Bahía Manzanillo; whereas at 5m depth, high concentrations persisted along the coast, with maximum values at E9 and E6 (about 14.4μM). By the end of the bloom, the concentration at 5m decreased (average of 8μM) between E2 and E10.
During the first two sampling days of the bloom, phosphate concentrations ranged between 0.2 and 0.8μM uniformly in both bays, but on May 17th a peak was detected toward the coast of Bahía Santiago (1.9μM), while at 5m, the low concentrations observed were similar to those recorded at the beginning of the event (0.8μM).
Silicates initially showed only a small variability, with low concentrations in both bays (about 6μM), but on May 12th both at the surface and, most notably, at 5m depth, the concentrations increased (E9 and E6), reaching a maximum of 26.1μM. At the end of the bloom, concentrations of values around 21μM were determined at surface stations located in the middle of the two bays (E10 and E5), while at 5m depth, the highest concentrations occurred at Bahía Manzanillo (16μM, E8).
Nutrient concentrations at E10 did not vary significantly for DIN (8.68μM on average), while phosphate reached a minimum on May 12th (0.37μM), that coincide with the maximum phytoplankton abundance. Silicates increased steadily in a notorious trend until May 17th, when the maximum was reached (24.31μM) (Table 1).
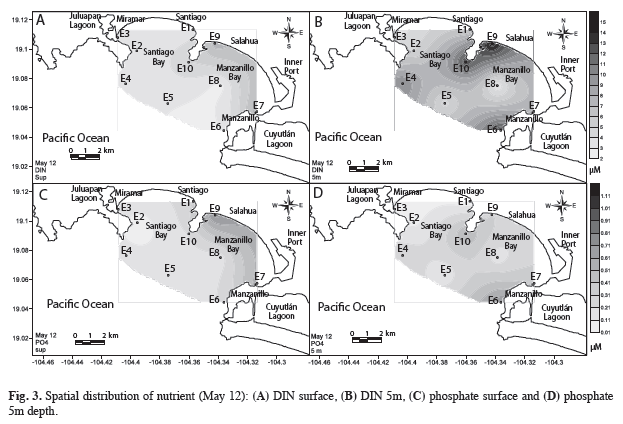
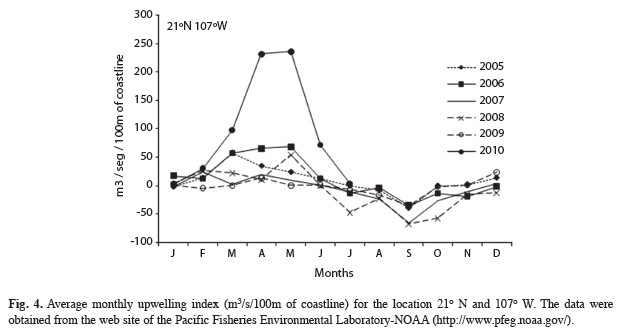
The spatial distribution of nutrients during the peak of phytoplankton abundance (May 12th) reflected the increased concentrations of DIN and silicates, both at the surface and at 5m, increasing from the ocean to the coast. Maximum values of both nutrients at E10 and E9 were maintained along the near shore, in the direction of the port, as determined based on the bay’s dynamic pattern. The distribution of phosphate showed the same distribution: with higher concentrations in surface waters towards the coast of Bahía Manzanillo (Fig. 3). It is important to note that the nutrient availability during May coincided with an upwelling period in the Central Mexican Pacific area, and was clearly the most intense event when compared to the preceding five years (Fig. 4).
Phytoplankton counting: The event was also monitored with qualitative daily observations of near shore waters. No water color changes were observed during the morning (from sunrise to midday); nevertheless, after 13 hours, water color changed throughout the afternoon, indicating high cell abundances >105cells/L.
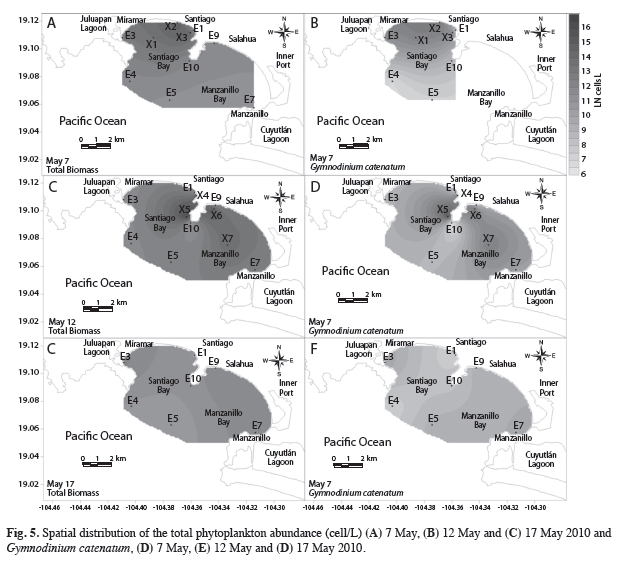
High abundances of phytoplankton at the surface were observed on May 7th in both bays, Bahía de Santiago and Bahía Manzanillo, with a maximum of 1.67×106cells/L at station X2. G. catenatum was present only in Bahía Santiago, where it reached a maximum abundance of 3.49×105cells/L for the same station (Fig. 5).
Five days later, on May 12th, high abundance of phytoplankton was observed in both bays, with a maximum of 1.02×107cells/L at X5, which coincided with a maximum of G. catenatum (3.65×106cells/L). Due to these high abundances, the water at Bahía Santiago became discolored. However, on May 17th, the bloom was not visible due to low G. catenatum densitites of 1.04×104cells/L at E5, and the maximum observed of 2.25×104cells/L at E3 (Fig. 5).
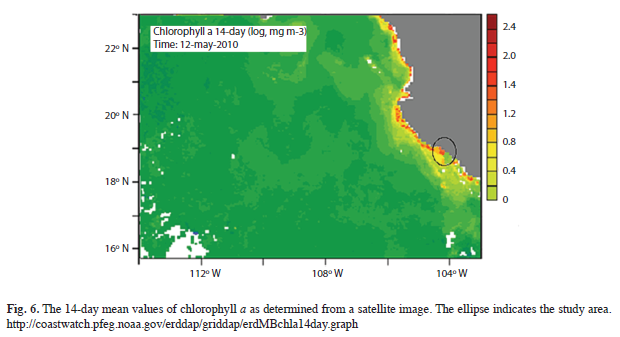
Satellite images (NOAA 2010), for the bloom period (mean composition from May 5th-17th), showed high chlorophyll a concentrations (1.2-2.0mg/m3) extending from Bahía Banderas (Jalisco, Mexico) to the Colima coast, indicating a high biomass concentration along the coastal area, including the study zone during that period (Fig. 6).
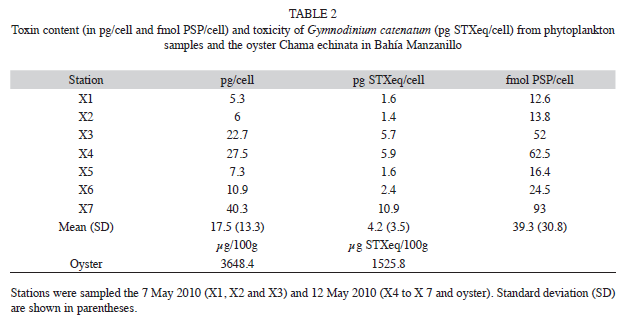
Toxin content and toxicity: Table 2 shows the toxin content and toxicity per sample station determined in a single red oyster sample. Toxin content in phytoplankton samples oscillated between 5.3 and 40.3pg/cell, with an average of 17.5pg/cell. Toxicity was also variable, ranging from 1.4 to 10.9pg STXeq/ cell, with an average of 4.2pg STXeq/cell. The high variability of both measurements is noteworthy (as evidenced by the standard deviation of 13.38 and 3.5 for toxin content and toxicity, respectively). Oyster analysis revealed a high toxin content and toxicity (3648μg/100g and 1525.8μg STXeq/100g, respectively); these values were above the saxitoxin concentration allowed for human consumption.
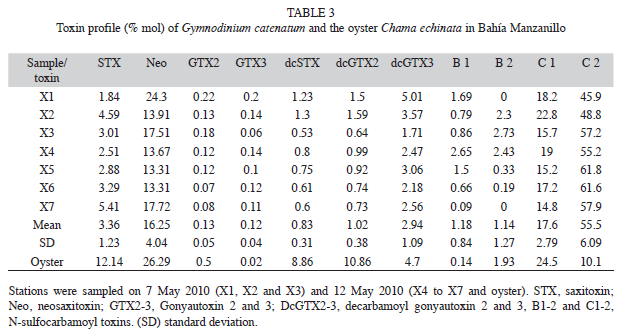
Toxin profile: Phytoplankton samples contained a group of 11 toxins (Table 3), predominantly the sulfocarbamoyl C2 (55.48%) and C1 (17.55%), followed by the more potent ones from the carbamoyl group, saxitoxin (16.2%) and neosaxitoxin (3.3%). Toxins of the decarbamoyl group (dc GTX2-3 and dcSTX) were less represented (<5%). An interesting general aspect was the small variability exhibited among the samples with respect to entire toxins % (i.e., the standard deviation). The most abundant paralytic toxins in oyster samples were those of the carbamoyl group (neosaxitoxin and saxitoxin), which represented 38% of the total toxins, followed by toxins of the sulfocarbamoyl group, with 34%.
Discussion
For this study, the lowest water temperatures occurred between March and May, which coincides with an annual pattern reported for Bahia Manzanillo, and algal blooms have been detected after these events (Morales-Blake et al. 2000, Figueroa-Torres & Zepeda-Esquivel 2001, González-Chan et al. 2008). In May, these conditions were present and allowed the proliferation of G. catenatum as reported by Band-Schmidt et al. (2004) who described that for Mexican waters, this species are able to grow in a broad array of habitats, with temperatures ranging from 15 to 29ºC and salinities from 26 to 30ups.
The origin of these algal blooms is unclear, but they could arise from offshore sources or from resting cysts present in the sediments (in shallow and low-dynamic areas). The latter possibility is, however, unlikely because near coast bay dynamics do not favor the bottom accumulation of either sandy, fine-grained sediments or, likewise, cysts (Lancin & Carranza 1976, Galicia-Pérez & Gaviño-Rodríguez 1996). According to the upwelling index from a nearby zone (at North of Bahia Banderas 21° N and 110° W), the G. catenatum bloom evaluated in this study, coincided with an upwelling period and compared to the preceding five years was clearly the most intense HAB during this interval (NOAA 2010). Torres- Orozco et al. (2005) and López-Sandoval et al. (2009) described similar conditions in the area between Cabo Corrientes and the Islas Marías. They found that the prevailing wind favored the development of coastal upwellings that generated frontal zones caused by the interaction between cold deep water and the surrounding surface water. In the present study, strong winds and cold water were likewise present, with a high content of inorganic nutrients (silicates) generated during the relaxation of a frontal zone. Therefore, the conclusions drawn by Torres-Orozco et al. (2005) may also be valid for the G. catenatum bloom of this study, but remains to be confirmed with further studies on bloom.
Daily changes in the degree of water discoloration variations could reflect the conjunction of two patterns: i) the wind influence, which in the morning, characteristically, flows from the coast to the ocean, carrying the bloom offshore, and in the early afternoon, that flows in the opposite direction, resulting in the near-shore bloom accumulation (Galicia-Pérez & Gaviño-Rodríguez 2001); and ii) the ability of G. catenatum to migrate (migration rate of 1.5m/h, as determined by Hallegraef & Fraga (1998) to sub-superficial waters during the night or early morning, with subsequent cell emergence in superficial waters in the afternoon (Hallegraeff & Fraga 1998). According to Smayda (2002) and Smayda & Reynolds (2001), during upwelling relaxation events (species type V) dinoflagellates such as G. catentum are adapted for survival within upwelling habitats and bloom during upwelling relaxations.
Our results suggest that G. catenatum is always present in the oceanic water column in low abundance. In favorable circumstances, such as upwelling relaxation, the dinoflagellate grows exponentially, forming blooms that can be transported to coastal areas. In Bahía Manzanillo, the accumulation of cells in different areas account for the typical pattern of water discoloration.
Diverse data on toxin content and toxicity for the many strains of G. catenatum found in Mexican waters have been published (Band-Schmidt et al. 2005, Gárate-Lizárraga et al. 2005, Band-Schmidt et al. 2006); however, most were obtained in studies performed under culture controlled conditions. To our knowledge, only one study has determined toxin profiles and toxicity under natural conditions in Mexico, reporting a record 1.01pg STXeq/ cell in Bahía Mazatlan during a bloom in 2001 (Gárate-Lizárraga et al. 2004a). We found even higher cell toxicity, oscillating between 1.4 and 10.9 STXeq/cell (Table 2), albeit this broad variation was most likely an abnormality, perhaps the result of physiological alterations of the cell growth phase. Factors that induce such alterations may be related to biomass production, which is influenced by the availability of inorganic nutrients. In this study, the concentrations and ratios of N and P were not limiting at the beginning of the bloom, and there was an increased availability of DIN. In addition, the oceanographic conditions, with low temperatures related to the presence of the frontal zone of an upwelling system, and the particular features of the studied geographic region, may also have influenced cell growth and thus the toxin content variability (Reguera & Oshima 1990, Flynn et al. 1996, Lippenmeier et al. 2003, Hernández-Sandoval 2010).
Cultured strains isolated from diverse sites on the Mexican coast differ in their toxicity (Band-Schmidt et al. 2006, Gárate-Lizárraga et al. 2006). In a previous paper, Band-Schmidt et al. (2010) pointed out that the toxicity of cultured strains of G. catenatum, isolated from mexican waters, were higher than those determined in field samples of the same strains. Our data supports this observation, as we found toxicity to be low under natural conditions and most likely a feature of the “Mexican” G. catenatum population, as reported by Gárate-Lizárraga et al. (2004a). As part of our study, we have isolated a strain from Bahía Manzanillo during the bloom, and we are currently measuring its toxicity under culture conditions.
The toxin profile of our phytoplankton samples consisted of 11 toxins corresponding to the profile recorded for several strains isolated from Mexican coasts (Band-Schmidt et al. 2005, Gárate-Lizárraga et al. 2005, Band-Schmidt et al. 2006). Our results confirm the dominance of toxins C1 and C2, followed by the more toxic carbamoyl group toxins. These profiles provide strong evidence for the existence of autochthonous G. catenatum strains in this region of the tropical Pacific Ocean.
The toxins decarbamoyl (dcGTX2 and dcGTX3) and N-sulfocarbamoyl (C1 and C2) usually has a high molar contribution to the toxicity of G. catenatum (Gárate-Lizárraga et al. 2004a, Hernández-Sandoval et al. 2009). In this work, a relatively high percentage of NeoSTX was noted, accounting for an average of 16.25% of the total toxins. (Bustillos-Guzmán et al. pers. omm.), in a strain of G. catenatum isolated from Bahía Concepción (Gulf of California), found that the molar percentage of each toxin changes with the growth phase of the culture.
Toxins of the sulfocarbamoyl group, mainly C1 and C2, predominate during adaptation and maximal growth, and thereafter toxins of the carbamoyl group dominate. Therefore it is plausible to infer that in the sampled bloom, G. catenatum population was in decay. This possibility is supported by the absence or low densities of dinoflagellates observed a week afterwards. It is important to note that natural toxicity and toxin composition is the result of the cell response to specific physical and chemical conditions from the sampled area (Cembella 1998, Montoya et al. 2010). Therefore in a direct comparison with other studies it is only valuable to look for a general pattern of local or regional populations, if any. Studies made in natural conditions of Mexican waters of G. catenatum present important variations in the toxin profile with no clear patterns (Gárate-Lizárraga et al. 2004a, 2004b, 2006, Hernández-Sandoval et al. 2009) indicating a clear necessity for further field investigation for this species.
Finally, despite concentrations found in oyster exceeded the Mexican regulation for saxitoxin (NOM-129-SSA-1995) of 80μg STXeq/100g allowed for human consumption there was no human poisoning during this event. Also, the presence of Neo should be considered with caution because the post-column oxidation used by Yu et al. (1998) is not very efficient in separating dcNeo from Neo (data not shown). Recently, a Mexican strain from the Gulf of California was negative for the presence of Neo (Bustillos et al., 2011).
To our knowledge, this is the first report of a G. catenatum bloom in Mexican waters where detailed physical and chemical data were obtained. Toxin analysis showed the typical toxin profile and low toxicity of natural populations, as described by Gárate-Lizárraga et al. (2004a). This low toxicity contrasts with the relatively high level recorded for cultured strains (average 23pg/cell). Oyster analysis revealed a toxin content and toxicity above the saxitoxin concentration for human consumption, with no human poisoning during this event.
Acknowledgments
This study was supported by the PROMEP project, Universidad de Colima, México, and 2009-2010 EMPAN (Establecimiento de una estación marina de mediciones físicas en relación a las proliferaciones algales nocivas en el Pacífico Mexicano. A/016933/08 AECID). We thank Fátima Castro-Ochoa, María Rivera Vilarelle, Karen Zepeda-Borja & Alejandro Reyes Herrera for their onsite effort and invaluable laboratory work. We also thank two anonymous reviewers for constructive comments on the manuscript. C.J.B.S. is a COFFA and EDI fellow of the Instituto Politécnico Nacional.
References
Band-Schmidt, C.J., L. Morquecho, C.H. Lechuga-Devéze & D.M. Anderson. 2004. Effects of growth medium, temperature, salinity and seawater source on the growth of Gymnodinium catenatum (Dinophyceae) from Bahía Concepción, Gulf of California, Mexico. J. Plankton Res. 26: 1459-1470. [ Links ]
Band-Schmidt, C.J., J. Bustillos-Guzmán, I. Gárate-Lizárraga, C.H. Lechuga-Devéze, K. Reinhardt & B. Luckas. 2005. Paralytic shellfish toxin profile in strains of the dinoflagellate Gymnodinium catenatum Graham and the scallop Argopecten ventricosus G.B. Sowerby II from Bahía Concepción, Gulf of California, Mexico. Harmful Algae 4: 21-31. [ Links ]
Band-Schmidt, C., J. Bustillos-Guzmán, L. Morquecho, I. Gárate-Lizárraga, R. Alonso-Rodríguez, A. Reyes- Salinas, K. Erler & B. Luckas. 2006. Variations of PSP toxin profiles during different growth phases in Gymnodinium catenatum (Dinophyceae) strains isolated from three locations in the Gulf of California, Mexico. J. Phycol. 42: 757-768. [ Links ]
Band-Schmidt, C.J., J. Bustillos-Guzmán, D. López-Córtes, I. Gárate-Lizárraga, E.J. Núnez-Vázquez & F.E. Hernández-Sandoval. 2010. Ecological and physiological studies of Gymnodinium catenatum in the Mexican Pacific: A Review Mar. Drugs 8: 1935-1961. [ Links ]
Bustillos-Guzmán, J., P. Vale & C. Band-Schmidt. 2011. Presence of benzoate type toxins in Gymnodinium catenatum Graham isolated from the Mexican Pacific. Toxicon 57: 922-926. [ Links ]
Cembella, A.D. 1998. Ecophysiology and metabolism of paralytic shellfish toxins in marine microalgae, pp. 381-403. In D.M. Anderson, A.D. Cembella & G.M. Hallegraeff (eds.). Physiological ecology of harmful algal blooms. NATO ASI Series, vol. G41. Springer- Verlag, Berlin/Heidelberg, Germany. [ Links ]
Figueroa-Torres, M.G. & M.A. Zepeda-Esquivel. 2001. Mareas rojas del Puerto Interior, Colima, México. Scientiae Naturae 3: 39-52. [ Links ]
Flynn, J.K., E.H. Flynn, B. John, B. Reguera, M.I. Reyero & J.M. Franco. 1996. Changes in toxins, intracellular and dissolved free amino acids of the toxic dinoflagellates Gymnodinium catenatum in response to changes in inorganic nutrients and salinity. J. Plankton Res. 18: 2093-2111. [ Links ]
Galicia-Pérez, M.A. 1987. Modelación hidrodinámica numérica de las bahías de Manzanillo y Santiago, Tesis de Mastría, UNAM, México. [ Links ]
Galicia-Pérez, M.A. & J.H. Gaviño-Rodríguez. 1996. Estudio numérico de refracción de oleaje y circulación de la bahía de Manzanillo. GEOS 16: 256. [ Links ]
Galicia-Pérez, M.A. & J.H. Gaviño-Rodríguez. 2001. Análisis espectral de algunos parámetros meteorológicos en Manzanillo, Colima. GEOS 21: 155-160. [ Links ]
Gárate-Lizárraga, I., J. Bustillos-Guzmán, K. Erler, M.S. Muñeton-Gómez, B. Luckas & A. Tripp-Quezada. 2004a. Paralytic shellfish toxins in the chocolate clam, Megapitaria squalida (Bivalvia: Veneridae), in Bahía de la Paz, Gulf of California. Rev. Biol. Trop. 52: 133-140. [ Links ]
Gárate-Lizárraga, I., J. Bustillos-Guzmán, R. Alonso- Rodríguez & B. Luckas. 2004b. Comparative paralytic shellfish toxin profiles in two marine bivalves during outbreaks of Gymnodinium catenatum (Dinophyceae) in the Gulf of California. Mar. Poll. Bull. 48: 378-402. [ Links ]
Gárate-Lizárraga, I., J.J. Bustillos-Guzmán, L. Morquecho- Escamilla, C.J. Band-Schmidt, R. Alonso-Rodríguez, K. Erler, B. Luckas, A. Reyes-Salinas & D.T. Góngora-González. 2005. Comparative paralytic shellfish toxin profiles in the strains of Gymnodinium catenatum Graham from the Gulf of California, Mexico. Mar. Poll. Bull. 50: 211-217. [ Links ]
Gárate-Lizárraga, I., J.J. Bustillos-Guzmán, D.J. López- Cortes, F. Hernández-Sandoval, K. Erler & B. Luckas. 2006. Paralytic shellfish toxin profiles in net phytoplankton samples from Bahía Concepción, Gulf of California, Mexico. Mar. Pollut. Bull. 52: 800-806. [ Links ]
González-Chan, R., L. Hernández-Silva, Navarro-Ornelas J.G. & M.Á. Blanco-Padilla. 2008. Proliferación algal nociva en las bahías del puerto de Manzanillo, Colima (marzo-mayo 2007). In Recursos y medio ambiente: Memorias del XIV Congreso Nacional de Ciencia y Tecnología Secretaría Educación Pública, 29-31 octubre 2007 Nuevo Vallarta, México. (compact disk). [ Links ]
Graham, H.W. 1943. Gymnodinium catenatum, a new dinoflagellate from the Gulf of California. Trans. Am. Microsc. Soc. 62: 259-261. [ Links ]
Grasshoff, K., M. Ehrrardt & K. Kremling. 1983. Methods of sea water analysis. Chemie, Weinheim, Germany. [ Links ]
Hallegraeff, G.M. 1993. A review of harmful algal blooms and their apparent global increase. Phycologia 32: 79-99. [ Links ]
Hallegraeff, G.M. & S. Fraga. 1998. Bloom dynamics of the toxic dinoflagellate Gymnodinium catenatum, with emphasis on Tasmanian and Spanish coastal waters, pp. 59-80. In D.M. Anderson, A.D. Cembella & G.M. Hallegraeff (eds.). Physiological ecology of harmful algal blooms. Springer-Verlag, Berlin, Germany. [ Links ]
Hernández-Sandoval, F.E., D.J. López-Cortés, C.J. Band- Schmidt, I. Gárate-Lizárraga, E.J. Nuñez-Vázquez & J. Bustillos-Guzmán. 2009. Paralytic toxins in bivalve mollusks during a proliferation of Gymodinium catenatum Graham in Bahía de La Paz, Mexico. Hidrobiológica 19: 245-256. [ Links ]
Hernández-Sandoval, F.E. 2010. Crecimiento y toxicidad de Gymnodinium catenatum de la bahía de la Paz y bahía de Mazatlán, Tesis de Doctorado, IPN, Baja California, Mexico. [ Links ]
Hummert, C., M. Ritscher, K. Reinhardt & B. Luckas. 1997. Analysis of the characteristic PSP profiles of Pyrodinium bahamense and several strains of Alexandrium by HPLC based on ion-pair chromatographic separation, post-column oxidation, and fluorescence detection. Chromatographia 45: 312-316. [ Links ]
INEGI. 2010. Precipitación mensual total del estado de Colima, observatorio Manzanillo. Instituto Nacional de Estadística y Geografía. (Downloaded: July 12, 2010, http://mapserver.inegi.gob.mx/geografia/espanolestados/col/precip_men_tot.cfm?c=1214&e=06&CFID=722292&CFTOKEN=21086864). [ Links ]
Kudela, R.M., J.Q. Lane & W.P. Cochlan. 2008. The potential role of anthropogenically derived nitrogen in the growth of harmful algae in California, USA. Harmful Algae 8: 103-110. [ Links ]
Kudela, R.M., S. Seeyave & W.P. Cochlan. 2010. The role of nutrients in regulation and promotion of harmful algal blooms in upwelling systems. Prog. Oceanogr. 85: 122-135. [ Links ]
Lancin, M. & E.A. Carranza. 1976. Estudio geomorfológico de la bahía y de la playa de Santiago en Manzanillo, Colima. Rev. Mex. Cienc. Geol. 2: 43-66. [ Links ]
Lippenmeier, S., D.M. Frampton, S.I. Blackburn, S. Geier & A.P. Negri. 2003. Influence of phosphorus limitation on toxicity and photosynthesis of Alexandrium minutum (Dinophyceae) monitored by in-line detection of variable chlorophyll fluorescence. J. Phycol. 38: 320-331. [ Links ]
López-Sandoval, D.C., J.R. Lara-Lara, M.F. Lavin, S. Álvarez-Borrego & G. Gaxiola-Castro. 2009. Productividad primaria en el Pacífico oriental tropical adyacente a Cabo Corrientes, México. Cien. Mar. 35: 169-182. [ Links ]
Montoya, N.G., V.K. Fulco, M.O. Carignan & J.I. Carreto. 2010. Toxin variability in cultured and natural populations of Alexandrium tamarense from southern South America- Evidences of diversity and environmental regulation. Toxicon 56: 1408-1418. [ Links ]
Morales-Blake, A., D.U. Hernández-Becerril & C. Cavazos-Guerra. 2000. Registros de mareas rojas en las bahías de Manzanillo, Colima, México pp. 155. In E. Ríos-Jara, E. Juárez-Carrillo, M. Pérez- Peña, E. López-Linares, E.G. Robles-Jarero, D.U. Hernández-Becerril & M. Silva-Briano (ed.). Estudios sobre plancton en México y el Caribe. Sociedad Mexicana de Planctología y Universidad de Guadalajara, Guadalajara, México. [ Links ]
NOAA. 2010. Pacific fisheries environmental laboratory. US Department of Commerce. (Downloaded: 12 July, 2010, http://www.pfeg.noaa.gov/). [ Links ]
NOM. 1995. Norma Oficial Mexicana NOM-129-SSA-1995. Bienes y servicios. Productos de la pesca: secos-salados, ahumados, moluscos cefalópodos y gasterópodos frescos, refrigerados y congelados. Especificaciones Sanitarias. Diario Oficial de la Federación. 6 de Marzo de 1995. México. [ Links ]
Ortiz-Lira, H. & M.C. Jiménez-Quiroz. 2006. Registro de eventos de Marea Roja en Manzanillo, Colima (1986- 2004), pp. 324-334. In M.C. Jiménez-Quiroz & E. Espino-Barr (ed.). Los recursos pesqueros y acuícolas de Jalisco, Colima y Michoacán. SAGARPA, Secretaría de Pesca, Guadalajara, Mexico. [ Links ]
Reguera, B. & M. Oshima. 1990. Response of Gymnodinium catenatum to increasing levels of nitrate: growth patterns and toxicity, pp 316-319. In E. Granéli, B. undstrøm, L. Edler & D. Anderson (ed.). Toxic marine phytoplankton. Elsevier, New York, USA. [ Links ]
Secretaría de Marina. 1973. Estudio geográfico de la región de Manzanillo, Colima. Secretaría de Marina, México, D.F., México. [ Links ]
Shipe, R.F., A. Leinweber & N. Gruber. 2008. Abiotic controls of potentially harmful algal blooms in Santa Monica Bay, California. Cont. Shelf. Res. 28: 2584-2593. [ Links ]
Smayda, T.J. 2002. Adaptations and selection of harmful and other dinoflagellate species in upwelling systems. 2. Motility and migratory behaviour. Prog. Oceanogr. 85: 71-91. [ Links ]
Smayda, T.J. & C.S. Reynolds. 2001. Community assembly in marine phytoplankton: application of recent models to harmful dinoflagellate blooms. J. Plankton. Res. 23: 447-461. [ Links ]
Throndsen, J. 1995. Estimating cell numbers, pp. 63-80. In G.M. Hallegraeff, D.M. Anderson & A.D. Cembella (eds.). Manual on harmful marine microalgae. IOC Manual and Guides UNESCO, Paris, France. [ Links ]
Torres-Orozco, E., A. Trasviña, A. Muhlia-Melo & S. Ortega- García. 2005. Mesoescale dynamics and yellowfin tuna catches in the Mexican Pacific. Cien. Mar. 31: 671-683. [ Links ]
Trainer, V.L., G.C. Pitcher, B. Reguera & T.J. Smayda. 2010. The distribution and impacts of harmful algal bloom species in eastern boundary upwelling systems. Prog. Oceanogr. 85: 33-52. [ Links ]
Trainer, V.L., M.L. Wells, W.P. Cochlan, C.G. Trick, B.D. Bill, K.A. Baugh, B.F. Beall, J. Herndon & N. Lundholm. 2009. An ecological study of a massive bloom of toxigenic Pseudo-nitzschia cuspidata off the Washington State coast. Limnol. Oceanogr. 54: 1461-1474. [ Links ]
Yu, R.C., C. Hummert, B. Luckas, P.Y. Qian, J. Li & J. Zhou. 1998. A modified HPLC method for analysis of PSP toxins in algae and shellfish from China. Chromatographia 48: 671-676. [ Links ]
*Correspondencia a: Sonia Quijano-Scheggia, Aramis Olivos-Ortiz, Juan H. Gaviño-Rodríguez, Marco A. Galicia-Pérez & Manuel Patiño-Barragan: Centro Universitario de Investigaciones Oceanológicas. Universidad de Colima. Carretera Manzanillo-Barra de Navidad km 20. Col. El Naranjo. C.P.28860. Manzanillo, Colima, México; quijanosonia@gmail.com, aolivos@ucol.mx, gavinho@ucol.mx, galicia@ucol.mx, mpkile@ucol.mx
José J. Bustillos-Guzmán, Francisco J. Hernández-Sandoval & David J. López-Cortés: Centro de Investigaciones Biológicas del Noroeste, Apdo. Postal 128, La Paz B.C.S. 23000, México; jose04@cibnor.mx, dlopez04@cibnor.mx, fhernan04@cibnor.mx
Esther Garcés: Instituto de Ciencias del Mar, CSIC. Paseo marítimo de la Barceloneta 37-49. E-08003. Barcelona, España; esther@icm.csic.es
Christine J. Band-Schmidt: Departamento de Plancton y Ecología Marina, Centro Interdisciplinario de Ciencias Marinas-Instituto Politécnico Nacional, Apdo. Postal 592, La Paz, B.C.S. 23000, México; cjband@yahoo.com
1*. Centro Universitario de Investigaciones Oceanológicas. Universidad de Colima. Carretera Manzanillo-Barra de Navidad km 20. Col. El Naranjo. C.P.28860. Manzanillo, Colima, México; quijanosonia@gmail.com, aolivos@ucol.mx, gavinho@ucol.mx, galicia@ucol.mx, mpkile@ucol.mx
2. Centro de Investigaciones Biológicas del Noroeste, Apdo. Postal 128, La Paz B.C.S. 23000, México; jose04@cibnor.mx, dlopez04@cibnor.mx, fhernan04@cibnor.mx
3. Instituto de Ciencias del Mar, CSIC. Paseo marítimo de la Barceloneta 37-49. E-08003. Barcelona, España; esther@icm.csic.es
4. Departamento de Plancton y Ecología Marina, Centro Interdisciplinario de Ciencias Marinas-Instituto Politécnico Nacional, Apdo. Postal 592, La Paz, B.C.S. 23000, México; cjband@yahoo.com
Received 11-II-2011. Corrected 30-V-2011. Accepted 29-VI-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}