










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.4 San José Dec. 2011
Ciclos biogeoquímicos en bosques naturales y plantaciones de coníferas en ecosistemas de alta montaña de Colombia
Juan Diego León1, María Isabel González2 & Juan Fernando Gallardo3
1. Universidad Nacional de Colombia, Sede Medellín, Facultad de Ciencias Agropecuarias, Medellín, Colombia, Calle 59A 63-20, 14-330; jdleon@unal.edu.co2. Área de Edafología y Química Agrícola, Universidad de Salamanca, Salamanca 37080 (España); mimg@usal.es
3. C.S.I.C., IRNASA, Salamanca 37071 (España); juanf.gallardo@irnasa.csic.es
Dirección de correspondencia
Abstract
Biogeochemical cycles in natural forest and conifer plantations in the high mountains of Colombia. Plant litter production and decomposition are two important processes in forest ecosystems, since they provide the main organic matter input to soil and regulate nutrient cycling. With the aim to study these processes, litterfall, standing litter and nutrient return were studied for three years in an oak forest (Quercus humboldtii), pine (Pinus patula) and cypress (Cupressus lusitanica) plantations, located in highlands of the Central Cordillera of Colombia. Evaluation methods included: fine litter collection at fortnightly intervals using litter traps; the litter layer samples at the end of each sampling year and chemical analyses of both litterfall and standing litter. Fine litter fall observed was similar in oak forest (7.5Mg ha/y) and in pine (7.8Mg ha/y), but very low in cypress (3.5Mg ha/y). Litter standing was 1.76, 1.73 and 1.3Mg ha/y in oak, pine and cypress, respectively. The mean residence time of the standing litter was of 3.3 years for cypress, 2.1 years for pine and 1.8 years for oak forests. In contrast, the total amount of retained elements (N, P, S, Ca, Mg, K, Cu, Fe, Mn and Zn) in the standing litter was higher in pine (115kg/ha), followed by oak (78kg/ha) and cypress (24kg/ha). Oak forests showed the lowest mean residence time of nutrients and the highest nutrients return to the soil as a consequence of a faster decomposition. Thus, a higher nutrient supply to soils from oaks than from tree plantations, seems to be an ecological advantage for recovering and maintaining the main ecosystem functioning features, which needs to be taken into account in restoration programs in this highly degraded Andean mountains. Rev. Biol. Trop. 59 (4): 1883-1894. Epub 2011 December 01.
Key words: tropical montane forests, litter production, standing litter, nutrient return, Quercus humboldtii.
Resumen
La caída y descomposición de hojarasca representan los principales ingresos de materia orgánica y nutrientes a los ecosistemas forestales. Se estudiaron la caída y acumulación de hojarasca fina y el retorno de nutrientes, en un robledal (Quercus humboldtii) y en plantaciones de pino (Pinus patula) y ciprés (Cupressus lusitanica) en tierras altas de Colombia. La caída de hojarasca fina fue similar entre el robledal (7.5Mg/ha.a) y el pinar (7.8Mg/ha.a), y muy inferior en el cipresal (3.5Mg/ha.a). El mantillo representó 1.76, 1.73 y 1.3Mg/ha.a en el robledal, pinar y cipresal, respectivamente. El tiempo medio de residencia (TMR) del mantillo siguió la secuencia: cipresal (3.3 años)>pinar (2.1 años)>robledal (1.8 años). La cantidad de nutrientes retenidos en el mantillo siguió la secuencia: pinar (115kg/ ha)>robledal (78kg/ha)>cipresal (24kg/ha). Los menores TMR de nutrientes se presentaron para la hojarasca foliar del robledal, en su mayoría inferiores a 1.0 años. En términos de la función ecosistémica en el robledal los procesos estudiados son muy superiores vía provisión de nutrientes al suelo y regulación de los ciclos biogeoquímicos, aspectos que deben ser considerados previa implementación de programas de repoblamiento forestal.
Palabras clave: bosques montanos tropicales, producción de hojarasca, acumulación de hojarasca, retorno de nutrientes, Quercus humboldtii.
La producción y velocidad de descomposición de los residuos orgánicos aportados por el dosel del bosque condicionan el espesor del mantillo u hojarasca acumulada sobre el suelo. Esta capa orgánica regula la mayoría de procesos funcionales en los ecosistemas forestales (Wilcke et al. 2002), protegiendo el suelo de la erosión y favoreciendo la infiltración (Roig et al. 2005). El papel del mantillo en la nutrición vegetal depende fundamentalmente de la capacidad de almacenamiento de nutrientes y del tiempo de retorno de éstos al suelo (Weerakkody & Parkinson 2006). Además esta capa representa la materia prima para la formación de humus y también para el suministro de C y N al suelo (Gallardo et al. 1998). En los bosques montanos tropicales los nutrientes liberados del mantillo representan la principal fuente de suministro y de mantenimiento de la fertilidad del suelo (Vitousek et al. 1995, Berg & Meentemeyer 2001, Parzych & Trojanowski 2006), que satisfacen las necesidades nutricionales de las plantas incluso directamente a través de la densa estera de raíces finas que se entremezclan en el interior del mantillo (García-Oliva et al. 2003, Barreto & León 2005).
En Colombia la deforestación de vastas extensiones de bosques altoandinos (Etter et al. 2006) ha conllevado la alteración no sólo de su alta diversidad y endemismo (Gentry & Dodson 1987, Henderson et al. 1991), sino también de procesos de valioso significado ecosistémico relacionados con el papel del mantillo forestal, como el reciclaje de nutrientes y la regulación de caudales de las cuencas hidrográficas. En las décadas de 1950 y 1960 algunos de estos bosques fueron reemplazados por plantaciones forestales de coníferas con la finalidad de controlar la sedimentación de los proyectos hidroeléctricos (León et al. 2010). Además de este papel se han señalado otros efectos benéficos de las plantaciones, como el incremento de biomasa y de aportes orgánicos al suelo (Sayer & Elliot 2005). Por el contrario se han destacado como efectos negativos de las plantaciones forestales con especies exóticas los relacionados con los ciclos biogeoquímico e hidrológico, tales como las acumulaciones significativas de mantillo (y, con ellas, inmovilización de nutrientes y pérdida de la fertilidad edáfica) y alteración de patrones de penetración del frente de humedad (Sawyer 1993, Lugo 1997, Dames et al. 1998, 2002). No obstante, la información disponible acerca de la influencia de diferentes tipos de cobertura vegetal (nativas e implantadas) sobre las propiedades del suelo y su relación con la función del ecosistema es insuficiente, por ejemplo, en lo que respecta a factores tales como la productividad o la descomposición orgánica (Palm et al. 2007).
El objetivo general de este trabajo fue determinar algunas de las alteraciones más significativas que se producen sobre procesos ecosistémicos clave, como la caída y descomposición de hojarasca del dosel y la liberación de nutrientes desde el mantillo hacia el suelo, como consecuencia del reemplazo de bosques nativos altoandinos por plantaciones forestales de coníferas. Para ello se estudió el ciclo de nutrientes (vía caída y acumulación de hojarasca que conforman el mantillo) en ambos tipos de bosques en tierras altas del centro de Antioquia.
Nuestra hipótesis central fue que los diferentes procesos del ciclo de nutrientes relacionados con la necromasa se alteran al instalar plantaciones de coníferas, implicando efectos negativos en el suministro de nutrientes al suelo y su recirculación en el ecosistema. De aquí que se pretenda comprobar que: i) los retornos de masa seca y bioelementos a través de la hojarasca fina son menores en las plantaciones de coníferas que en el bosque nativo; y ii) las mayores tasas de descomposición del mantillo y liberación de nutrientes al suelo se producen en el bosque nativo.
Materiales y métodos
Muestreo de la caída de hojarasca: Se instalaron 20 trampas circulares de tela fina por bosque (área individual de 0.5m2) a una altura de 1m de la superficie del suelo (Ramírez et al. 2007). La hojarasca fina se recolectó quincenalmente por rodal, se combinó y se separaron las siguientes fracciones: (a) hojas de la especie dominante (HED); (b) hojas de otras especies (HOE); (c) material leñoso (ML), mayormente ramitas <2cm, piñas y trozos de corteza; (d) material no identificado (MNI), esto es, órganos vegetales no reconocibles o diferentes a las categorías anteriores.
Muestreo del mantillo: Al final de cada año se recogieron 15 muestras de 20x20cm2 del mantillo de cada bosque y se separaron las fracciones siguientes: Hojas (H), material leñoso (ML), raíces (R) y otros restos (OR). Únicamente en el robledal fue posible separar las hojas del mantillo según su grado de fragmentación, a saber, hojas enteras (He), hojas fragmentadas (Hf) y hojas muy fragmentadas (Hmf).
Descomposición de la hojarasca y tiempo medio de residencia del mantillo: Para determinar el ritmo de descomposición de la hojarasca se utilizó un índice de mineralización (suponiendo que existe un sistema de equilibro). La constante de descomposición (ko) de la hojarasca se determinó según la Ecuación 1 (Olson 1963, Hernández et al. 1992):
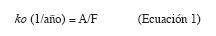
Siendo A la producción anual de hojarasca (Mg/ha.a) y F la cantidad de mantillo sobre el suelo forestal (Mg/ha.a) justo antes de la caída de nuevo material. Valores ko>1 indican un tiempo medio de residencia (TMR) inferior a un año (Scott et al. 1992). El TMR (años) de la hojarasca y de los nutrientes se puede obtener mediante el inverso de ko:
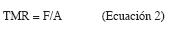
Se calculó además la constante de descomposición kj de Jenny et al. (1949), que supone que en los bosques lluviosos tropicales la caída de hojas se produce permanentemente, aceptando la existencia de un equilibrio en pequeños intervalos de tiempo, oscilando kj entre 0 y 1:
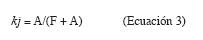
Retorno, acumulación y liberación de nutrientes: La tasa de retorno potencial (TRP) de nutrientes, vía hojarasca foliar del dosel, se calculó multiplicando la materia seca de dicha fracción por la concentración de cada nutriente en ella. La retención de nutrientes por la hojarasca foliar del mantillo (RNM) se obtuvo mediante el producto entre la materia seca de la fracción foliar del mantillo y la concentración del nutriente. La tasa de retorno real de cada nutriente (TRR) se calculó multiplicando la TRP por la constante de descomposición kj respectiva. Finalmente se obtuvo para cada nutriente un índice de eficacia del retorno (IER) mediante el cociente entre la TRP y la reserva o cantidad del nutriente almacenada en la hojarasca foliar del mantillo (RNM). Valores IER cercanos a uno indican que la cantidad que llega anualmente de cada nutriente al suelo vía hojarasca es próxima a la que se acumula de éste en el suelo, mientras que valores inferiores a uno indican que hay una acumulación biológica del nutriente en el mantillo.
Determinaciones químicas: Las muestras quincenales de hojarasca fina se integraron por períodos de dos meses para cada fracción. Las muestras de mantillo se integraron cada año por fracción y, adicionalmente, según el grado de fragmentación para las hojas de roble. Las muestras se secaron a 65°C, se combinaron, homogenizaron, molieron y calcinaron a 550ºC en mufla. Las cenizas se disolvieron en ácido nítrico, se diluyeron y se analizaron el Ca, Mg, K y elementos traza (Cu, Fe, Mn, Zn) usando espectrofotometría de absorción atómica. El P total se determinó por espectrofotometría UVVIS. Las determinaciones de C, N y S totales se realizaron mediante combustión seca en un Analizador LECO CNS-2000.
Se aplicó un análisis de varianza ANOVA, previa comprobación de la homogeneidad de la varianza (prueba de Bartlett). Las diferencias entre los valores experimentales se determinaron mediante la prueba de comparación de medias de Tukey (p<0.05).
Resultados
Producción de hojarasca fina: La producción anual de hojarasca fina del cipresal fue muy baja, cerca de la mitad de las del robledal y del pinar, y ambas similares entre sí (Cuadro 3). Del total de hojarasca fina, la fracción foliar (HED+HOE) representó 71, 63 y 71% en el robledal, pinar y cipresal, respectivamente.
Acumulación de mantillo: El total de mantillo acumulado en el robledal fue muy similar al del pinar (17.6 y 17.3Mg/ha respectivamente; Cuadro 4). La mayor acumulación de hojarasca foliar del pinar (6.6Mg/ha) representó un 38% del total del mantillo. El material leñoso representó en el robledal y el pinar el 23 y 28%, respectivamente, mientras en el cipresal fue sólo de 4%. El mantillo del robledal mostró una alta ocupación por raíces finas (22%).
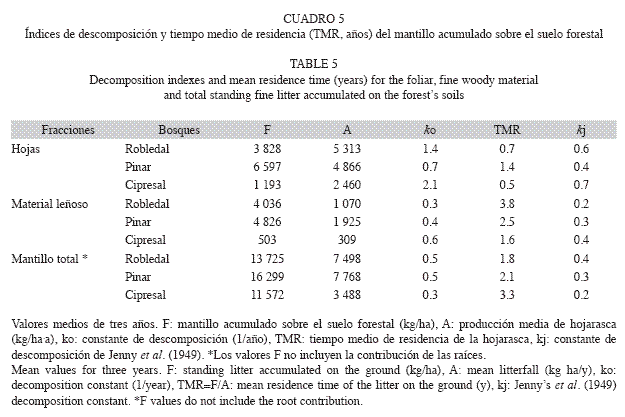
Descomposición de la hojarasca y tiempo medio de residencia del mantillo: Los índices de descomposición (Cuadro 5) mostraron un comportamiento similar, aunque kj aumentó más lentamente, dado su límite superior 1. Los mayores valores de los índices para la hojarasca foliar fueron los del cipresal (ko=2.1, kj=0.7), con un TMR=0.5 años, seguidos por el robledal y el pinar; este patrón se mantuvo también en los restos leñosos (ML). Al considerar todo el mantillo (sin raíces) la descomposición fue superior en el robledal (ko=0.5, kj=0.4) con un TMR=1.8 años.
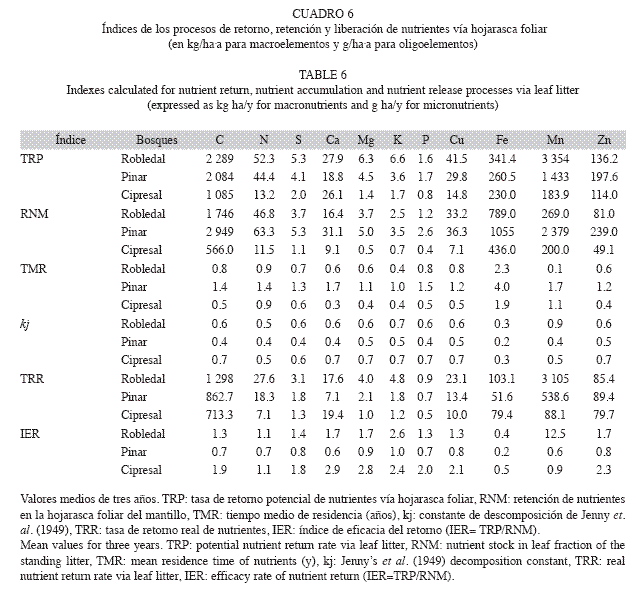
Retorno, acumulación y liberación de nutrientes: Las mayores cantidades de nutrientes retornados a través de la hojarasca foliar (TRP) se presentaron en el robledal (Cuadro 6), representando el retorno de carbono (C) 2.3Mg/ha.a. Los macronutrientes retornados mayoritariamente a partir del dosel (vía hojarasca foliar) fueron N y Ca. Por el contrario los retornos de los macronutrientes Mg, S y K fueron menores, registrándose los más bajos valores en el cipresal y mínimos los de P. Los retornos potenciales de oligoelementos siguieron el orden: Mn>Fe>Zn>Cu. La mayor acumulación de nutrientes en la hojarasca foliar del mantillo (RNM) correspondió al pinar, con un valor medio de 111kg/ha de macronutrientes, 4kg/ha de micronutrientes y aproximadamente 3Mg/ha de C. Los menores valores kj y, por ende, los mayores TMR se encontraron en el pinar. En éste debe transcurrir entre 1.0 y 1.7 años para la liberación efectiva de los macronutrientes, al contrario que en el cipresal, donde dicho período fue de 0.3-0.9 años. Para los oligoelementos el mayor TMR fue el de Fe (1.9-4.0 años). Al igual que para la TRP los mayores valores de TRR se encontraron en el robledal. La mayor TRR se determinó en N para el robledal (28kg/ha.a) y pinar (18kg/ha.a) y para Ca en el cipresal (19kg/ha.a). Los retornos reales de C por descomposición de la hojarasca foliar siguieron la secuencia robledal (1.3Mg/ ha)>pinar (0.9Mg/ha)>cipresal (0.7Mg/ha). La mayor TRR de oligoelementos se observó para Mn y la menor para Cu. La eficacia del retorno (IER) indicó que en el cipresal retornaron más rápidamente los nutrientes al suelo; se observaron allí los máximos absolutos del IER para Ca (2.9) y Mg (2.8). El máximo absoluto del IER en el robledal se obtuvo para Mn (13.0) y en el pinar para K (1.0).
Discusión
Producción de hojarasca fina: No se registraron diferencias significativas interanuales en la producción de hojarasca foliar ni de hojarasca fina total entre bosques. La producción media de hojarasca fina total del robledal (7.5Mg/ha.a) corresponde con la de otros bosques montanos tropicales (7.0-8.0Mg/ha.a: Veneklaas 1991, Nadkarni & Matelson 1992, Vitousek et al. 1995, McDonald & Healey 2000, Kitayama & Aiba 2002, Weerakkody & Parkinson 2006). La producción del pinar fue similar a la de otras plantaciones tropicales (Lundgren 1978, Cuevas & Lugo 1998), mientras la del cipresal fue cercana a la de bosques de coníferas de regiones templadas (Dames et al. 1998, Santa Regina & Tarazona 2001). El aporte de la fracción foliar total fue la de mayor peso en la hojarasca fina, siendo muy semejante a la indicada por Meentemeyer et al. (1982), quienes sugirieron una participación de esta fracción en los trópicos inferior al 70%.
Acumulación de mantillo: El mantillo acumulado fue muy semejante en el robledal y el pinar (17Mg/ha) y cercano al límite superior de otros reportes (10.1-16.5Mg/ha: Lugo et al. 1990, Cuevas et al. 1991, Nadkarni & Matelson 1992, Smith et al. 1998, Sundarapandian & Swamy 1999, Weerakkody & Parkinson 2006). La acumulación de hojas en el pinar fue notoriamente superior a la del robledal, aunque ambos mostraron una producción similar de hojarasca y una acumulación de mantillo total sin diferencias significativas entre ambos bosques, denotando una menor velocidad de descomposición de las hojas de pino. Aun cuando la biomasa y el área basal del pinar son superiores a los del robledal, la similar producción de hojarasca aérea refleja el carácter caducifolio de las especies nativas presentes en el segundo. En el robledal se evidenció una fuerte fragmentación de la hojarasca foliar, dada la superioridad de las fracciones Hf y Hmf sobre la fracción He. La acumulación de hojas en el mantillo del robledal (3.8Mg/ ha) concuerda con las de otros estudios de la región tropical (1.8 a 6.8Mg/ha: Scott et al. 1992, Sundarapandian & Swamy 1999, McDonald & Healey 2000). La escasa participación de la fracción foliar en el mantillo del cipresal (1.2Mg/ha) sugirió su rápida descomposición. Sin embargo, estudios realizados en la zona de trabajo (Ruiz et al. 2005 a,b, León et al. 2010) constataron la ocurrencia en el cipresal de flujos significativos de escorrentía superficial (superiores al 10% de la lluvia), promovidos principalmente por el desarrollo de procesos de repelencia al agua (Duque et al. 2004) que causan arrastre y exportan hojarasca depositada sobre el piso forestal, lo que reduce el espesor del mantillo.
Descomposición de la hojarasca y tiempo medio de residencia del mantillo: La baja participación de hojas en el mantillo del robledal (28% sin considerar raíces) con respecto a la de la producción aérea (71%), indicó su fragmentación hacia la fracción OR y una rápida descomposición (kj=0.6, ko=1.4). Los menores TMR para el mantillo total del robledal (1.8 años) resultan probablemente del aporte de la fracción OR (33% del total), finos fragmentos de materiales desconocidos que tienden a disponerse en la parte más profunda del mantillo por el arrastre pluvial, lo que facilita su descomposición. Además, el mayor carácter lábil de las hojas de roble (C/N=39) con respecto a las de pino (C/N=51) y ciprés (C/N=56) acelera la descomposición (Torreta & Takeda 1999, Castellanos-Barliza & León 2011). En las dos últimas especies la mayor abundancia de sustancias polifenólicas podría inhibir el crecimiento microbiano y frenar la actividad descomponedora (Egunjobi & Onweluzo 1979). Así, el paso en el pinar de una participación del 63% de la fracción foliar en la hojarasca desprendida del dosel al 40% en el mantillo sugiere mayor bio-resistencia de las acículas (kj=0.4, ko=0.7), esto es, una mineralización más lenta y una mayor acumulación sobre el suelo. En el cipresal el paso de una participación del 71% de hojas en la producción aérea a sólo 10% en el mantillo, indicaría que las hojas de ciprés son las más lábiles de las tres especies (kj=0.7, ko=2.1), lo cual es opuesto a los hallazgos de otros estudios (Lisanework & Michelsen 1994) y con la relación C/N aquí determinada. Como se indicó tal situación no refleja una intensa descomposición, sino una pérdida de mantillo arrastrado por la escorrentía superficial que determina el excesivamente bajo TMR de la hojarasca foliar (0.5 años). Los patrones de producción y posterior acumulación de los restos leñosos (ML) mostraron cierta semejanza a los de la fracción foliar, en el cipresal la situación es similar a la de la hojarasca foliar en cuanto a la sobreestimación de su descomposición.
Los valores ko de la hojarasca foliar del robledal fueron inferiores a los de bosques tropicales de tierras bajas (1.4-2.0: Anderson et al. 1983, Smith et al. 1998) y de tierras altas (2.0: Burghouts et al. 1998). De manera similar el valor ko del mantillo del robledal (0.5) se localizó en el límite inferior del intervalo correspondiente a otros bosques montanos tropicales (0.4-1.2: Nadkarni & Matelson 1992, Kitayama & Aiba 2002, Weerakkody & Parkinson 2006). Consecuentemente el TMR del mantillo del robledal (1.8 años) fue mayor que los obtenidos para el subhorizonte Oi en otros bosques montanos tropicales (Heaney & Proctor 1989, Nadkarni & Matelson 1992, Bruijnzeel et al. 1993, Kitayama & Aiba 2002, Wilcke et al. 2002, Weerakkody & Parkinson 2006). Los valores ko de la hojarasca foliar (0.7) y del mantillo (0.5) del pinar fueron inferiores a los reportados para otras especies de pino de tierras bajas tropicales (Lugo et al. 1990, Cuevas et al. 1991, Smith et al. 1998), donde las condiciones climáticas favorecen la descomposición.
Retorno, acumulación y liberación de nutrientes: La TRP de N vía hojarasca foliar del robledal coincidió con la de otros bosques montanos tropicales (22.1-96.7kg/ha.a:
Lundgren 1978, Fassbender & Grimm 1981, Veneklaas 1991, Herbohn & Congdon 1993, Vitousek et al. 1995, Liu et al. 2002). Por el contrario la TRP de P se localizó cerca al límite inferior de tales estudios (1.1-5.2kg/ha.a) mostrando un mayor carácter limitante. A pesar de la baja producción de hojarasca foliar del cipresal, la TRP de Ca (26.0kg/ha.a) fue cercana a la del robledal (28.0kg/ha.a), dada su elevada concentración foliar (roble: 4.5mg/g, ciprés: 7.6mg/g). Las mayores TRP de oligoelementos se presentaron para Mn y Fe, siendo mayores los del primero a excepción del cipresal, donde su bajo valor (0.18kg kg/ha.a) se atribuye a un mayor pH edáfico. La TRP de Mn en el robledal fue muy superior a las de otros bosques montanos tropicales (1.8-4.7kg/ha.a: Fassbender & Grimm 1981, Wilcke et al. 2002).
De la comparación entre las cantidades de nutrientes foliares retenidas en el mantillo (RNM) y las aportadas por la hojarasca (TRP) el retorno de nutrientes al suelo se produjo con mayor celeridad en el cipresal y en menor medida en el robledal. No obstante, la exportación del mantillo por el flujo de escorrentía superficial en el cipresal condujo a la subvaloración del TMR de los diferentes nutrientes en este bosque. De hecho, ha sido ampliamente aceptado como parámetro para predecir la velocidad de descomposición de la hojarasca foliar el coeficiente C/N (Taylor et al. 1989, Aceñolaza & Gallardo 1999, Arunachalam & Singh 2002), cuyo valor para el ciprés fue el mayor encontrado para las tres especies, con lo cual se esperaría una más lenta descomposición. En concordancia con esto, Aerts & Heil (1993) reportaron la relación C/N como buena predictora de la descomposición de la hojarasca especialmente en sustratos con bajo contenido de lignina; además, Prause & Fernández-López (2007) indicaron que la concentración foliar inicial de N puede ser empleada como un indicador de la velocidad de descomposición para bosques subtropicales. El valor de la relación C/N para ambas especies de coníferas fue incluso superior a los considerados críticos (30-40: Torreta & Takeda 1999), mientras que el mínimo absoluto C/N en el robledal indicó una mayor mineralización y, con ello, una más rápida liberación de N y otros elementos, expresadas en menores TMR para la mayoría de ellos. La baja TRR de P en los tres bosques, sugirió su deficiencia para la vegetación. Los relativamente altos valores de retorno real (TRR) del robledal (especialmente de N) parecieran indicar la inexistencia de limitaciones nutricionales (con excepción del P), situación contraria a la del cipresal. De las tres especies el roble mostró la mejor adaptación a la acidez, dada la fuerte liberación de Mn. Los valores del IER obtenidos señalaron que en el pinar domina un patrón de bio-acumulación de nutrientes en la necromasa, corroborando que es la especie con más limitaciones nutricionales. Por el contrario en el robledal la cantidad de nutrientes que anualmente llega por hojarasca sobrepasa en mucho la cantidad que queda bio-acumulada, lo que puede indicar abundancia nutricional. Aunque esta última situación se determinó aún con mayor fuerza en el cipresal, como ya se ha señalado las pérdidas de mantillo por escorrentía superficial son las que producen tales resultados.
En síntesis, se pudo constatar que en los sitios altoandinos estudiados los bosques nativos de roble producen una mayor cantidad de hojarasca fina que las plantaciones forestales de coníferas, suministrando con ella una fuente importante de materia orgánica y C para el suelo. Asimismo se verifican retornos (potencial y real) de elementos muy superiores en el robledal que favorecen el reciclaje de nutrientes y la actividad biológica del suelo. Así, desde la perspectiva de los servicios ecosistémicos que el suelo proporciona, se puede afirmar que los robledales estudiados son muy superiores a las plantaciones de coníferas en términos de la provisión de nutrientes y de la regulación de los ciclos biogeoquímicos. Esto se produce seguramente como resultado del balance biogeoquímico que se ha establecido durante largos periodos de tiempo entre las especies nativas de los robledales climácicos y la comunidad microbiológica descomponedora del suelo, configurando un modelo de reciclaje de nutrientes co-adaptado. Los resultados de este estudio señalan que existen grandes diferencias en la función ecosistémica en cuanto a los procesos relacionados con el ciclo biogeoquímico entre las plantaciones forestales y los bosques nativos altoandinos de roble, lo cual debe ser considerado como base para la selección de uno u otro tipo de cobertura vegetal de acuerdo con los objetivos de programas de restauración o repoblación forestal.
Agradecimientos
Al personal del Área de Biogeoquímica del Laboratorio de Ecología César Pérez Figueroa y de la Estación Forestal Piedras Blancas de la Universidad Nacional de Colombia, Sede Medellín. A Empresas Públicas de Medellín por el acceso a los predios e información meteorológica. Se hace un agradecimiento especial a C.M. Zapata y J.A. Ramírez por su participación en la instalación y monitoreo de los ensayos. Este trabajo fue financiado por la Dirección de Investigaciones de la Universidad Nacional de Colombia.
Referencias
Aceñolaza, P.G. & J.F. Gallardo. 1999. Leaf decomposition and nutrient release in montane forests of Northwestern Argentina. J. Trop. Forest. Sci. 11: 619-630. [ Links ]
Aerts, R. & G.W. Heil. 1993. Heathlands: patterns and processes in a changing environment. Kluwer, Dordrecht, Holanda. [ Links ]
Anderson, J.M., J. Proctor & H.W. Vallack. 1983. Ecological studies in four contrasting lowland rain forest in Gunung Mulu National Park, Sarawak. III. Decomposition processes and nutrient losses from leaf litter. J. Ecol. 71: 503-527. [ Links ]
Arunachalam, A. & N.D. Singh. 2002. Leaf litter decomposition of evergreen and deciduous Dillenia species in humid tropics of north-east India. J. Trop. Forest. Sci. 14: 105-115. [ Links ]
Barreto, L.H. & J.D. León. 2005. Masa total y contenido de nutrientes en raíces finas de ecosistemas forestales (Pinus patula Schltdl y Cham., Cupressus lusitánica Mill. y Quercus humboldtii Bonpl.) de Piedras Blancas, Antioquia-Colombia. Rev. Fac. Nal. Agr. Medellín 58: 2907-2929. [ Links ]
Berg, B. & V. Meentemeyer. 2001. Litterfall in Europian pine and spruce forests as related to climate. Can. J. Forest. Res. 31: 292-301. [ Links ]
Bruijnzeel, L.A., M.J. Waterloo, J. Proctor, A.T. Kuiters & B. Kotterink. 1993. Hydrological observations in montane rain forest on Gunung Silam, Sabah, Malaysia, with special reference to the "Massenerhebung" effect. J. Ecol. 81: 145-167. [ Links ]
Burghouts, T.B.A., N.M. Van Stralen & L.A. Bruijnzeel. 1998. Spatial heterogeneity of element and litter turnover in a Bornean forest. J. Trop. Ecol. 14: 477-506. [ Links ]
Castellanos-Barliza, J. & J.D. León. 2011. Descomposición de hojarasca y liberación de nutrientes en plantaciones de Acacia mangium (Mimosaceae) establecidas en suelos degradados de Colombia. Rev. Biol. Trop. 59: 113-128. [ Links ]
Cuevas, E., S. Brown & A.E. Lugo. 1991. Above- and belowground organic matter storage and production in a tropical pine plantation and a paired broadleaf secondary forest. Plant. Soil. 135: 257-268. [ Links ]
Cuevas, E. & A. Lugo. 1998. Dynamics of organic matter and nutrient return from litterfall in stands of ten tropical tree plantation species. For. Ecol. Manag. 112: 263-279. [ Links ]
Dames, J.F., M.C. Scholes & C.J. Straker. 1998. Litter production and accumulation in Pinus patula Schiede plantations of the Mpumalanga Province, South Africa. Plant. Soil 203: 183-190. [ Links ]
Dames, J.F., M.C. Scholes & C.J. Straker. 2002. Nutrient cycling in a Pinus patula plantation in the Mpumalanga Province, South Africa. Appl. Soil. Ecol. 20: 211-226. [ Links ]
Egunjobi, J.K. & B.S. Onweluzo. 1979. Litter fall, mineral turnover and litter accumulation in Pinus caribaea L. stands at Ibadan, Nigeria. Biotropica 11: 251-255. [ Links ]
Etter, A., C. McAlpine, K. Wilson, S. Phinn & H. Possingham. 2006. Regional patterns of agricultural land use and deforestation in Colombia. Agr. Ecosyst. Environ. 114: 369-386. [ Links ]
Fassbender, H.W. & V. Grimm. 1981. Ciclos biogeoquímicos en un ecosistema forestal de los Andes Occidentales de Venezuela. II. Producción y descomposición de los residuos vegetales. Turrialba 31: 39-47. [ Links ]
Galindo, R., J. Betancur & J.J. Cadena. 2003. Estructura y composición florística de cuatro bosques andinos del santuario de flora y fauna Guanentá-Alto río Fonce, cordillera oriental colombiana. Caldasia 25: 313-335. [ Links ]
Gallardo, J.F., A. Martín & I. Santa Regina. 1998. Nutrient cycling in deciduous forest ecosystems of the ‘Sierra de Gata’ mountains: Aboveground litter production and potential return. Ann. Forest. Sci. 55: 749-769. [ Links ]
García-Oliva, F., B. Sveshtarova & M. Oliva. 2003. Seasonal effects on soil organic carbon dynamic in a tropical decidous forest ecosystem in western Mexico. J. Trop. Ecol. 19: 179-188. [ Links ]
Gentry, A.H. & D. Dodson. 1987. Contribution of non trees to species richness of a tropical rain forest. Biotropica 19: 149-156. [ Links ]
Heaney, A. & J. Proctor. 1989. Chemical elements in litter in forests on Volcán Barva, Costa Rica, p. 255-271. In J. Proctor (ed.). Mineral nutrients in tropical forest and savanna ecosystems. Blackwell, Oxford, Inglaterra. [ Links ]
Henderson, A., S.P. Churchill & J.L. Luteyn. 1991. Neotropical plant diversity. Nature 351: 21-22. [ Links ]
Herbohn, J.L. & R.A. Congdon. 1993. Ecosystem dynamics at disturbed and undisturbed sites in north Queensland wet tropical rain forest. II. Litterfall. J. Trop. Ecol. 9: 365-380. [ Links ]
Hernández, I.M., I. Santa Regina & J.F. Gallardo. 1992. Dinámica de la descomposición de la hojarasca forestal en bosques de la Cuenca del Duero: Modelización de la pérdida de peso. Arid. Soil Res. Rehabil. 6: 339-355. [ Links ]
Jenny, H., S.P. Gessel & F.T. Bingham. 1949. Comparative study of decomposition of organic matter in temperate and tropical regions. Soil Sci 68: 419-432. [ Links ]
Kitayama, K. & S. Aiba. 2002. Ecosystem structure and productivity of tropical rainforests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu, Borneo. J. Ecol. 90: 37-51. [ Links ]
León, J.D., G. Vélez & A.P. Yepes. 2009. Estructura y composición florística de tres robledales en la región norte de la cordillera central de Colombia. Rev. Biol. Trop. 57: 1165-1182. [ Links ]
León, J.D., M.I. González & J.F. Gallardo. 2010. Distribución del agua lluvia en tres bosques altoandinos de la Cordillera Central de Antioquia, Colombia. Rev. Fac. Nal. Agr. Medellín 63: 5319-5336. [ Links ]
Lisanework, N. & A. Michelsen. 1994. Litterfall and nutrient release by decomposition in three plantations compared with a natural forest in the Ethiopian highland. For. Ecol. Manag. 65: 149-164. [ Links ]
Liu, W., E.D. Fox & Z. Xu. 2002. Biomass and nutrient accumulation in montane evergreen broad-leaved forest (Lithocarpus xylocarpus type) in Ailao mountains, SW China. For. Ecol. Manag. 158: 223-235. [ Links ]
Lugo, A.E. 1997. The apparent paradox of reestablishing species richness on degraded lands with monocultures. For. Ecol. Manag. 99: 9-19. [ Links ]
Lugo, A.E., E. Cuevas & M.J. Sánchez. 1990. Nutrients and mass in litter and top soil of ten tropical tree plantations. Plant. Soil 125: 263-280. [ Links ]
Lundgren, B. 1978. Soil conditions and nutrient cycling under natural and plantation forests in Tanzanian Highlands. Reports in forest ecology and forest soils 31. Swedish University of Agricultural Sciences, Upsala, Suecia. [ Links ]
McDonald, M.A. & J.R. Healey. 2000. Nutrient cycling in secondary forests in the Blue Mountains of Jamaica. For. Ecol. Manag. 139: 257-278. [ Links ]
Meentemeyer, V., E.O. Box & R. Thompson. 1982. World patterns and amounts of terrestrial plant litter production. Bioscience 32: 125-128. [ Links ]
Nadkarni, N.M. & T.J. Matelson. 1992. Biomass and nutrient dynamics of fine litter of terrestrially rooted material in a neotropical montane forest, Costa Rica. Biotropica 24: 113-120. [ Links ]
Olson, J.S. 1963. Energy storage and the balance of producers and decomposers in ecologycal systems. Ecology 44: 322-331. [ Links ]
Parzych, A. & J. Trojanowski. 2006. Precipitation and duff fall as natural sources of nitrogen and phosphorus for forest soils in the Słowiński National Park. Baltic Coastal Zone 10: 47-59. [ Links ]
Prause, J. & C. Fernández-López. 2007. Litter decomposition and lignin/cellulose and lignin/total nitrogen rates of leaves in four species of the Argentine Subtropical Forest. Agrochimica 51: 294-300. [ Links ]
Ramírez, J.A., C.M. Zapata, J.D. León & M.I. González. 2007. Caída de hojarasca y retorno de nutrientes en bosques montanos andinos de Piedras Blancas, Antioquia, Colombia. Interciencia 32: 303-311. [ Links ]
Roig, S., M. Del Río, I. Cañellas & G. Montero. 2005. Litter fall in Mediterranean Pinus pinaster Ait. Stands under different thinning regimes. For. Ecol. Manag. 206: 179-190. [ Links ]
Ruiz, O.A., J.J. Acosta & J.D. León. 2005a. Escorrentía superficial en bosques montanos naturales y plantados de Piedras Blancas, Antioquia (Colombia). Rev. Fac. Nal. Agr. Medellín. 58: 2635-2649. [ Links ]
Ruiz, O.A., J.J. Acosta & J.D. León. 2005 b. Flujo de nutrientes en la escorrentía superficial de bosques montanos en Piedras Blancas, Antioquia (Colombia). Meteorología Colombiana 9: 23-29. [ Links ]
Santa Regina, I. & T. Tarazona. 2001. Nutrient cycling in a natural beech forest and adjacent planted pine in northern Spain. Forestry 74: 11-28. [ Links ]
Sawyer, J. 1993. Plantations in the tropics: environmental concerns. IUCN, Gland, Suiza. [ Links ]
Sayer, J. & C. Elliot. 2005. The role of commercial plantations in forest landscape restoration, p. 379-383. In S. Mansourian, D. Vallauri & N. Duddley (eds.). Forest restoration in landscapes. Beyond planting trees. Springer, Nueva York, EEUU. [ Links ]
Scott, D.A., J. Proctor & J. Thompson. 1992. Ecological studies on a lowland evergreen rain forest on Maracá island, Roraima, Brazil. II. Litter and nutrient cycling. J. Ecol. 80: 705-717. [ Links ]
Smith, K., H.L. Gholz & F.D.A. Oliveira. 1998. Litterfall and nitrogen-use efficiency of plantations and primary forest in the eastern Brazilian Amazon. For. Ecol. Manag. 109: 209-220. [ Links ]
Sundarapandian, S.M. & P.S. Swamy. 1999. Litter production and leaf-litter decomposition of selected tree species in a tropical forest at Kodayar in the Western Ghats, India. For. Ecol. Manag. 123: 231-244. [ Links ]
Taylor, B.R., D. Parkinson & W.F.J. Parsons. 1989. Nitrogen and lignin content as predictors of litter decay rates: a microcosm test. Ecology 70: 97-104. [ Links ]
Torreta, N.K. & H. Takeda. 1999. Carbon and nitrogen dynamics of decomposing leaf litter in a tropical hill evergreen forest. Eur. J. Soil. Biol. 35: 57-63. [ Links ]
Veneklaas, E.J. 1991. Litterfall and nutrient fluxes in two montane tropical rain forests, Colombia. J. Trop. Ecol. 7: 319-336. [ Links ]
Vitousek, P.M., G. Gerrish, D.R. Turner, L.R. Walker & D. Mueller-Dombois. 1995. Litterfall and nutrient cycling in four Hawaiian montane rainforests. J. Trop. Ecol 11: 189-203. [ Links ]
Weerakkody, J. & D. Parkinson. 2006. Input, accumulation and turnover of organic matter, nitrogen and phosphorus in surface organic layers of an upper montane rainforest in Sri Lanka. Pedobiologia 50: 377-383. [ Links ]
Wilcke, W., S. Yasin, U. Abramowski, C. Valarezo & W. Zech. 2002. Nutrient storage and turnover in organic layers Andean tropical montane rain forest in Ecuador. Eur. J. Soil. Sci. 53: 15-27. [ Links ]
Correspondencia a: Juan Diego León. Universidad Nacional de Colombia, Sede Medellín, Facultad de Ciencias Agropecuarias, Medellín, Colombia, Calle 59A 63-20, 14-330; jdleon@unal.edu.co
María Isabel González. Área de Edafología y Química Agrícola, Universidad de Salamanca, Salamanca 37080 (España); mimg@usal.es
Juan Fernando Gallardo. C.S.I.C., IRNASA, Salamanca 37071 (España); juanf.gallardo@irnasa.csic.es
Recibido 04-II-2011. Corregido 05-IV-2011. Aceptado 09-V-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}