










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.1 San José Mar. 2012
Prosopis laevigata and Mimosa biuncifera (Leguminosae), jointly influence plant diversity and soil fertility of a Mexican semiarid ecosystem
Rosalva García-Sánchez1*,2*, Sara Lucía Camargo-Ricalde3*, Edmundo García-Moya1, Mario Luna-Cavazos1, Angélica Romero-Manzanares1 & Noé Manuel Montaño3
*Dirección para correspondencia
Abstract
Prosopis laevigata and Mimosa biuncifera are frequently found in arid and semiarid shrublands, but scarce information is available about their influence on plant community structure and soil fertility. We compared plant community structure, diversity and soil nutrients of three semiarid shrubland sites located in Mezquital Valley, Mexico. These sites differ in their dominant species: Site 1 (Bingu) P. laevigata, Site 2 (González) M. biuncifera, and Site 3 (Rincón) with the presence of both legumes. The results showed that the plant community with P. laevigata and M. biuncifera (Site 3) had more cover, taller plants and higher plant diversity than sites with only one legume (Site 1 and Site 2). Soil organic matter (SOM), soil organic carbon (SOC), total nitrogen (TN), phosphorus-Olsen (P) and C mineralization were higher in the soil under the canopy of both legumes than in bare soil. In contrast, soil cation concentrations were lower under the canopy of P. laevigata, but not for M. biuncifera. In addition, the density of arbuscular mycorrhizal fungi spores was higher within the soil under the canopy of M. biuncifera than in the soil under the canopy of P. laevigata. Thus, resource islands (RI) created by P. laevigata increased the amounts of SOC, TN and P when compared with the RI of M. biuncifera. This study provided evidences about the importance of species identity in order to expand the niche availability for the establishment of other plants, and highlights that P. laevigata and M. biuncifera jointly influencing plant colonization within semiarid ecosystems.
Key words: Mezquital Valley, mesquite, Mimosoideae, plant diversity, semiarid shrublands, soil nutrients.
Resumen
Prosopis laevigata y Mimosa biuncifera coexisten en los matorrales semiáridos; sin embargo, se desconoce su influencia sobre la diversidad de la comunidad vegetal y el suelo. Este estudio evaluó el efecto de P. laevigata y M. biuncifera sobre la estructura, diversidad vegetal y nutrimentos del suelo, en tres matorrales del Valle del Mezquital, México. Los sitios difieren en la especie dominante: Sitio 1, P. laevigata; Sitio 2, M. biuncifera y Sitio 3, ambas leguminosas. En cada sitio se recolectó suelo, tanto abajo y fuera del dosel de las leguminosas, además, se realizaron transectos para medir e identificar las plantas arbóreas y arbustivas, se calculó el índice de valor de importancia y la diversidad del matorral. Asimismo, se registró mayor riqueza y diversidad en el Sitio 3 (ICE 29 spp. y H’ 2.7), en comparación con el Sitio 1 (24 spp. y 2.4) y Sitio 2 (26 spp. y 2.1). La materia orgánica y el carbono orgánico del suelo, así como el N total, el P-Olsen y la mineralización de C fueron mayores en el suelo bajo dosel de ambas leguminosas. La abundancia de esporas de hongos micorrizógenos arbusculares fue favorecida por M. biunficera. La influencia de P. laevigata para crear islas más ricas en recursos fue mayor que en M. biunficera, lo anterior sugiere que cada leguminosa modifica de una forma diferente el microambiente, sin embargo, juntas aumentan la disponibilidad de nichos para el establecimiento de otras especies, lo queayuda a comprender el papel de P. laevigata y M. biuncifera sobre la colonización vegetal en ecosistemas semiáridos.
Palabras clave: diversidad vegetal, matorrales semiáridos, mezquite, Mimosoidae, nutrimentos, Valle del Mezquital.
Identity, richness and interactions among plant species affect the structure of any plant community and soil fertility, since species always differ in some of their functional traits (Chapin et al. 2002). Plants compete for water and nutrients, at the same time, they protect one another from extreme climates and provide additional resources by the incorporation of fresh litter for microbial activity, creating fertility islands (García-Moya & McKell 1970), or resource islands underneath their canopy (Reynolds et al. 1999), which favors the establishment of plant communities (Perroni- Ventura et al. 2006, López & Ortuño 2008). Keystone species are critical for these processes, because they have strongly influence on seed banks and seedling recruitment, even becoming the habitat of microorganisms, birds, rodents, and insects, whose role is critical for processes such as decomposition, spread, pollination, and plant establishment (Valiente- Banuet & Ezcurra 1991, Camargo-Ricalde & Dhillion 2003, Wright et al. 2006). These species frequently increase soil limiting resources and offer environmental conditions (i.e. higher humidity and lower soil temperature) for the growth of other plants under their canopies, favoring plant community diversity. However, these effects may vary among species and could be dependent on the ecological contexts (Maestre et al. 2003, Cortina & Maestre 2005).
Vegetation patterns in arid and semiarid ecosystems are distributed in mosaics, which may start through keystone plants that are able to modify the microenvironment (Aguiar & Sala 1999), as occurs with Acacia schaffneri (S. Watson) FJ Herm. (Yeaton & Romero- Manzanares 1986). Prosopis and Mimosa may become as keystone species in semiarid environments. Several studies have reported a nursing effect underneath the canopy of Prosopis articulata S. Wats. (Carrillo-García et al. 1999), P. laevigata (Humb. et Bonpl.) ex Willd (Cruz-Rodríguez et al. 1997, Frías-Hernández et al. 1999, Perroni-Ventura et al. 2006) and M. luisana Brandegee (Valiente-Banuet & Ezcurra 1991), recording a higher plant diversity under their canopies than in open areas. In addition, other studies have shown a major accumulation of soil organic matter and nutrients under the canopy of P. laevigata (Reyes-Reyes et al. 2002, Montaño et al. 2006), P. juliflora (Tiedemann & Klemmedson 1973, Herrera- Arreola et al. 2007), Mimosa biuncifera Benth (Herrera-Arreola et al. 2007), M. monancistra Benth. (Flores et al. 2007), M. lacerata Rose, M. luisana and M. texana (A. Gray) var. Filipes (Britton & Rose) Barneby (Camargo-Ricalde et al. 2002, Camargo-Ricalde et al. 2010a). These species improve fertility and create resource islands, due to that modify of the micro-environmental conditions and function as reservoirs of soil nutrients, fungi spores andmicroorganisms under their canopies (Reynolds et al. 1999, Camargo-Ricalde & Dhillion 2003, González-Ruiz et al. 2008). For instance, arbuscular mycorrhizal fungi are critical to plant establishment, because they improve water and nutrients status of the host plant growing in arid regions (Tarafdar & Panwar 2008, Camargo-Ricalde et al. 2010b). Even so, the effect of Prosopis and Mimosa species has been documented for each single species, without elucidating the interactions that could take place among them and with other plant species within the plant community. This point is critical to understand how different legume species could affect soil resources distribution and the surrounding plant community.
As a result of landscape fragmentation, Prosopis laevigata and Mimosa biuncifera can be found together or separated forming patches within plant communities in Mezquital Valley, central Mexico. These patches offer an excellent opportunity to examine how both legumes contribute to plant diversity and soil fertility in semiarid shrublands. Therefore, the aim of this study was to explore how the jointly influenceof P. laevigata and M. biuncifera, can be related to: 1) the community plant structure where they occurred, 2) trees and shrubs diversity, and 3) soil properties, in a semiarid ecosystem located in the Mezquital Valley, Hidalgo state, Mexico.
Materials and Methods
Study area: The study area is located in the Mezquital Valley, Hidalgo state, México. Mean annual temperature is 24°C and mean annual rainfall is 550mm, is strongly seasonal, with seven dry months, and a rainy season occurring from June to October (INEGI 1987). There are two main types of soils, Pellic Vertisols and Rendzic Leptosol (FAO-WRB 2006), both derived from sedimentary marine rocks; pH varies from 7.5-8.2 (Gonzalez-Quintero 1968). The vegetation type is a semiarid shrubland (“matorral xerófilo”) (Rzedowski 1978), where P. laevigata and M. biuncifera are either dominant or co-dominant within their communities. Both species have a high value silvopasture for local people; they use them as a source of wood for fuel and construction, shadow and fodder for livestock, and as living fences (Galindo & García-Moya 1986, Camargo-Ricalde et al. 2001).
Site selection and experimental design: Three study sites located in the Mezquital Valley were chosen in relation to the presence of P. laevigata and M. biuncifera (Leguminosae- Mimosoideae). The three sites have a Rendzic Leptosol soil type, and similar vegetation type, but differ in dominant plant species. Site 1 was located near Bingu town (20°36’50’’- 20°36’56’’ N and 99°06’55.5’’-99°07’05’’ W), where P. laevigata is the dominant species. Site 2 was located in the vicinity of González town (20°24’30’’-20°24’35’’ N and 98°58’03’’-98°58’11’’ W), where M. biuncifera is the dominant species. Site 3 was close to Rincón town (20°16’16’’-20°16’20’’ N and 98°54’34’’-98°54’46’’ W), where both legumes are co-dominant species. We contrasted these sites to elucidate a possible relation between both P. laevigata and M. biuncifera on plant community and soil properties.
Vegetation sampling and variables: Eight transects of 10x20m (1 600m2) were established per site, transects were set 25m apart from each other. All shrubs and trees were collected for identification; voucher specimens were deposited at the FEZA herbarium. The data from all transects was used to calculate absolute and relative density (ind./ha), canopy cover (m2/ha), plant frequency, and the importance
value (IV) per species (Brower & Zar 1990). The species richness and shared species among sites were also measured. In order to describe the structural attributes of P. laevigata and M. biuncifera, density of both plant species were also estimated; though only eight plants per species were selected randomly per site, were measured for plant height, cover, as well as distance between plants for each species.
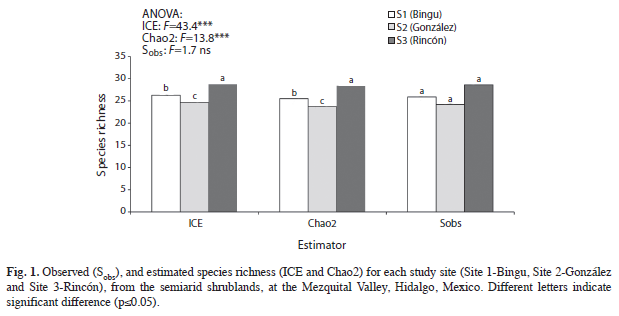
The species richness was assessed as the number of plant species observed per site (Sobs), and to approach the true species richness, two non-parametric estimators provided by the EstimateS program (Colwell 2009) were applied for the three study sites. The estimators of species richness were incidence-based coverage (ICE) and Chao 2 based on the incidence (presence/absence) of species. Species diversity was estimated by the Shannon-Weiner (rare species) and Simpson (dominant species) indexes, and by the cumulative number of species, as a function of the cumulative number of plants (rarefaction curves), which were estimated using EstimateS; in order to examine the floristic similarity among the study sites, the Jaccard similarity index (JI) was calculated, based on species presence/absence data, where JI=1 in cases of complete similarity between pairs of sites, and 0% if the sites have no species in common (Magurran 2004).
Soil sampling and analysis: Soil samples were collected under the canopy (UC) of eight plants of similar size of both P. laevigata and M. biuncifera per site; each plant was considered as a replicate and a distance of at least 15m among plants was established. Soil samples, from the surface to a depth of 15cm, were collected during the dry season, May 2006, where most of the root biomass and soil organic matter were concentrated (Montaño et al. 2006, Pavón 2007). The soil samples were collected from the middle of the canopy of each plant, while, other soil samples were collected from eight open areas without vegetation (OA), located outside the canopy of P. laevigata or M. biuncifera. A total of 64 soil samples were collected, stored in black plastic bags and refrigerated at 4°C until laboratory analysis.
Soil samples were air dried and sieved (2mm) previous to analysis. Bulk density was determined with the test tube method (Jury et al. 1991). Soil pH was measured in a soil:distilled water suspension (1:2 w/v) with a pH meter equipped with a glass electrode. The electrical conductivity (CE) was measured in a 1:5 soil-water suspension, using a conductivity cell (NOM 2000). Texture was estimated by the Bouyocus hydrometer method as described by Gee & Bauder (1986). SOM was quantified by the Walkley-Black procedure, and soil organic carbon (SOC) was estimated as of SOM by assuming that it contains 58% of total organic carbon (Nelson & Sommers 1996). Soil TN was determined after an acid digestion by a semi-micro-Kjeldahl method followed by a colorimetrical analysis (Bremmer & Mulvaney 1982). Available phosphorus (PO4) was measured using the methodology described by Olsen et al. (1954). Exchangeable calcium (Ca, λ=422.7nm), magnesium (Mg, λ=285.2nm) and potassium (K, λ=766.5nm) were extracted
using 1N ammonium acetate (NOM 2000), and then measured using an Atomic Absorption Spectrophotometer (Varian Spectrum-A).
The carbon mineralization (CO2) was measured after five days of aerobic incubations at 28°C; each sub-sample was periodically wetted with deionized water to reach field water holding capacity. The CO2 evolved was collected in 0.5N NaOH traps. Carbonates were precipitated by adding 1N KCl2 and then titrated with 0.5N HCl using phenolphtaleine as indicator (Robertson et al. 1999). The spores of arbuscular mycorrhizal fungal were extracted from 100g of dry soil using a wet-sieving and density gradient procedure, with a modification of sucrose-centrifugation technique (Daniels & Skipper 1982). The intact spores, with filled cytoplasm, were counted under a dissecting microscope at 5X. Spore density was expressed as the spore number per 100g of soil. All values were expressed on a dry-weight basis unless otherwise stated.
A one-way ANOVA was used to examine differences among sites in relation to vegetation variables. Soil variables were analyzed using a randomized nested ANOVA, where the factors were the species (P. laevigata and M. biuncifera) and the microenvironments (under the canopy of the legumes, and open area) nested within the sites (Site one-Bingu, Site 2-González and Site 3-Rincón). In all the cases, when the ANOVA indicated a significant factor, means were compared with a Tukey post-hoc test. Normality test was examined for all data, and when a normal distribution and homogeneity of variance were not found, the data were log-transformed to meet ANOVA assumptions (Sokal & Rohlf 1995), but reported in their original scale of measurement. All statistical analyses were performed with Statistica 6 software (StatSoft 2000), and in all cases p≤0.05 was taken to be significant. A Canonical Correspondence Analysis (CCA) was used to search differences in the floristic composition patterns among sites, legume species and microenvironments. In this analysis, a data matrix containing the IV for each plant species, per eight sites was related with a second matrix based on seven soil variables per site. The soil variables used in the CCA were only those without an auto-correlation. Soil variables were onward selected employing the Monte Carlo permutation test, and this analysis was performed in PC-ORD software (ter Braak 1990, McCune & Mefford 1999).
Results
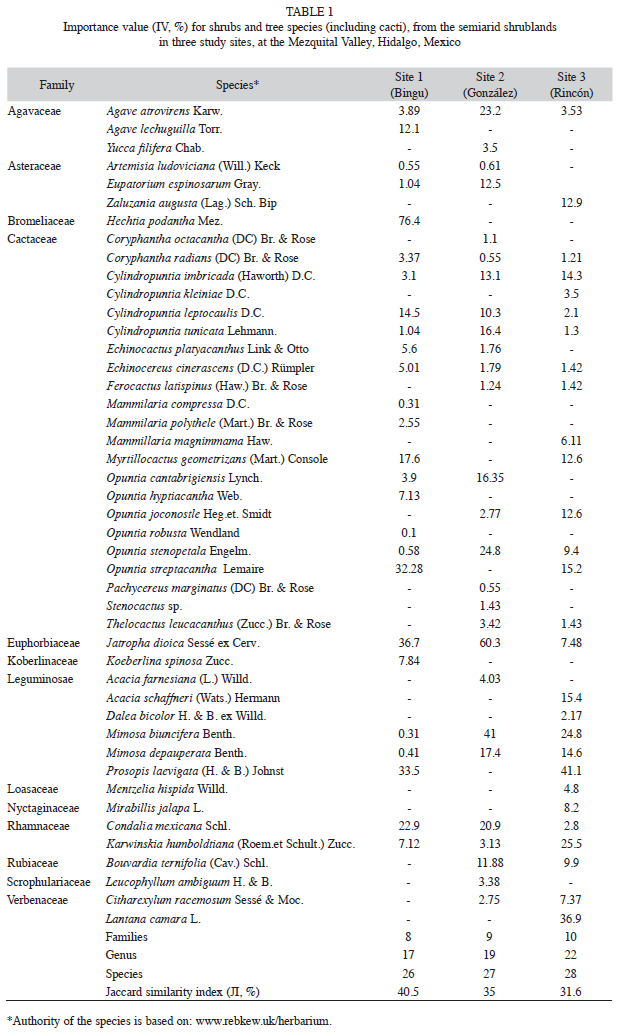
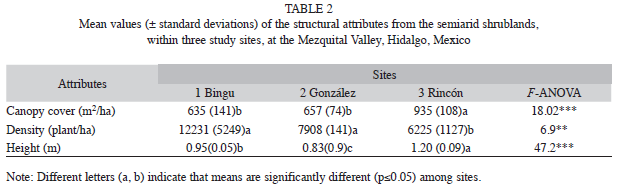
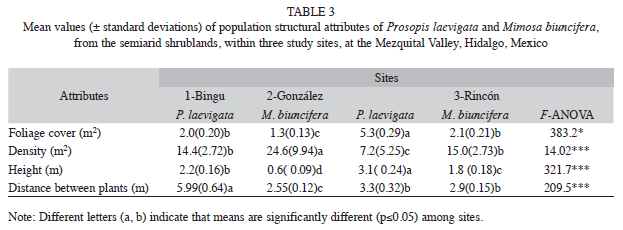
Vegetation structure: A total of 45 shrub and tree species belonging to 31 genera and 13 families were recorded in all three sites (Table 1). Cactaceae and Leguminosae were the best represented. Both plant families represented 65.4% of the total of species collected in Site 1, 55% in Site 2, and 64.3% in Site 3. Cactaceae had the greatest species richness, and Loasaceae (Site 3) and Scruphulariaceae (Site 2) had the lowest species richness; hence, they got a very low IV (≤5%) compared to other plant families in the three sites, which also differed from 31% to 40% in similarity of plant species (Table 1). Site 3 had 1.4 times more canopy cover and plant height than the other two sites. In contrast, density was higher in Site 1 and Site 2 than in Site 3 (Table 2). Prosopis laevigata and M. biuncifera showed differences in their structural attributes. For instance, both legumes had bigger cover and height, but lower density when both coexist (Table 3).
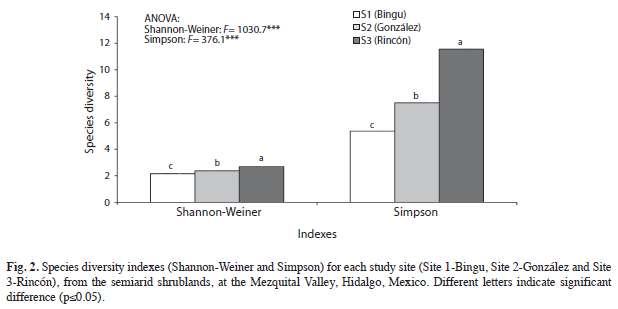
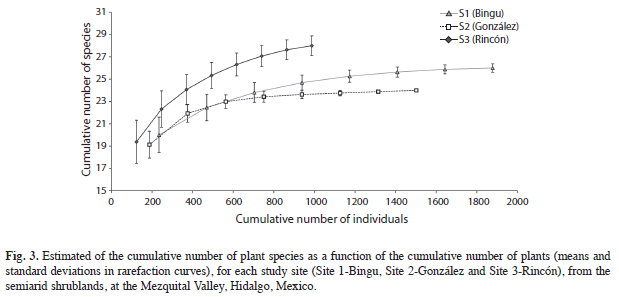
Species richness and diversity: The nonparametric estimators (ICE and Chao2) indicated significant statistical differences (p≤0.05) in species richness among sites; but no significant differences were pointed out in Sobs (Fig. 1). Site 2 showed lower richness and diversity of species that Site 1; while Site 3 had the highest diversity of species (Fig. 2) and, the rarefaction curves are showed in figure 3.
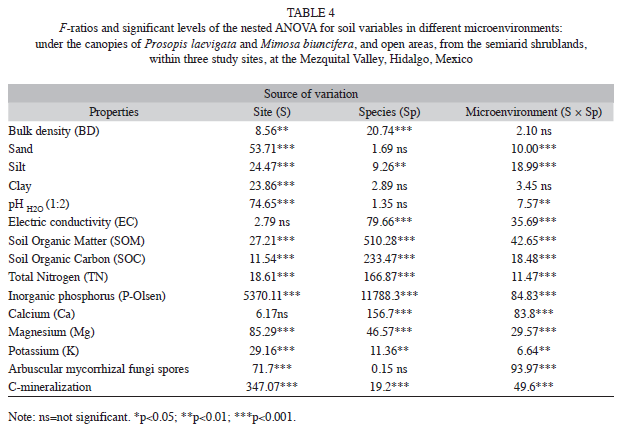
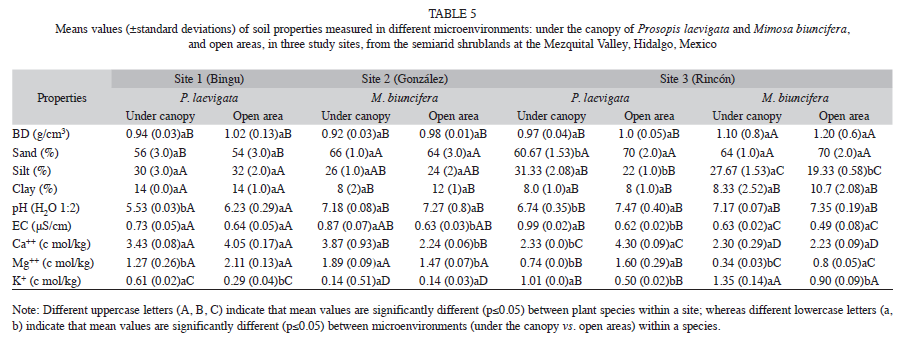
Influence of P. laevigata and M. biuncifera on soil properties: In Site 3, the soil under the canopy of M. biuncifera showed the highest bulk density, lower sand and higher silt content than in open areas (Tables 4 and 5). The most common soil textural class was sandy loam. Soil pH varied among sites and microenvironments; though it did not vary between species, tending from neutral to slightly alkaline except in Site 1 (Table 5). Soil pH under the canopy of P. laevigata was more acid than in open areas in sites 1 and 3; however, it was not affected by M. biuncifera (Table 4). In sites 1 and 2, the EC of the soil under the canopy of P. laevigata was lower than the soil under the canopy of M. biuncifera; in contrast, in Site 3 the soil under the canopy of P. laevigata had the highest EC when compared to M. biuncifera (Tables 4 and 5).
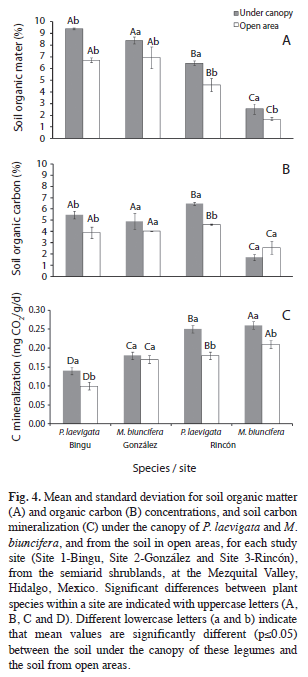
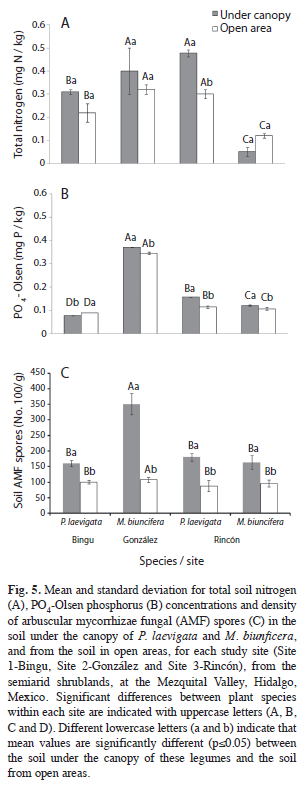
Soil organic matter (SOM) and soil organic carbon (SOC) showed similar patterns among sites and species (Table 5, Fig. 4A and 4B). Thus, sites 1 and 2 had higher SOM and SOC concentrations than Site 3. Particularly in Site 3, SOM and SOC under the canopy of P. laevigata were higher than the soil under the canopy of M. biuncifera. In the three study sites, SOM concentrations were higher under the canopy of both legumes than in open areas. In contrast, SOC concentrations were higher under the canopy of P. laevigata than in open areas, but this was not found for M. biuncifera (Fig. 4B). In Site 3, P. laevigata had higher soil N concentration than the soil under the canopy of M. biuncifera; however, when these legumes occur in different sites, P. laevigata had lower N concentration than M. biuncifera. In addition, in Site 3, soil N concentration was higher under the canopy of P. laevigata than in open areas (Table 5, Fig. 5A). Soil available P was higher in Site 2 than in the other two sites. The soil under the canopy of P. laevigata and M. biuncifera had higher available P than the soil in open areas, with the exception of P. laevigata in Site 1 (Table 5, Fig. 5B).
Soil Ca and Mg concentrations were higher in Sites 1 and 2; P. laevigata had higher concentrations of both cations under its canopy than M. biuncifera. Soil Ca and Mg concentrations were lower under the canopy of P. laevigata than in open areas; on the contrary, the concentration of these soil cations was higher under the canopy of M. biuncifera than in open areas (Tables 4 and 5). Soil K concentration was higher under the canopy of both legumes than in open areas in the three study sites (Tables 4 and 5).
Influence of P. laevigata and M. biuncifera on soil biological activity: Site 3 registered the highest C mineralization. The soil under the canopy of P. laevigata had lower C mineralization than the soil under the canopy of M. biuncifera. Soil C mineralization under the canopy of P. laevigata and M. biuncifera was higher than in open areas, with exception of M. biuncifera in Site 2 (Tables 4 , Fig. 4). The abundance of arbuscular mycorrhizal (AM) fungal spores was higher in Site 2. There was a higher abundance of AM fungal spores in the soil under the canopy of both legumes in the three sites than in open areas; though in Site 2, M. biuncifera had three times more spores in the soil under its canopy than in open areas (Table 4, Fig. 5).
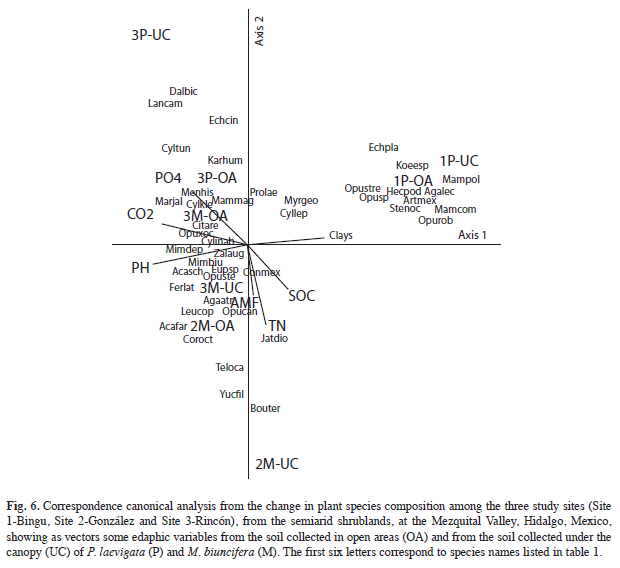
Relation among plant community, soil properties and the presence of legumes: CCA canonical axes together accounted for 75% of the variation in the data, with 29, 24 and 22% respectively. The first axis was positively correlated with clays (0.70) and negatively with pH (-0.92) and CO2 (-0.85). This indicates that Site 1 was different from sites 2 and 3. The first one is characterized by Hechtia podantha, Agave lechuguilla, Mammilaria compresa and M. polytele, while the other two sites are characterized by Mimosa depauperata, Opuntia joconostle, Cylindropuntia kleiniae, M. biuncifera and Coryphantha radians. The second axis was correlated with TN (-0.76), PO4 (0.42) and AM fungal spores (-043), and separated Sites 1 and 3 where P. laevigata occurs, from Site 2 where M. biuncifera is alone; while open areas were more closely related among them (Fig. 6).
Discussion
The overall results showed that the presence of both, P. laevigata and M. biuncifera within the plant community of Site 3 improve the plant structure-diversity due to a complementary effect, compared to the other two sites where the legumes occurred separately. Both legume species may directly increase the colonization of associated perennial plants by sheltering them from high temperatures, and by providing a richer microenvironment in resources than in the other two sites. Higher species richness has been reported under the canopy of P. laevigata in semiarid shrublands in Northern Guanajuato state (Cruz-Rodríguez et al. 1997) and in the Tehuacán-Cuicatlán Valley, Puebla, Mexico (Perroni-Ventura et al. 2006), as well as in other desert ecosystems where Prosopis species occur (Carrillo-García et al. 1999, Gómez-Aparicio et al. 2005, López & Ortuño 2008). Likewise, some Mimosa species act as nurse plants for cacti species (Valiente-Banuet & Ezcurra 1991, Reyes-Olivas et al. 2002) and perennial plants such as Jatropha dioica (Yeaton & Romero-Manzanares 1986). Our results suggest that, within the Mezquital Valley, P. laevigata and M. biuncifera may increase plant diversity because they may act jointly as focal plants for seeds arrival, the subsequent seedling establishment and the survival of adult plants. This agrees with the hypothesis proposed by Aguiar & Sala (1999), which states that, the spaces around and underneath woody species may be progressively colonized by associated plants until a patch of vegetation is built.
Influence of P. laevigata and M. biuncifera on the plant community structure and on the soil properties under their canopies also takes place when both legumes are growing in different sites, although, it is less effective, presumably due to the fact that there is not any complementary influence of both legumes (Brooker et al. 2008). P. laevigata is a deciduous species that deposits certain amounts of organic matter in the soil, which probably is used by opportunistic plant species, reducing the establishment of non-opportunistic ones. In Site 1, H. podantha and J. dioica had 33% of IV within this plant community. These two species have a high capacity of vegetative propagation; which allows a quick capture of resources. In Site 2, M. biuncifera also favors the establishment of J. dioica; both plant species are able to spread after disturbance, to colonize greatly eroded soils and to tolerate low soil nutrient levels (Yeaton & Romero-Manzanares 1986, Luna-Suárez et al. 2000); such attributes are traits that make both species to be more successful than other plants. We suggest that the combination of these biological attributes associated to these plants may explain the low species diversity reported in the sites 1and 2, with different focal legumes.
In Site 3, the resources islands produced by P. laevigata and M. biunficera possibly create a broader range of available niches for other plant species as suggested by Diamond & Case (1986), Tilman (2004), Cortina & Maestre (2005). This agrees with other studies (Maestre & Cortina 2005, Brooker et al. 2008) showing that the soils with more available resources favor the establishment of a greater number of plant species. For instance, in a Mexican semiarid ecosystem, Perroni-Ventura et al. (2006) found that plant richness is related to those nutrients linked to the dynamics of organic matter incorporated by P. laevigata. We propose that in Site 3 the coexistence of both legumes promote a positive feedback on soil fertility, which is reflected on characteristics of plant community. Prosopis laevigata and M. biuncifera lead to more favorable conditions for plant growth under their canopies and around, due to resource enrichment, resulting presumably of root and litter deposition (Pavón et al. 2005, Perroni-Ventura et al. 2006, Flores et al. 2007, Camargo-Ricalde et al. 2010a).
The opposite situation occurs in open areas, due to the almost null presence of plants, which usually are small with little cover, and to higher loss of organic material caused by wind and water erosion. This observation agrees with Reynolds et al. (1999), who mentioned that, in arid and semiarid ecosystems, the spatial distribution of soil resources is linked to plant canopy cover. Consequently, more canopy cover, bigger plant size, higher plant richness and diversity, and a closer plant to plant distance, might create microenvironments with higher soil organic matter and nutrients, and more soil microorganisms (Herman et al. 1995, Reynolds et al. 1999, Maestre & Cortina 2005, Montaño et al. 2006, González-Ruiz et al. 2008).
The generation of a milder microenvironment as in Rincón (Site 3), rich in organic matter and soil nutrients, promotes the microbial activity within the site having-as a consequence- a decrease in SOM due to a greater C mineralization. However, C mineralization varied in relation to the identity of the focal legume. For instance, P. laevigata produces higher quantity and quality of litter than M. biuncifera, which explains why C mineralization was higher under the canopy of P. laevigata than in open areas within the three study sites. Herrera-Arreola et al. (2007) support our findings, they reported that the leaves of M. biuncifera have lower N concentration and higher lignin and polyphenol content than the leaves of P. juliflora, and by Reyes-Reyes et al. (2003), who also found that an input of P. laevigata leaves into the soil increases two times the C mineralization in relation to M. biuncifera leaves input. In addition, P. laevigata and M. biuncifera, establish a symbiotic relationship with N-fixing Rhizobium bacteria, thus increasing the N content of their litter (Frías-Hernández et al. 1999, Pavón et al. 2005, González-Ruiz et al. 2008). Nevertheless, both species have a differential impact on soil N; a higher quantity and quality of litter may explain the accumulation of N in the soil under the canopy of P. laevigata than that reported in open areas or in the soil under the canopy of M. biuncifera, even when both species are coexisting in the same place (Site 3). Additionally, P. laevigata probably has a mechanism that depleted soil Ca and Mg concentration, which is absent or not so efficient in M. biuncifera (Camargo-Ricalde et al. 2010a). A possible mechanism may be attributed to both legumes, which differ in their production of organic acids and accumulation of organic residues under their canopies decreasing soil pH (Finzi et al. 1998). Other studies have also documented this trend in the soils under the canopy of P. laevigata and M. monancistra than in open areas (Frías-Hernández et al. 1999, Montaño et al. 2006, Flores et al. 2007). Altogether, these facts suggest that P. laevigata have a higher impact on soil nutrients than M. biunficera within the Mezquital Valley.
The highest plant diversity and biological activity in Site 3, apparently decreased soil P availability; it is possible that the overall of plants within the community could have used the soil inorganic P to supply their nutrimental demands. A lower abundance of AM fungal spores in sites 1 and 3 than in Site 2, suggests that these fungi might be establishing an active association with the plants, which depleted soil available P pool. AM fungi may be advantageous to desert plants, when P is practically insoluble, and its diffusion in the soil is further decreased by low soil moisture (Tarafdar & Panwar 2008). Furthermore, P availability under the canopy of both legumes may also be favored by a low soil Ca and Mg concentration, and a slightly acid pH, which agrees with a higher AM fungal spore abundance in the soil under the canopies of both legumes (Camargo-Ricalde et al. 2010b). Smith & Read (2008) indicate that the plant-root colonization by AM fungi decreases when P availability is high in the soil, increasing AM fungal spore production.
Within the three study sites, both legumes mainly modify the soil properties linked to biological processes. This argument is supported by the Canonical Correspondence Analysis that pointed out that SOC, CO2-respiration, soil pH, AM fungal spores, total N and available P were correlated with the actual distribution of the plant species in the neighborhoods of both legumes, which increase soil resources, influencing the structure these plant communities, by favoring diverse niches for seeds or plant and AM fungal propagules.
In conclusion, this study provides evidences that may help to understand the role of P. laevigata and M. biuncifera on plant colonization within semiarid ecosystems. Both legumes jointly modify soil resources; this fact could be reflected in the high plant diversity within the semiarid shrublands at the Mezquital Valley. However, further studies are needed to test this hypothesis and to evaluate in more detail the vegetation dynamics as a function of scrubs patches. The influence of P. laevigata to create resources islands was higher than the one showed M. biuncifera, generating broader niche availability for the establishment of other plants. The conservation of both legumes may be used for biodiversity and soil conservation programs, accelerating plant succession processes within this semiarid ecosystem.
Acknowledgments
We thank Mariano García Díaz, Gabriela de Jesús Espino Ortega, Luis Emilio de la Cruz López, Susana Adriana Montaño-Arias, Jazmín González Rivera, Israel Castro López and Gilberto Platas for their help in the field-work. We would like to thank Eduardo Chimal Sánchez for his technical support in the laboratory. This research was supported by the Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (IN-216610), Dirección General de Asuntos del Personal Académico, Universidad Nacional Autónoma de México (UNAM). This paper was part of R. García- Sánchez doctoral thesis, Posgrado en Botánica, Colegio de Postgraduados, Campus Montecillo.
References
Aguilar, M.R. & O.E. Sala. 1999. Patch structure dynamics and implications for the functioning of arid ecosystems. Tree 14: 273-277. [ Links ]
Bremmer, J.M. & C.S. Mulvaney. 1982. Total nitrogen, p. 595-624. In A.L. Page, R.H. Miller & D.R. Keeney (eds.). Methods of soil analysis. Part 2. Amer. Soc. Agron. Madison, Wisconsin, USA. [ Links ]
Brooker, R.W., F.T. Maestre, R.M. Callaway, Ch. L. Lortie, L.A. Cavieres, G. Kunstler, P. Liancourt, K. Tielbörger, J.M.J. Travis, F. Anthelme, C. Armas, L. Coll, E. Corcket, S. Delzon, E. Forey, Z. Kikvidze, J. Olofsson, F. Pugnaire, C.L. Quiroz, P. Saccone, K. Schiffers, M. Seifan, B. Touzard & R. Michalet. 2008. Facilitation in plant communities: the past, the present and the future. J. Ecol. 96: 18-34. [ Links ]
Brower, J.E. & J.H. Zar. 1990. Field and laboratory methods for general ecology. Brown, Iowa, USA. [ Links ]
Camargo-Ricalde, S.L. & S.S. Dhillion. 2003. Endemic Mimosa species can serve as mycorrhizal “resource islands” within semiarid communities of the Tehuacán-Cuicatlán Valley, Mexico. Mycorrhiza 13: 129-136. [ Links ]
Camargo-Ricalde, S.L., S.S. Dhillion & R. Grether. 2002. Community structure of endemic Mimosa species and environmental heterogeneity in a semiarid Mexican Valley. J. Veg. Sci. 13: 697-704. [ Links ]
Camargo-Ricalde, S.L., I. Reyes-Jaramillo & N.M. Montaño. 2010a. Forestry insularity effect of four Mimosa L. species (Leguminosae-Mimosoideae) on soil nutrients of a Mexican semiarid ecosystem. Agrof. Syst. 80: 385-397. [ Links ]
Camargo-Ricalde, S.L., N.M. Montaño, I. Reyes-Jaramillo, C. Jiménez-González & S.S. Dhillion. 2010b. Effect of mycorrhizae on seedlings of six endemic Mimosa L. species (Leguminosae-Mimosoideae) from the
semi-arid Tehuacán-Cuicatlán Valley, Mexico. Tress Strc. Func. 24: 67-78. [ Links ]
Camargo-Ricalde, S.L., R. Grether, A. Martínez-Bernal, V. García-García & S. Barrios-del- Rosal. 2001. Especies útiles del género Mimosa (Fabaceae-Mimosoideae) en México. Bol. Soc. Bot. Méx. 68: 33-44. [ Links ]
Carrillo-García, A., J.L. León, Y. Bashan & G.J. Bethlenfalvay. 1999. Nurse plants mycorrhizae and plant establishment in disturbed area of the Sonoran desert restoration. Ecol. Res. 4: 321-335. [ Links ]
Chapin, F.S. III, P.A. Matson & H.A. Mooney. 2002. Principles of terrestrial ecosystem ecology. Springer, New York, New York, USA. [ Links ]
Colwell, R.K. 2009. EstimateS V6 01b1. Statistical estimation of species richness and shared species richness and shared species from samples. User Guide and Application. Connecticut, USA. [ Links ]
Cortina, J. & T.F. Maestre. 2005. Plants effects on soil in dry lands implications for community dynamics and ecosystem restoration, p 84-118. In D. Binkley & O. Menyailo (eds.). Tree species effects on soils: Implications for Global Change NATO Science Series. Kluwer Academic, Dordrecht, The Netherland. [ Links ]
Cruz-Rodríguez, J.A., E. García-Moya, J.T. Frías-Hernández, G. Montesinos & J.L. Flores. 1997. Influencia de los mezquites en la composición y cobertura de la vegetación herbácea de un agostadero semiárido del Norte de Guanajuato. Bol. Soc. Bot. Méx. 61: 21-30. [ Links ]
Daniels, N. & H. Skipper. 1982. Methods for the recovery and quantitative estimation of propagules from soil, p. 29-35. In Methods and principles of mycorrhizal research. American Phytopathology Society, Washington, D.C., USA. [ Links ]
Diamond, J. & T.J. Case. 1986. Community Ecology. Harper & Row, New York, USA. [ Links ]
FAO (Food and Agriculture Organization). 2006. Word reference base for soil resources (WRB). World Soil Resources Reports 103 Rome, Italy. [ Links ]
Finzi, A.C., C.D. Canham & N.V. Breemen. 1998. Canopy tree-soil interactions within temperate forest: species effects on pH and cations. Ecol. Appl. 8: 447-454. [ Links ]
Flores, E., J. Frías, P. Jurado, V. Olalde, J.D. Figueroa, A. Valdivia & E. García-Moya. 2007. Efecto del gatuño sobre la fertilidad del suelo y la biomasa herbácea en pastizales del centro de México. Terra Lat. 25:
311-319. [ Links ]
Frías-Hernández, J.T., L.L. Aguilar, V.P. Olalde, J.A. Balderas, L.G. Gutiérrez, J.J. Alvarado, J. Castro, H. Vargas, A. Albores & L. Dendooven. 1999. C and N soil characteristics in a semiarid highlands of Central Mexico as affected by mesquite trees (Prosopis laevigata). Arid Soil Res. Rehab. 13: 305-312. [ Links ]
Galindo, A.S. & E. García-Moya. 1986. The uses of mesquite (Prosopis spp.) in the highlands of San Luis Potosi, Mexico. For. Ecol. Manag. 16: 49-56. [ Links ]
García-Moya, E. & C.M. McKell. 1970. Contribution to the nitrogen economy of a desert-wash plant community. Ecology 51: 81-88. [ Links ]
Gee, G.W. & J.W. Bauder. 1986. Particle size analysis, p. 383-411. In A. Klute (ed.). Methods of soil analysis Vol I Physical and mineralogical methods. Amer. Soc. Agron. Madison, Wisconsin, USA. [ Links ]
Gómez-Aparicio, L., J.M. Gómez, R. Zamora & J.L. Boettinger. 2005. Canopy vs soil effects of shrubs facilitating tree seedlings in Mediterranean montane ecosystems. J. Veg. Sci. 16: 191-198. [ Links ]
González-Quintero, L. 1968. Tipos de vegetación del Valle del Mezquital, Hidalgo. Departamento de Prehistoria, Instituto Nacional de Antropología e Historia, México. [ Links ]
González-Ruiz, T., Z.S. Rodríguez & R. Ferrera-Cerrato. 2008. Fertility islands around Prosopis laevigata and Pachycereus hollianus in the drylands of Zapotitlán Salinas, México. J. Arid Environ. 72: 1202-1212. [ Links ]
Herman, R.P., K. Provencio, J. Herrera-Matos & R. Torrez. 1995. Resource islands predicts the distribution of heterotrophic bacteria in Chihuahuan desert soils. Appl. Environ. Microb. 61: 816-1821. [ Links ]
Herrera-Arreola, G., Y. Herrera, B.G. Reyes-Reyes & L. Dendooven. 2007. Mesquite (Prosopis juliflora (Sw) DC), huisache (Acacia farnesiana (L) Willd.) and catclaw (Mimosa biuncifera Benth.) and their effect on dynamics of carbon and nitrogen in soils of the semi-arid highlands of Durango Mexico. J. Arid Environ. 69: 583-598. [ Links ]
INEGI (Instituto Nacional de Estadística Geografía e Informática). 1987. Carta climática Hoja Pachuca del estado de Hidalgo. Escala 1:250 000. Aguascalientes, Aguascalientes, México. [ Links ]
Jury, W.A., W.R. Gardner & W.H. Gardner. 1991. Soil Physics. John Willey & Sons, New York, USA. [ Links ]
López, R.P. & T. Ortuño. 2008. La influencia de los arbustos sobre la diversidad y abundancia de plantas herbáceas de la Prepuna a diferentes escalas espaciales. Ecol. Aus. 18: 119-131. [ Links ]
Luna-Suárez, S., J.T. Frías-Hernández, V. Olalde-Portugal & L. Dendooven. 2000. Catclaw (Mimosa biuncifera): a pest or means to restore soil fertility in heavily erodes soil from the fertility in heavily eroded soil from the central highlands of Mexico? Biol. Fert. Soils 32: 109-113. [ Links ]
Maestre, F.T., S. Bautista & J. Cortina. 2003. Positive, negative, and net effects in grass-shrub interactions in Mediterranean semiarid grasslands. J. Ecol. 12: 3186-3197. [ Links ]
Maestre, F.T & J. Cortina. 2005. Remnant shrubs in Mediterranean semi-arid steppes: effects of shrub size, abiotic factors and species identity on understory richness and occurrence. Act. Oecol. 27: 161-169. [ Links ]
Magurran, A.E. 2004. Measuring biological diversity. Blackwell Sci, Victoria, United Kingdom. [ Links ]
McCune, B. & M.J. Mefford. 1999. PC-ORD. Multivariate analysis of ecological data, version 4. MjM software design. Gleneden Beach, Oregon, USA. [ Links ]
Montaño, N.M., R. García-Sánchez, G. Ochoa de la Rosa & A. Monroy-Ata. 2006. Relación entre la vegetación arbustiva, el mezquite y el suelo de un ecosistema semiárido en México. Terra Lat. 24: 193-205. [ Links ]
Nelson, D. & L. Sommers. 1996. Total carbon, organic carbon, and organic matter, p. 961-1010. In A. Miller & D. Keeney (eds.). Methods of soil analysis. Part II. Amer. Soc. Agron. Madison USA. [ Links ]
NOM. 2000. Norma Oficial Mexicana NOM-021-RECNAT- 2000. Establece las especificaciones de fertilidad, salinidad y clasificación de suelos. Estudio, muestreo y análisis. Norlex internacional, Edición electrónica de leyes, México. [ Links ]
Olsen, S.R., C.V. Cole, F.S. Watanabe & L.A. Dean. 1954. Estimation of available phosphorus in soil by extraction with sodium bicarbonate. USDA. Circ. 939 U.S. Washington, D.C., USA. [ Links ]
Pavón, N.P. 2007. Fine root biomass and production in a semiarid Mexican shrubland. The Southwestern Nat. 52: 116-167. [ Links ]
Pavón, N.P., O. Briones & J. Flores-Rivas. 2005. Litterfall production and nitrogen content in an intertropical semi-arid Mexican scrub. J. Arid Environ. 60: 1-13. [ Links ]
Perroni-Ventura, Y., C. Montaña & F. García-Oliva. 2006. Relationship between soil nutrient availability and plant species richness in a tropical semi-arid environment. J. Veg. Sci. 17: 719-728. [ Links ]
Reyes-Olivas, A., E. García-Moya & L. López-Mata. 2002. Cacti-shrub interactions in the coastal desert of northern Sinaloa, Mexico. J. Arid Environ. 52: 431-445. [ Links ]
Reyes-Reyes, G., L. Barón-Ocampo, I. Cuali-Álvarez, J.T. Frías-Hernández, V. Olalde-Portugal, L. Varela & L. Dendooven. 2002. C and N dynamics in soil from the central higlands of Mexico as affected by mesquite (Prosopis spp.) and huizache (Acacia tortuosa): a laboratory investigation. Appl. Soil Ecol. 19: 27-34. [ Links ]
Reyes-Reyes, G.B., E. Zamora-Villafranco, M.L. Reyes- Reyes, J.T. Frías-Hernández, V. Olalde-Portugal & L. Dendooven. 2003. Decomposition of leaves of huizache (Acacia tortuosa) and mesquite (Prosopis spp) in soil of the highlands of Mexico. Plant Soil 256: 359-370. [ Links ]
Reynolds, J.F., R.A. Virginia, P.R. Kemp, A.G. de Soyza & D.C. Tremmel. 1999. Impact of drought on desert shrubs: effects of seasonality and degree of resource islands development. Ecol. Monog. 69: 69-106. [ Links ]
Robertson, P.G., D.C. Coleman, C.S. Bledsoe & P. Sollins. 1999. Standard soil methods for long-term ecological research (LTER). Oxford University, New York, USA. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa, México D.F., México. [ Links ]
Smith, S.E. & D.I. Read. 2008. Mycorrhizal symbiosis. Academy, San Diego, USA. [ Links ]
Sokal, R. & F. Rohlf. 1995. Biometry. Freeman, San Francisco, California, USA. [ Links ]
StatSoft, INC. 2000. Statistica for Windows vers. 6. Tulsa. Oklahoma, Tulsa, USA. [ Links ]
Tarafdar, J.C. & J. Panwar. 2008. Role of arbuscular mycorrhizae in P nutrition of plants under arid and semiarid environments, p. 41-53. In N.M. Montaño, S.L. Camargo-Ricalde, R. García-Sánchez & A. Monroy- Ata (eds.). Arbuscular mycorrhizae in arid and semiarid ecosystems. Mundi-Prensa, INE-SEMARNAT, UAM-Iztapalapa, FES-Zaragoza UNAM, México D.F., México. [ Links ]
ter Braak, C.J.F. 1990. Update notes for CANOCO program version 3.10. Agricultural Mathematics Group, Wageningen, USA. [ Links ]
Tiedemann, A.R. & J.O. Klemmedson. 1973. Effect of Mesquite on Physical and Chemical Properties of the Soil. J. Rang. Manag. 26: 27-29. [ Links ]
Tilman, D. 2004. Niches tradeoffs, neutrality, and community structure: A stochastic theory of resource competition, invasion, and community assembly. PNAS 30: 10854-10861. [ Links ]
Valiente-Banuet, A. & E. Ezcurra. 1991. Shade as a cause of the association between the cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisiana in the Tehuacán Valley. México. J. Ecol. 79: 961-971. [ Links ]
Wright, J.P., C.G. Jones, B. Boeken & M. Shachak. 2006. Predictability of ecosystems engineering effects on species richness across environmental variability and spatial scales. J. Ecol. 94: 815-824. [ Links ]
Yeaton, R.I. & A. Romero-Manzanares. 1986. Organization of vegetation mosaics in the Acacia shaffneri-Opuntia streptacantha association, southern Chihuahuan Desert, Mexico. J. Ecol. 74: 211-217. [ Links ]
*Correspondencia:
Rosalva García-Sánchez: Posgrado en Botánica, Colegio de Postgraduados, C.P. 56230. Montecillo, Texcoco, Estado de México, México; rosalbagarcia@colpos.mx. Unidad de Investigación en Ecología Vegetal, Facultad de Estudios Superiores Zaragoza (FES Zaragoza), Universidad Nacional Autónoma de México. AP. 09230, D.F., México.
Sara Lucía Camargo-Ricalde: Departamento de Biología, División de Ciencias Biológicas y de la Salud, Universidad Autónoma Metropolitana, Unidad Iztapalapa. AP. 55-535. D.F., México; slcr@xanum.uam.mx.
Edmundo García-Moya: Posgrado en Botánica, Colegio de Postgraduados, C.P. 56230. Montecillo, Texcoco, Estado de México, México; edmundo@colpos.mx.
Mario Luna-Cavazos:Posgrado en Botánica, Colegio de Postgraduados, C.P. 56230. Montecillo, Texcoco, Estado de México, México; mluna@colpos.mx.
Angélica Romero-Manzanares:Posgrado en Botánica, Colegio de Postgraduados, C.P. 56230. Montecillo, Texcoco, Estado de México, México; dahly@colpos.mx
Noé Manuel Montaño: Departamento de Biología, División de Ciencias Biológicas y de la Salud, Universidad Autónoma Metropolitana, Unidad Iztapalapa. AP. 55-535. D.F., México; nmma@xanum.uam.mx
1. Posgrado en Botánica, Colegio de Postgraduados, C.P. 56230. Montecillo, Texcoco, Estado de México, México; rosalbagarcia@colpos.mx, edmundo@colpos.mx, mluna@colpos.mx, dahly@colpos.mx
2. Unidad de Investigación en Ecología Vegetal, Facultad de Estudios Superiores Zaragoza (FES Zaragoza), Universidad Nacional Autónoma de México. AP. 09230, D.F., México.
3. Departamento de Biología, División de Ciencias Biológicas y de la Salud, Universidad Autónoma Metropolitana, Unidad Iztapalapa. AP. 55-535. D.F., México; slcr@xanum.uam.mx, nmma@xanum.uam.mx
Received 10-III-2011. Corrected 10-VIII-2011. Accepted 13-IX-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}