










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Mechanisms of Oryza sativa (Poaceae) resistance to Tagosodes orizicolus (Homoptera: Delphacidae) under greenhouse condition in Venezuela
Alex González1*, Natalia Labrín1, Rosa M. Álvarez2*, Yorman Jayaro1, Carlos Gamboa1†, Edicta Reyes2 & Venancio Barrientos1
*Dirección para correspondencia
Abstract
Tagosodes orizicolus is one of the main plagues of rice in tropical America causing two types of damages, the direct one, feeding and oviposition effect, and an indirect one, by the transmission of the “Rice hoja blanca virus”. During 2006-2007 we carried out research under greenhouse conditions at Fundación Danac, Venezuela, in order to determine the mechanisms of antixenosis, antibiosis and tolerance to T. orizicolus, which could be acting in commercial varieties and advanced lines of the rice genetic breeding programs of INIA and Fundación Danac. The method of free feeding was used for the antixenosis evaluation, whereas the method of forced feeding was used for antibiosis evaluation (effect on survival and oviposition). Additionally, we used the indirect method based on biomass depression to estimate the tolerance. Some of the evaluated traits included: grade of damage, number of insects settling on rice plants, percentage of sogata mortality at the mature state, number of eggs in the leaf midrib and an index of tolerance. The results showed that rice genotypes possess different combinations of resistance mechanisms, as well as different grades of reactions. The susceptible control ‘Bluebonnet 50’ was consistently susceptible across experiments and the resistant control ‘Makalioka’ had high antixenosis and high antibiosis based on survival and oviposition. The rest of the genotypes presented lower or higher degrees of antixenosis and antibiosis for survival and viposition. The genotype ‘FD0241-M-17-6-1-1-1-1’ was identified with possible tolerance to the direct damage of sogata.
Key words: rice, sogata, “Rice hoja blanca virus” (RHBV), antixenosis, antibiosis, tolerance.
Resumen
Tagosodes orizicolus es una de las principales plagas del cultivo del arroz en América tropical causando dos tipos de daños a la planta, el directo, por efecto de la alimentación y oviposición; el indirecto, por la transmisión del virus de la hoja blanca del arroz. Durante el período 2006-2007 se llevó a cabo una investigación bajo condiciones de umbráculo en la Fundación Danac, Venezuela, con el objeto de determinar los mecanismos de resistencia de antixenosis, antibiosis y tolerancia a T. orizicolus que pudieran estar actuando en líneas avanzadas y cultivares del programa de mejoramiento enético de arroz de las instituciones INIA y Fundación Danac. Para la evaluación de antixenosis se utilizó el método de libre elección, para antibiosis (efecto sobre sobrevivencia y oviposición de los insectos) el de alimentación forzada, adicionalmente se usó un método indirecto para estimar la tolerancia. Dentro de los principales parámetros determinados se encontraban: grado de daño, número de insectos posados sobre plantas de arroz, porcentaje de mortalidad de sogata en estado adulto, número de huevos y se calculó un índice de tolerancia con base en la biomasa. Los resultados mostraron que los genotipos de arroz evaluados poseen diferentes combinaciones en cuanto a los mecanismos de resistencia, así como el grado de su efecto. El testigo ‘Bluebonnet 50’ se mantuvo consistentemente susceptible en las evaluaciones y el testigo resistente ‘Makalioka’ fue el único genotipo que presentó alta antixenosis y alta antibiosis para sobrevivencia y oviposición. El resto de los genotipos presentaron un menor o mayor grado de resistencia de tipo antixenosis y antibiosis para sobrevivencia y oviposición. Se identificó el genotipo ‘FD0241-M-17-6-1-1-1-1’ con posible tolerancia al daño directo de sogata.
Palabras clave: arroz, sogata, virus de la hoja blanca del arroz, antixenosis, antibiosis, tolerancia.
Rice in tropical America is affected by the planthopper Tagosodes orizicolus (Muir) (Common name: sogata) causing direct damage when the nymphs and adults feed from the mesophyll and phloem sap at different stages of plant development; or when the adults oviposit on the plant leaves (Zeigler et al. 1994). An indirect damage can also be caused by sogata as vector of the Rice hoja blanca virus (RHBV), which is one the most important disease caused by this pest (Vivas et al. 2002), reducing over 40% of rice production in Venezuela since its appearance (Malaguti 1956).
The males of T. orizicolus (sogata) are about 2mm long, dark brown or black, and the females are between 3.33 and 3.35mm long, amber lighter than males, the back of the chest to the keel is paler and this color extends to the head. The eggs are transparent and are between 0.50 and 0.70mm long. The nymphs are whitish in color and their size varies between 0.65-0.60mm long x 0.20-0.30mm wide. As they grow, the sharpness of the lateral lines of their body increases (Meneses et al. 2001). The females can lay up to 350 eggs in the spongy parenchyma into the leaf midrib. The hatching can take between four and eight days, but it can vary depending on temperature. Nymphs go through five stages whose duration can vary between 15 and 20 days. Finally, adults have a life span ranging from 14 to 36 days (Pantoja et al. 1997).
The symptoms of direct damage are accelerated leaf senescence and leaf necrosis, which begin from the apex and edges to the basal part of the leaf reducing growth and eventually causing the death of the plants (Álvarez et al. 2000). Resistance to direct damage caused by sogata is controlled by the combined action of a major gene with some modifier genes (Pardey et al. 2000, Romero 2005).
Painter (1951) identified three mechanisms of plant resistance to insects: antibiosis, tolerance and non-preference. Subsequently, other researchers recognized the behavior of insects associated with non-preference, the category called antixenosis has been widely accepted (Panda & Khush 1995). Each one of these three main categories of plant resistance may act by different mechanisms.
The genetic characteristics of the plant can be modified by conventional breeding or by the use of biotechnology techniques, which allow the flow of genes between selected plants (Badii & Garza 2007). Rice breeding programs have focused into obtaining new cultivars with increased resistance to direct and indirect damage produced by the insect. However, few studies has been made on the mechanisms of resistance in rice (Pardey et al. 1996b, Álvarez et al. 2000, Pardey et al. 2000, Calvert et al. 2004, Calvert et al. 2005). Many cultivars used as source of resistance to sogata in breeding programs have not been characterized for their resistance type. On the other hand, it have been used resistance sources that present the antibiosis mechanism, which is undesirable because sogata is able to overcome resistant in few generations (Pantoja et al. 1993, Pantoja et al. 1997).
Improvement plants resistance to RHBV and its vector is considered of paramount importance in rice breeding programs in Venezuela, because sogata populations increase considerably during the rice planting season (November-April). In the central plains area the natural populations of sogata have been characterized as high since the mid 50’s of past century, capturing around 1 700 000 insects per light trap during the dry season (Fusagri 1995, Vivas et al. 2001). On the other hand, the resurgence of populations and consequent losses caused by direct damage have been due to the improper use of pesticides (Vivas et al. 2007, Vivas et al. 2009). It is not known whether there are insect biotypes in Venezuela, as it is the case in other countries (Hernández et al. 2004).
In Venezuela, the breeding and release of new rice varieties have been conducted since 1953 by the government agency INIA and by the non-governmental sector since the middle of the decade of 1990. Fundación Danac is one of these non-governmental institutions that have constantly released new technological products, contributed to the maintenance of the Venezuelan rice circuit, and making more efficient and competitive the rice production (Fundación Danac 2010).In spite of all the efforts to obtain rice varieties with increased resistance breeding to sogata, most of the commercial varieties in Venezuela present tolerance to direct damage caused by the insect, but they do not have a suitable level of resistance to the RHBV (Vivas 1997). In the last years, only two varieties (Fundarroz PN1 and Venezuela 21) have been reported in Venezuela with high level of resistance to the virus (Labrín et al. 2009). The first one is not cultivated commercially due to instability grain quality.
In order to evaluate the genetic constitution of commercial varieties and elite lines of the breeding programs of INIA and Danac, several experiments were carried out under greenhouse conditions, with the aim of characterizing the different resistance mechanisms to T. orizicolus that could be acting. Additionally, the tolerance level of cultivars and lines was evaluated.
Materials and Methods
Study site: Five experiments were established from August 2006-September 2007 in a greenhouse of Foundation Danac, located at 10°21’50” N - 68°59’15” W in San Javier Yaracuy State, Venezuela. The average temperature and humidity of the greenhouse was 29.5ºC and 68.7% HR, respectively.
Rice genotypes: Three current commercial cultivars were evaluated: ‘Venezuela 21’, ‘Centauro’ and ‘D-Sativa’, and four promissory advanced lines ‘PN00A016C’, ‘PFD-2\1\1\2> 293-M-6-2-2-1’, ‘FD0241-M-17-6-1-1-1-1’ and ‘FD0241-M-18-3-1-2-M-1’ the genetic breeding programs of INIA and Fundacion Danac. The controls used were ‘Makalioka’ and ‘Fedearroz 50’ as resistant, ‘Araure 4’ as intermediate, and ‘Bluebonnet 50’ as susceptible to sogata.
Insect populations: Insects were collected from an insect population at Los Bancos de San Pedro in the Sistema de Riego Rio Guárico (SRRG), Guárico, Venezuela, located at 8°83’53” N - 67°53’40” W, where a strong incidence of insects has been reported (Fusagri 1995).
Laboratory rearing: The insects were kept in a cage inside the greenhouse for their reproduction. Sequential colonies of T. orizicolus, free of the RHBV, were used (Triana et al. 2003).
Antixenosis using free-choice test: a) ircle (adapted from Heinrichs et al. 1985): Ten seeds of each genotype and control were planted in pots of 7.50cm high x 9.00cm in diameter. A complete randomized block design with four replications was used. Plants were thinned to five per pot and fertilized with urea eight days after planting. Plants of 25 days were placed into four entomological cages (80cm x 100cm x 65cm) in a circular arrangement. A dose of five insects per plant, of fourth and fifth instar, was used and placed in the center of the circle in each cage. After 24, 48 and 72h of infestation, the number of settled insects per plant were counted. Finally, the direct damage was evaluated 30 days later, when ‘Bluebonnet 50’ reached grade nine in the range of the Standard System Evaluation for Rice (IRRI 2002). The resistance range used was: resistant ≤3, intermediated between three and five and susceptible >5.
b) Row (adapted from Triana et al. 2003): In plastic trays, 10 seeds of each material per row at 10cm long and 3cm between rows were planted. A completely randomized design with four replications was used. Five plants were left in the row and fertilized with urea eight days after planting. 25 days after planting, trays were placed into an entomological cage (80cm height x 100cm length x 50cm deep) and plants were infested with five nymphs per plants at the fourth and fifth instar. Plants were shacked out every day until 30 days after infestation. The evaluation of direct damage was made when Bluebonnet 50 reached the highest in the range according to the Standard System Evaluation for Rice (IRRI 2002).
Antibiosis using no-choice test for survival: Ten seeds of each genotype were planted in pots of 9.50cm x 11.50cm under a completely randomized design with three replications. Plants were thinned to five per pot and fertilized with urea eight days after planting. 15 days after planting, plants were covered with micro cages (made with 2L clear plastic bottles), then the plants were infested with 10 pairs of young adult insects (21 to 25 days after hatching) per pot; the number of dead insects per plot was counted daily until all of them died. Dead insect data obtained were transformed to percentage of mortality. The analysis was made with the percentage of mortality at eight days after infestation, when the mortality of insect on the susceptible cultivar ‘Bluebonnet 50’ reached 20%; it was made to avoid the antibiosis effect from the natural mortality of insects (Álvarez et al. 2000).
Direct damage in plants using no-choice test for survival: The conditions and design method in this assay were the same used in the latter experiment. The infestation was made only with five pairs of young adults insects (21- 25 days after hatching) per plot; those plants were not shacked out. Direct damage in each plant was evaluated 30 days after infestation to determine the genotype reaction under forced feeding conditions (Álvarez et al. 2000). Phenotypic evaluation was done using the Standard System Evaluation for Rice (IRRI 2002).
Comparison of direct damage between free-choice in row and no-choice test: The mean of the direct damage of genotypes obtained in the free-choice in row test and nochoice test for survival were compared.
Antibiosis using no-choice test for oviposition: For each rice genotype, five seeds per plot were planted in pots of 9.50cm x 11.50cm under a complete randomized design with four replications. One plant per plot was left eight days after planting and fertilized with urea. 29 days after planting, plants were covered (with micro cages made with 2L clear plastic bottles), and then a pair of young adult insects was release. Ten and twenty days after infestation, replacement of genotype was made and the number of eggs was counted in each plant, dissecting the tissue with the tip of a syringe needle and using a stereoscope LiederTM MZ- 720X (China) (10X) to visualize the eggs (Álvarez et al. 2000).
Tolerance in greenhouse conditions: Two trials were made for evaluation of tolerance. One of them was conducted under a completely randomized design with two replications, and the experimental unit (EU) consisted of two 30 days old plants per pot; those plants were covered with micro cages (made from 2L clear plastic bottles) and infested with a dose of eight insects per plant. Other identical trial was performed without insect infestation. When the susceptible check ‘Bluebonnet 50’ reached nine of the Standard System Evaluation for Rice (IRRI 2002) in the infested trial, all the plants were removed from the greenhouse; infected plants were placed at -20ºC for 15 minutes, this process allowed to cut down the plants, inactivate and remove insect and evaluate the direct damage. Subsequently, insects and piece of plants with and without infestations were dried at 60ºC for 48h. Dry weight for insects (DWI) and dry weight of infested and uninfested plants was registered. Subsequently, the Functional Plant Loss Index (FPLI) was calculated using the formula proposed by Heinrichs et al. (1985).
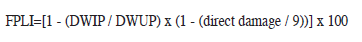
Where: DWIP: dry weight of infested plants. DWUP: dry weight of uninfected plants. Data from the six experiments were analyzed by analysis of variance (ANOVA) and mean test using InfoStat program (InfoStat 2004) with a significance level of 0.05. When necessary, the equivalent non-parametric tests were performed.
Results
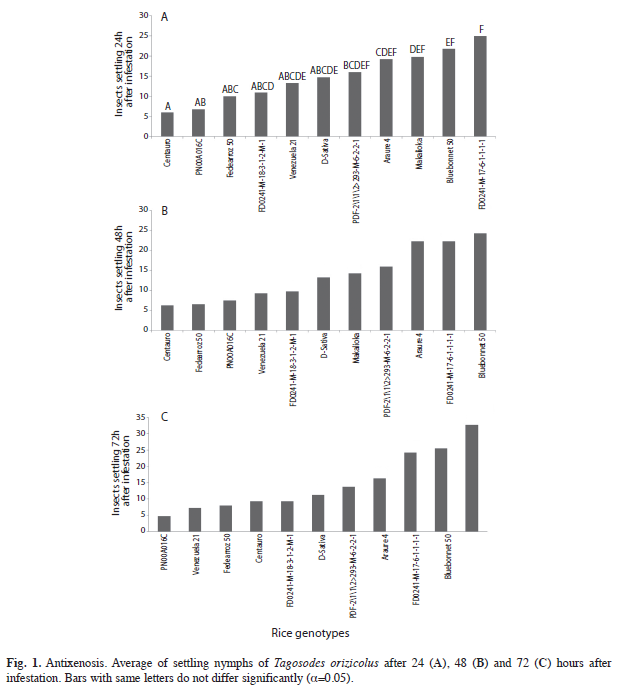
Antixenosis using free-choice test: a) Circle: The ANOVA showed significant differences in the number of settled insects between all the genotypes at 24, 48 and 72h after infestation [p(24h)=0.0006, p(48h)=0.0003, p(72h)=0.0001]. The genotypes ‘FD0241- M-17-6-1-1- 1-1’, ‘Bluebonnet 50’, ‘Makalioka’ and ‘Araure 4’ were more attractive to the insects 24h after infestation (Fig. 1). However, sogata showed less preference to ‘Makalioka’ at 48 and 72h. ‘Bluebonnet 50’ and ‘FD0241-M-17-6-1-1-1-1’ were the most preferred genotypes since the beginning until the end of the trial. The cultivars Araure 4, D-Sativa and the line ‘PFD-2\1\1\2>293- M-6-2-2-1’ showed intermediate antixenosis, while ‘FD0241-M-18-3-1-2-M-1’, ‘Centauro’, ‘Makalioka’, ‘Fedearroz 50’, ‘Venezuela 21’ and ‘PN00A016C’ showed the highest grade of antixenosis, reflected by the low preference of insects.
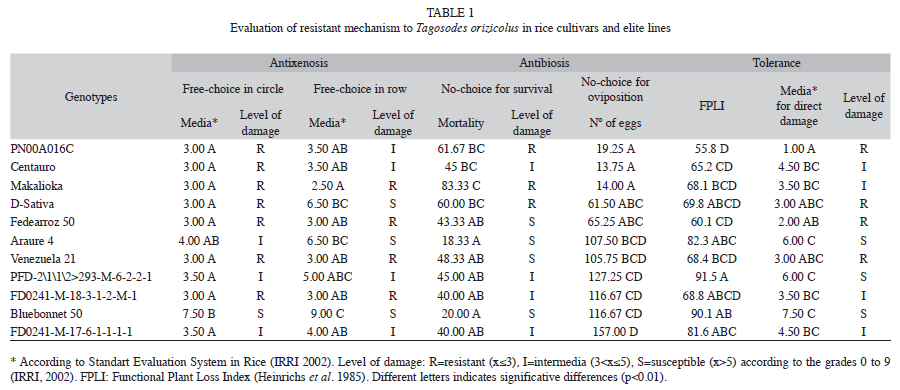
b) Row: The non-parametric analysis detected significant differences between cultivars (Kruskal-Wallis; p=0.0024), being ‘Bluebonnet 50’, ‘Araure 4’ and ‘D-sativa’ which showed the highest susceptibility level to direct damage. On the other hand, ‘PN00A016C’, ‘Centauro’, ‘PFD-2\1\1\2>293- M-6-2-2-1’ and ‘FD0241-M-17-6-1-1-1-1’ showed intermediate resistance level, while ‘Makalioka’, ‘FD0241-M-18-3-1-2-M-1’, ‘Fedearroz 50’ and ‘Venezuela 21’ exhibited the highest resistance level to direct damage caused by T. orizicolus (Table 1).
The circle and row feeding experiments showed contrasting results regarding genotype resistance and susceptibility to mechanical damage. In the circle feeding experiment, seven resistant varieties, three with intermediate reaction and one susceptible, could be differentiated; whereas in the row feeding experiment a greater number of susceptible genotypes were observed. ‘PN00A016C’ and ‘Centauro’ changed from resistant in the circle feeding experiment to susceptible in the row feeding exposure. The most drastic effect was observed on ‘D-Sativa’ that changed damage level from resistant to susceptible. A similar result was observed in ‘Araure 4’ which changed from intermediate level observed in the circle method to susceptible in row method.
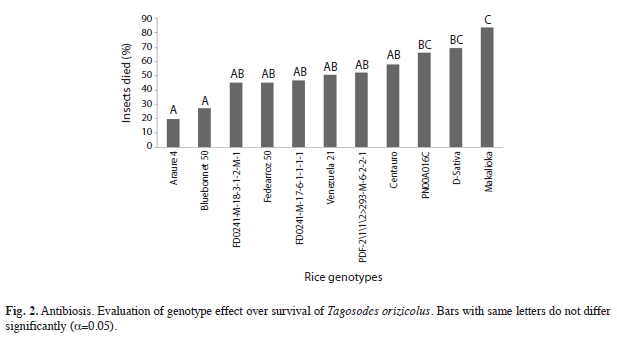
Antibiosis using no-choice test for survival: Significant differences (p=0.0044) on insects mortality were detected between rice genotypes using no-choice test for survival. The Duncan mean test showed four homogeneous groups for survival effect on sogata. ‘Makalioka’ exhibited the highest antibiosis level, where 83.33% of adult insects died, while ‘Araure 4’ and ‘Bluebonnet 50’ did not show any deleterious effects on sogata; the other genotypes showed intermediate and high level of antibiosis (Fig. 2).
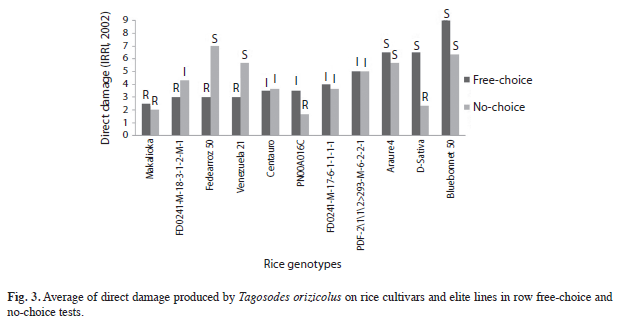
Comparison of direct damage between free-choice in row and no-choice test: There were no significant differences in direct damage (p=0.1631) between mean treatments in no-choice test. However, the mean value over all the genotypes was used to compare both feeding methods (Fig. 3). The cultivars ‘Venezuela 21’, ‘Fedearroz 50’ and ‘FD0241-M-18- 3-1-2-M-1’ showed typical reactions of high antixenosis, where the genotypes showed resistance in free-choice and were susceptible or intermediate in no-choice conditions. ‘Makalioka’, ‘Centauro’, ‘FD0241-M-17-6-1-1-1-1’ and ‘PFD-2\1\1\2>293-M-6-2-2-1’ were the cultivars that showed more stable resistance to sogata across both methods.
Antibiosis using no-choice test for oviposition: In this trial, statistical differences (p=0.0006) between cultivars were detected. Six different groups were obtained with the Duncan´s test (Table 1) where ‘Makalioka’, ‘Centauro’ and ‘PN00A016C’ exhibited higher antibiotic effect, and FD0241-M-17-6-1-1-1-1 had lowest effect on oviposition of the insects. The genotype ‘FD0241-M-17-6-1-1-1-1’ showed lower level of antibiosis for oviposition than ‘Araure 4’ and ‘Bluebonnet 50’, despite it exhibited moderate level of antibiosis for survival.
Tolerance in greenhouse conditions: The FPLI showed statistical differences between treatments (p=0.043), seeing that some genotypes lost a significant amount of biomass as a result of insect infection. Differences related to the FPLI and direct damage in each treatment was observed through mean test (Table 1). Considering the FPLI and direct damage, ‘Centauro’ was the most tolerant cultivar to T. orizicolus, because it had one of the lowest FPLI, with a considerable level of direct damage. Several genotypes showed low FPLI, but associated with a low level of direct damage. The line FD0241-M-17-6-1-1-1-1 showed a lower FPLI than susceptible ‘Bluebonnet 50’, with direct damage mean of 4.50; which could be indication of tolerance for that genotype.
Discussion
Antixenosis using free-choice test: This method, proposed by Heinrichs et al. (1985) to detect antixenosis in rice for Nilaparvata lugens Stal (Delphacidae), Nephotettix virescens Distant (Cicadellidae), Sogatella furcifera Horvath (Delphacidae) y Recilia dorsalis Motshulsky (Cicadellidae), was effective to study feeding preference of T. orizicolus under greenhouse conditions.
There are few references related to freechoice feeding using circle arrangements in rice exposed to T. orizicolus; however the number of settling insect expressed as percentage at 24, 36 and 72h were similar to those reported by Calvert et al. (2004), where ‘Fedearroz 50’ and ‘Makalioka’ were consistently less preferred by sogata.
The results from free-choice feeding in the row arrangement were similar to previous reports, where ‘Makalioka’ and ‘Bluebonnet 50’ were more resistant and susceptible, respectively (Triana et al. 1994). High susceptibility was shown by ‘Araure 4’ using this method under greenhouse conditions (Álvarez et al. 2002), but under commercial cropping conditions, where the insect density was higher (30 insect/double mesh passed), this cultivar exhibited intermediate resistance (Fusagri 1995). However, it was found that the number of T. orizicolus adults in ‘Araure 4’ was six fold higher during the dry season in the Sistema de Riego Rio Guárico (SRRG) than the rainy season (Vivas & Clavijo 2000).
No report was found comparing simultaneously free-choice feeding method, circle and row arrangements. The daily removal of insects from row method probably decreased the antixenosis mechanism in some cultivars due to longer exposure of plants to insect effects. However, row method has been used to evaluate direct damage in rice breeding programs, because it provides a better discrimination among resistant, intermediate and susceptible genotypes (Pardey et al. 1996b). Furthermore, this method has been useful to study the genetics of resistance against the direct damage produced for sogata (Pardey et al. 1996a).
Antibiosis using no-choice test for survival: These results were similar to those found by Triana et al. (1994), where ‘Makalioka’ and ‘Bluebonnet 50’ showed the highest and lowest insect mortality, respectively. Also, it was also possible to confirm that T. orizicolus lived longer on susceptible cultivars than on resistant ones; the same result was observed by Pardey et al. (1996b). Most of the evaluated cultivars showed high to intermediate antibiosis survival; actually those genotypes are being used as progenitors in the breeding programs in Venezuela, which could increase the possibility to select new biotypes of insect (Weber & Parada 1994). Antibiosis should be multigenic characteristic to be stable in rice; otherwise the insects would be able to develop new biotypes and races (Heinrichs 1994), which could be resistant to antibiosis mechanism from plants (Baddii & Garza 2007).
Comparison of direct damage between free-choice in row and no-choice test: The difference observed between free-choice and no-choice tests is that in the former insects have the option to feed and multiply on the genotypes they select, whereas in the no choice test the insects are forced to feed on each genotype (Pardey et al. 1996b). The latter is similar to what occurs in commercial planting where one
cultivar dominates, in which case, antixenosis resistance should be short lasting (Badii & Garza 2007). ‘Venezuela 21’ and ‘Fedearroz 50’ were resistant to insects under free-choice feeding; however, they were susceptible under no-choice test. Pardey et al. (1996b) reported very similar results in ‘Fedearroz 50’. On the other hand, ‘D-sativa’, which is the most planted cultivar in Venezuela (Fundación Danac 2010), exhibited resistance under no-choice feeding and susceptibility under free-choice. It is possible that under free-choice, ‘D-sativa’ had a heavier dose of infestation than under no-choice feeding.
Antibiosis using no-choice test for oviposition: These results were similar to those obtained by Pardey et al. (1994b) who determined that ‘Makalioka’ had higher antibiosis effect for oviposition, the same has been reported for ‘Fedearroz-50’ (Calvert et al. 2004). Morphological differences have been observed in cultivars which have high antibiosis, those differences are associated with the presence of pubescent leaves, which hamper the oviposition of insects or the nutritional quality for the insects (Baddi & Garza 2007), where the absence of some essential amino acid makes he insects weaker to reproduce (Heinrichs
1994, Panda & Khush 1995).
Tolerance in greenhouse conditions: None of the yield parameters could be evaluated because plants did not reach the reproductive phase due to the direct damage caused by the insects. Therefore, biomass was reduced during the first growth phases, limiting its capacity to recover and produce grain.
Tolerance to sogata in ‘Araure 4’ has been reported previously (Vivas 1997); however, in this study this cultivar showed a high FPLI and grade six for direct damage, these results are consistent with those found under field condition (Vivas et al. 2002). The cultivar Centauro showed a higher DWI, in high insect density conditions, showing grade five of direct damage. However, ‘Centauro’ showed one of the lowest FPLI values observed among all genotypes. According to the method and results of our study, we suggest that Centauro is a tolerant cultivar to direct damage caused by T. orizicolus (Heinrichs et al. 1985). Additionally, the line FD0241-M-17-6-1-1-1-1 showed indications of tolerance, although in lower level that ‘Centauro’.
The existence of high antibiosis in ‘Makalioka’ interferes with the determination of the tolerance mechanism, due to the high mortality of adult insects. Thus, despite having a FPLI similar to the observed in ‘Centauro’, the lower level of damage expressed in ‘Makalioka’, suggests the possibility that the small loss of biomass is due to the early insects death rather than the presence of the tolerance mechanism. The graphing analysis proposed by Heinrichs et al. (1985) could not be done due to the low number of insects collected at the end of this experiment, which allowed the determination of the DWI value. Moreover, the number of live insects was very low or completely absent at the nymph stage, this could be caused by the low number of females at the time of infestation, since we used insects from fourth and fifth nymphal stages, that did not allow differentiation between males and females.
Mechanism of rice genotypes resistance to T. orizicolus: From the agricultural perspective, a resistant cultivar is defined when it yields is better under the pressure of an insect plague compared with a susceptible one (Niks & Lindhout 1999). Thus, the resistance of a plant to a plague is a relative term based on the comparison with a susceptible plant (Niks & Linhout 1999, Baddi & Garza 2007). In this research a thorough comparison of the different groups was made, and it was possible to see that all cultivars expressed some kind of resistance mechanism differing in type and expression level. For example, ‘Makalioka’ showed higher levels of resistance to direct damage produced by sogata, this resistance was determined by the antixenosis mechanism which increased as the exposition time to insect was longer; besides, survival and oviposition antibiosis were high, coinciding with other reports (Pardey et al. 1996b, Calvert et al. 2004).
The cultivars Fedearroz 50 and Venezuela 21 are susceptible to direct damage, even though they have high antixenosis and medium antibiosis for survival and oviposition under greenhouse conditions.‘D-sativa’ has medium antixenosis and medium-high antibiosis for survival and oviposition; however, it becomes susceptible to direct damage at certain insect population densities.
‘Centauro’ is a moderately resistant cultivar to direct damage. The resistance is provided by strong antixenosis, high antibiosis for oviposition, medium-high antibiosis for survival and tolerance to direct damage. Based on antibiosis values, it could be a risk to plant this cultivar at a commercial scale in Guárico State during the dry season, when the natural sogata population densities are high.
‘Bluebonnet 50’ appears to be preferred by the insect for feeding and oviposition, it showed low antibiosis for oviposition and high susceptibility to direct damage caused by sogata, coinciding with previous reports (Pardey et al. 1996b, Calvert et al. 2004). ‘Araure 4’ behaved similarly to ‘Bluebonnet 50’, the difference is that ‘Araure 4’ is more tolerant to direct damage making it a good choice for feeding insect mass rearing. Under greenhouse conditions the tolerance test allowed the identification of ‘Centauro’ as possible genotype tolerant to direct damage produced by T. orizicolus, which should be corroborated with field trials.
In summary, the tested cultivars expressed a resistance mechanism to sogata differing in he type of mechanism as well as in expression level. Most commercial rice cultivars tested and elite lines have inadequate antibiotic effects on the insect and they can lead in the future the development of new insect biotypes which can be more aggressive in the production area of the central plains of Venezuela. However, the line ‘FD0241-M-17-6-1-1-1-1’ showed an intermediate behavior for all mechanisms, identifying itself as resistant progenitor to T. orizicolus to be use in the breeding programs in Venezuela.
Acknowledgments
Authors want to thank to Fondo Nacional de Ciencia Tecnología e Innovación (FONACIT) of Venezuela for partial funding given to Project Nº 2000001658, entitled “Rice Germoplasm Evaluation resistance to direct damage produce by sogata (Tagosodes orizicolus Muir) and hoja blanca virus”, whose partial results are presented in this paper.
References
Álvarez, R.M., C.E. Gamboa, M. Triana, M.C. Duque & J. Silva. 2000. Mecanismo de resistencia a Tagosodes orizicolus Muir (Homoptera: Delphacidae) de tipo antibiótico y no preferencia en algunas líneas de arroz (Oryza sativa). Investigación Agrícola 5:1 (Consultado: 28 julio 2008, http://www.redpavpolar.info.ve/danac/viewarticle.php?id=22&layout=html). [ Links ]
Baddi, M.H. & V. Garza. 2007. Resistencia en insectos, plantas y microorganismos. Universidad Autónoma de Ciudad Juárez. CULCyT. Año 4, Nº 18. Enero-febrero. (Consultado: 7 julio 2008, http://www2.uacj.mx/IIT/CULCYT/EneroFebrero2007/5ARTCULOBADII.PDF). [ Links ]
Calvert, L., C. Pardey, M. Triana & R. Meneses. 2004. Avances en la búsqueda de mecanismos de resistencia a Tagosodes orizicolus (Muir) Plaga del arroz en América latina. Simposio de manejo agroecológico de plagas en la agricultura tropical. Rev. Protección Veg. 19: 3. [ Links ]
Calvert, L., Lozano, I., N. Villarreal, L. Reyes, L. Romero, M. Lorieux, A. Garavito, M. Duque & C. Martínez. 2005. Hacia la comprensión de la base genética de la resistencia al complejo virus de la hoja blanca y Tagosodes orizicolus en arroz. XLV Reunión Anual APS, San José, Costa Rica. [ Links ]
Fundación Danac. 2010. Fundación para la Investigación Agrícola Danac. Informe Danac 2008-2009: Acciones para el sector agroalimentario. San Javier, Danac. 63p. (Consultado: 25 mayo 2010, http://www.danac.org.ve/) [ Links ]
Fusagri. 1995. La sogata: Plaga del arroz en el Sistema de Riego Rio Guárico. Fundación Servicio Para el Agricultor. Noticias Agrícolas. Año 39, Nº 14. Marzo-abril. [ Links ]
Heinrichs, E.A. 1994. Biology and management of rice insects. Wiley Eastern Limited, New Delhi, India. [ Links ]
Heinrichs, E.A., F.G. Medrano & H.R. Rapusas. 1985. Genetic evaluation for insect resistance in rice. International Rice Research Institute. Los Baños, Laguna, Philippines. [ Links ]
Hernández, M., T. Quesada, C. Muñoz & A.M. Espinoza. 2004. Genetic diversity of Costa Rican populations of the rice planthopper Tagosodes orizicolus (Homoptera: Delphacidae). Rev. Biol. Trop. 52: 795-806. [ Links ]
Infostat. 2004. Infostat. Versión 2004. Universidad Naciona de Córdoba. Córdoba, Argentina. [ Links ]
IRRI. 2002. Reference Guide - Standard Evaluation System for Rice. International Rice Research Institute. (Consultado: 2 mayo 2008, http://www.knowledgebank.irri.org/ses/). [ Links ]
Labrín, N., A. Ebert, I. Pérez-Almeida, C. Astorga, G. Rivas & L. Calvert. 2009. Evaluación de la resistencia de cultivares de arroz venezolanos ante el virus de la hoja blanca y su asociación con marcadores microsatélites. Fitopatol. Venez. 22:13-18. [ Links ]
Malaguti, G. 1956. La “hoja blanca”, extraña enfermedad del arroz en Venezuela. Agronomía Trop. 6: 141-145. [ Links ]
Martínez, C. 2005. Hacia la comprensión de la base genética de la resistencia al complejo del virus de la hoja blanca y Tagosodes orizicolus en arroz. Resúmenes de la XLV Reunión Anual de la APS. San José, Costa Rica. [ Links ]
Meneses, C.R., Y.A. Gutiérrez, R.A. García, P.G. Antigua, S.J. Gómez, F. Correa & L. Calvert. 2001. Guía para el trabajo de campo en el manejo integrado de plagas del arroz. Colombia, FLAR. [ Links ]
Painter, R.H. 1951. Insect resistance en crop plants. Mac- Millan, New York, USA. [ Links ]
Panda, N. & G.S. Khush. 1995. Host plant resistance to insects. CAB International & International Rice Research Institute. Manila, Philippines. [ Links ]
Pantoja, A., A. Fischer, F. Correa-Victoria, L. R. Sanint & A. Ramírez. 1997. MIP en Arroz: Manejo integrado de plagas; artrópodos, enfermedades y malezas. Centro Internacional de Agricultura Tropical, Cali, Colombia. [ Links ]
Pantoja, A., R. Zeigler & F. Correa. 1993. Screening rice germoplams for Tagosodes orizicolus resistance and detecting the percentage of vector, p. 53-55. In Proceeding of monitoring tour and workshop on integrate management of rice in the Caribbean. CIAT (eds.). Cali, Colombia. [ Links ]
Pardey, C., D. Baena & F. Cuevas. 1996a. Herencia de la resistencia al daño mecánico causado por Tagosodes orizicolus (Muir) [Homoptera-Delphacidae] en arroz Oryza sativa. Acta Agron. 46: 9-14. [ Links ]
Pardey, C., F. Cuevas, C. Martinez & A. Pantoja. 2000. Herencia de la resistencia al daño mecánico por Tagosodes orizicolus (Muir) (Homoptera: Delphacidae) en arroz. J. Agri. Univ. Puerto Rico: 84: 47-52. [ Links ]
Pardey, C., F. Cuevas, D. Baena & C. Martínez. 1996b. Caracterización de la resistencia al daño mecánico de Tagosodes orizicolus (Muir) (Homoptera: Delphacidae) en doce cultivares de arroz (Oryza sativa). Rev. Col. Entomol. 22: 37-43. [ Links ]
Romero, L.E. 2005. Aproximación hacia la identificación de marcadores moleculares microsatélites en arroz (Oryza sativa L.) (Gramíneae) para la resistencia a Tagosodes orizicolus M. (Homoptera-Delphacidae). Tesis Licenciatura, Universidad del Valle, Cali, Colombia. [ Links ]
Triana, M., M. Cruz, R. Meneses & L. Calvert. 2003. Metodologías para la cría y evaluación de Tagosodes orizicolus (Muir). Manual Técnico. Centro Internacional de Agricultura Tropical. Cali. Colombia. También disponible en línea: http//www.ciat.cgiar.org/riceweb/index1.htm [ Links ]
Triana, M., A. Pineda, C. Pardey, A. Pantoja & M. Duque. 1994. Mecanismos de resistencia al daño mecánico por Tagosodes orizicolus (Muir) en arroz. Rev. Col. Entomol. 20: 23-26. [ Links ]
Vivas, L.E. 1997. Dinámica poblacional de la sogata del arroz Tagosodes orizicolus (Homoptera: Delphacidae) en el Guárico occidental. Tesis de Maestría, Universidad Central de Venezuela, Maracay, Venezuela. [ Links ]
Vivas, L.E. & S. Clavijo. 2000. Fluctuación poblacional de Tagosodes orizicolus (Muir) 1926 (Homoptera: Delphacidae) en el sistema de riego Río Guárico, Calabozo, estado Guárico, Venezuela. Bol. Entomol. Venezuela 15: 217-227. [ Links ]
Vivas, L.E, S. Clavijo & H. González. 2001. Distribución temporal y espacial y numero de muestras optimo para la estimación de las poblaciones de sogata Tagosodes orizicolus, (Muir) 1926 (Homoptera : Delphacidae) en el cultivo de arroz, en Calabozo estado Guárico, Venezuela. Investigación Agrícola 6:1. (Consultado: 15 febrero 2007, http://www.redpavfpolar.info.ve/danac/volumen6/art1/index.html). [ Links ]
Vivas, L.E., L. Lugo, M. Acevedo & S. Clavijo. 2002. Determinación de la preferencia de Tagosodes orizicolus (MUIR) 1926 (Homoptera: Delphacidae) sobre variedades de arroz en Calabozo, estado Guárico, Venezuela. Investigación Agrícola. 7: 1. (Consultado: 28 febrero 2007, http://www.redpav-fpolar.info.ve/danac/viewarticle.php?id=33). [ Links ]
Vivas, L.E., D. Astudillo & L. Campo. 2007. Evaluación del insecticida Etofenprox 10,9% para el control del insecto sogata en el cultivo del arroz en Calabozo Estado Guárico, Venezuela. Agronomía Trop. 57: 287-297. [ Links ]
Vivas, L.E., D. Astudillo & L. Campo. 2009. Evaluación del insecticida Thiamethoxan 25% para el manejo del insecto sogata en el cultivo del arroz en Calabozo, Estado Guárico, Venezuela. Agronomía Trop. 59: 89-98. [ Links ]
Weber, G. & O. Parada. 1994. Development of an Integrated Pest Management System for rice in Latin America, p. 733- 748. In E.A. Heinrichs (ed.). Biology and management of rice insects. Wiley Eastern Limited, New Delhi, India. [ Links ]
Zeigler, R.S., A. Pantoja, M.C. Duque & G. Weber. 1994. Characteristics of resistance in rice to rice hoja blanca virus (VHB) and its vector. Ann. Appl. Biol. 124: 429-440. [ Links ]
*Correspondencia a: Alex González, Natalia Labrín, Yorman Jayaro, Carlos Gamboa, Venancio Barrientos. Fundación para la Investigación Agrícola Danac, Carretera San Javier vía Guarataro, Finca Naranjal, Apdo. 182. San Felipe, Venezuela; alex.gonzalez@danac.org.ve, natalia.labrin@danac.org.ve, yorman.jayaro@danac.org.ve, venancio.barrientos@danac.org.ve
Rosa M. Álvarez,Edicta Reyes. Instituto Nacional de Investigaciones Agropecuarias, INIA-Araure, Venezuela; rmalvarez@inia.gob.ve, ereyes@inia.gob.ve
1. Fundación para la Investigación Agrícola Danac, Carretera San Javier vía Guarataro, Finca Naranjal, Apdo. 182. San Felipe, Venezuela; alex.gonzalez@danac.org.ve, natalia.labrin@danac.org.ve, yorman.jayaro@danac.org.ve, venancio.barrientos@danac.org.ve
2. Instituto Nacional de Investigaciones Agropecuarias, INIA-Araure, Venezuela; rmalvarez@inia.gob.ve, ereyes@inia.gob.ve
Received 22-XI-2010. Corrected 03-VI-2011. Accepted 07-VII-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}