










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.3 San José Sep. 2011
Insect herbivores associated with Baccharis dracunculifolia (Asteraceae): responses of gall-forming and free-feeding insects to latitudinal variation
Marcílio Fagundes1 & G. Wilson Fernandes2
1. Laboratório de Biologia da Conservação DBG/ CCBS, Universidade Estadual de Montes Claros, Montes Claros, MG, Brazil, 39401-089; marcilio.fagundes@gmail.com
2. Ecologia Evolutiva & Biodiversidade/DBG/ICB/Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil; gwfernandes@gmail.com
Dirección para correspondencia:
1. Laboratório de Biologia da Conservação DBG/ CCBS, Universidade Estadual de Montes Claros, Montes Claros, MG, Brazil, 39401-089; marcilio.fagundes@gmail.com
2. Ecologia Evolutiva & Biodiversidade/DBG/ICB/Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil; gwfernandes@gmail.com
Dirección para correspondencia:
Abstract
The spatial heterogeneity hypothesis has been invoked to explain the increase in species diversity from the poles to the tropics: the tropics may be more diverse because they contain more habitats and microhabitats. In this paper, the spatial heterogeneity hypothesis prediction was tested by evaluating the variation in richness of two guilds of insect herbivores (gall-formers and free-feeders) associated with Baccharis dracunculifolia (Asteraceae) along a latitudinal variation in Brazil. The seventeen populations of B. dracunculifolia selected for insect herbivores sampling were within structurally similar habitats, along the N-S distributional limit of the host plant, near the Brazilian sea coast. Thirty shrubs were surveyed in each host plant population. A total of 8 201 galls and 864 free-feeding insect herbivores belonging to 28 families and 88 species were sampled. The majority of the insects found on B. dracunculifolia were restricted to a specific site rather than having ageographic distribution mirroring that of the host plant. Species richness of free-feeding insects was not affected by latitudinal variation corroborating the spatial heterogeneity hypothesis. Species richness of gall-forming insects was positively correlated with latitude, probably because galling insect associated with Baccharris genus radiated in Southern Brazil. Other diversity indices and evenness estimated for both gall-forming and free feeding insect herbivores, did not change with latitude, suggesting a general structure for different assemblages of herbivores associated with the host plant B. dracunculifolia. Thus it is probable that, insect fauna sample in each site resulted of large scale events, as speciation, migration and coevolution, while at local level, the population of these insects is regulated by ecological forces which operate in the system. Rev. Biol. Trop. 59 (3): 1419-1432. Epub 2011 September 01.
Key words: community organization, latitudinal diversity gradient, spatial heterogeneity hypothesis, species diversity.
Resumen
La hipótesis de heterogeneidad espacial se ha utilizado para explicar el aumento en la diversidad de especies desde los polos a los trópicos: los trópicos pueden ser más diversos ya que están conformados por una mayor cantidad de hábitats y micro-hábitats. En este estudio, la hipótesis de heterogeneidad espacial se puso a prueba evaluando la variación en la riqueza de dos gremios de insectos herbívoros (formadores de agallas y de alimentación libre) asociados con B. dracunculifolia (Asteracea) a lo largo de un gradiente latitudinal en Brasil. Las diecisiete poblaciones de B. dracunculifolia seleccionadas para el muestreo de insectos herbívoros estaban en hábitats con una estructura similar, a lo largo del límite Norte-Sur de distribución de la planta hospedera, cerca de la costa brasileña. De cada población de planta hospedera, se muestrearon treinta arbustos y se obtuvo un total de 8 201 agallas y 864 insectos de alimentación libre pertenecientes a 28 familias y 88 especies. La mayoría de los insectos que se encontraron en B. dracunculifolia estaban restringidos a un sitio específico en lugar de tener una distribución geográfica similar a la de la planta hospedera. La riqueza de especies de insectos de alimentación libre no se vió afectada por el gradiente latitudinal, por lo que se corroboró la hipótesis de heterogeneidad espacial. Mientras que la riqueza de especies de insectos formadores de agallas se correlacionó positivamente con la latitud, probablemente debido a que los insectos asociados al género Baccharis se extendieron hacia el sur de Brasil. Otros índices de diversidad y equidad estimados no variaron con la latitud para ninguno de los dos grupos de insectos herbívoros evaluados, lo que sugirie una estructura general para diferentes conjuntos de herbívoros asociados con la planta hospedera B. dracunculifolia. Por lo tanto, es probable que, la muestra de insectos en cada sitio sea resultado de eventos a gran escala, como la especiación, migración y coevolución; mientras que a nivel local la población de estos insectos está regulada por fuerzas ecológicas que operan dentro del sistema.
Palabras clave: organización de comunidades, gradiente latitudinal de diversidad, hipótesis de heterogeneidad espacial, diversidad de especies.
Latitudinal variations in species diversity are among the most conspicuous and universal patterns in community ecology (Pianka 1966). According Willig et al. (2003), just with notable exceptions, the pattern of higher diversity at low latitudes holds true regardless of the biota’s taxonomic affiliation (e.g., mammals, fishes, arthropods and plants), geographic context (e.g., all continents and oceans) or time domain (recent and 70 Mya). Explanations for higher (Buckley et al. 2010) or habitat heterogeneity (Whitehouse et al. 2009) to explain the increase of diversity towards the tropics.
In this paper the focus is on the spatial heterogeneity hypothesis, and the study objects are different insect herbivore guilds associated with a single host plant. On the local scale, the spatial heterogeneity hypothesis suggests that the tropics are more diverse because they contain more microhabitats (Pianka 1966). On a regional scale, this hypothesis has been proposed to explain the increase of species diversity from the poles to the tropics: the tropics are more diverse because they contain more habitats and microhabitats (Dawidowitz & Rosenzweigh 1998). This greater number of habitats and microhabitats allows taxa to partition the environment more finely, and more species to co-exist in the tropics (Pianka 1966). A positive relationship between habitat complexity and species diversity has been shown in a variety of environments and for a large number of taxa (Otte 1976, Terborgh 1992, Gaston & Williams 1996, Ribeiro et al. 1998, Thomaz et al. 2008). On the other hand, as a corollary, the spatial heterogeneity hypothesis also implies that, within structurally uniform environments, species diversity should not change along a latitudinal gradient.
The most species alive are tropical arthropods associated with plants (Price et al. 1995). Herbivorous arthropods maintain many important ecosystem processes and form sizeable parts of terrestrial food webs (Lewinsohn & Roslin 2008). In fact, in terms of biomass, insects in tropical forests constitute several tons per hectare compared to a few kilograms per hectare for birds and mammals. Moreover, insects in the tropics munch through an estimated 680 kg/ha.y of leaves compared to 100 to 300 kg/ha.y of leaves by vertebrates (Dajoz 2000). Herbivorous insects are composed of various feeding guilds (e.g. free-feeding and galling insects), with different specialization levels on their host plant and habitat (Koricheva et. al. 1998, Novotny & Basset 2005). In fact, comparatively to free-feeding insects, galling insects generally are considered more specialized herbivores (Frenzel & Brandl 1998). Thus, it is probable that, when the insect fauna associated with single host plant is analyzed, free-feeding insects (generally with more oligophagous feeding habit) are more dependent on habitat characteristics, while galling insects respond more finely to specific host plant attributes (Koricheva et al. 1998, Golden & Crist 1999).
Despite the high diversity of insect herbivores in the tropics, few studies have adequately addressed the influence of historical and biogeographical processes on species richness patterns of tropical insect herbivores (Price et al. 1995, Ribeiro et al. 1998, Majer et al. 2001). Moreover, many insect taxa as aphids (Dixon et al. 1987), Ichneumonidae (Sime & Brower 1998) and gall-forming insects (Fernandes & Price 1988) do not fit the general pattern of increasing species richness with decreasing latitude. Evolutionary explanations to these latitudinal diversity anomalies have focused on either variation in rates of diversification or the amount of time available for speciation within a region (Buckley et al. 2010). While some studies include herbivores from several insect orders, studies of more than one guild on the same hosts are lacking from tropical regions (Novotny & Basset 2005). In this study the prediction of spatial heterogeneity hypothesis was tested by evaluating the variation in diversity of two guilds of insect herbivores (galling and free-feeding) along the distributional limits of the host plant, near the Brazilian sea coast. The data were collected within homogeneous habitat and the arthropod sampled was associated with the host plant Baccharis dracunculifolia.
Materials and methods
Study system: The Asteraceae comprises approximately 1 110 genera and 25 000 species. The genus Baccharis belongs to the subtribe Baccharidinae, which is restricted to the American Continent (Barroso 1976). The Baccharidinae probably appeared during Middle Miocene (Boldt 1989) when South American and African continents were totally separated by the Atlantic Ocean (Cox & Moore 1993), justifying its natural occurrence just in the American continent. Baccharis is the largest genus of Baccharidinae (approximately 400 species) and most species are found in the South and Southeast regions of Brazil, suggesting that this region represent the center of the genus origin (Jarvis et al. 1991). B. dracunculifolia (alecrim vassourinha) is a widespread, perennial and woody dioecious shrub, 2-3m high, which occurs in Southern and Southeastern Brazil, Argentina, Uruguay, Paraguay and Bolivia (Espírito-Santo et al. 2003, Fagundes et al. 2005). It grows in open and disturbed habitats, especially along highways and in abandoned pastures. Because it is an evergreen plant, its leaves and branches are important food resources for a variety of insect herbivores, especially coleopterans, heteropterans, hemipterans and orthopterans (Fagundes et al. 2005). Furthermore, the shrub B. dracunculifolia also supports the largest known fauna of galling insects (17 species) in the Neotropics (Fernandes et al. 1996). The degree of habitat disturbance did not influence the richness of galls associated with the host plant B. dracunculifolia (Julião et al. 2005).
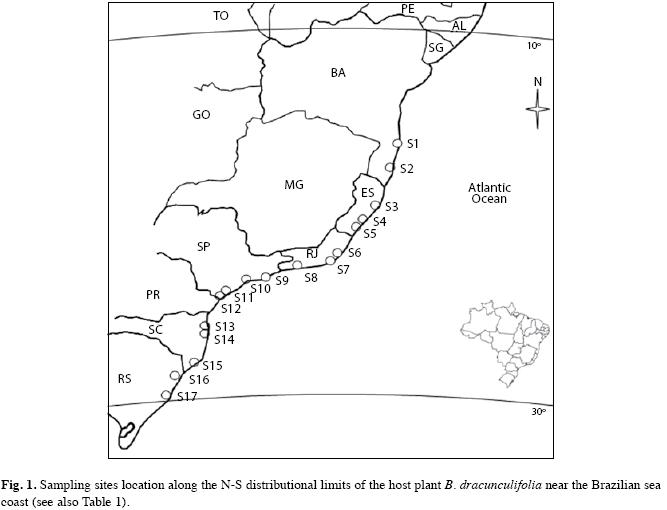
Study areas: The B. dracunculifolia shrubs selected for insect samples belonged to seventeen populations located along the Brazilian sea coast, from the Southern to the Northern distributional limit of the host plant inside Brazil. All plant populations occurred in disturbed environments along highways, within 500m of the seashore and were similar in phenology, size and density. The climate of all the seventeen sample points is broadly influenced by Atlantic Ocean air mass. However, from Northern São Paulo State through Rio de Janeiro, Espírito Santo and Bahia States the climate is moist tropical (Af according Köppen classification). In contrast, from Southern São Paulo State through Parana, Santa Catarina e Rio Grande do Sul States the climate is classified as subtropical (Cfa according Köppen classification). All selected host plant populations were located at similar altitude (06-130m) and were grown on typical seashore sand soil with high levels of moisture and salinity (Table 1, Fig. 1). The surrounding vegetation ranged from 0.3 to 2.0m height and was comprised by grasses and other invasive species belonging principally to the Asteraceae, Convolvulaceae, Malvaceae, Melastomataceae and Solanaceae botanical families. At each host plant population, thirty B. dracunculifolia shrubs were selected to take herbivores samples. The shrubs selected for this were young, non-flowering or fruiting plants with 1.5-2.0m high and stem diameters <15cm.
Herbivore samples: Herbivore insects associated with B. dracunculifolia were censused by direct sampling on the host plant. Thus, each shrub was inspected during ten minutes and all free-feeding herbivore insects and galls observed were handling collected. The herbivores were sampled during the summer season (October of 2001 through January of 2002) in order to minimize the possible effects of climate variation in insect population dynamics. All galling and free feeding herbivore insect samples were taken to the Laboratório de Biologia da conservação at Universidade Estadual de Montes Claros (Unimontes) where they were assigned into morphospecies and identified. We fail to test the insect ability to feed on B. dracunculifolia. Therefore, potentially transient insect herbivores could have been included in data collection and analysis.
The effects of latitudinal variation on the two guilds of insect herbivores were tested using simple linear regressions. For each guild, linear regression analyses were performed using latitudinal variation as the independent variable and the Hill’s number of diversity (N0, N1 and N2) or evenness as dependent variable. The Hill’s number of diversity N0, N1 and N2 represent, respectively, the total number of species (richness), number of abundant species, and number of very abundant species within a species assemblage. Thus, the unity of Hill’s numbers is species, making easy data interpretation and comparisons with other results. The number of abundant species (N1) represent the exponential function of Shannon’s index (N1=eH’), while the number of very abundant species (N2) is the inverse of Simpson’s index (N2=1/S). In addition, evenness was determined by the ratio of very abundant species (N2) to abundant species (N1), known as the modified Hill’s ratio. See Ludwig & Reynolds (1998) for a detailed description of this diversity index and evenness.
Results
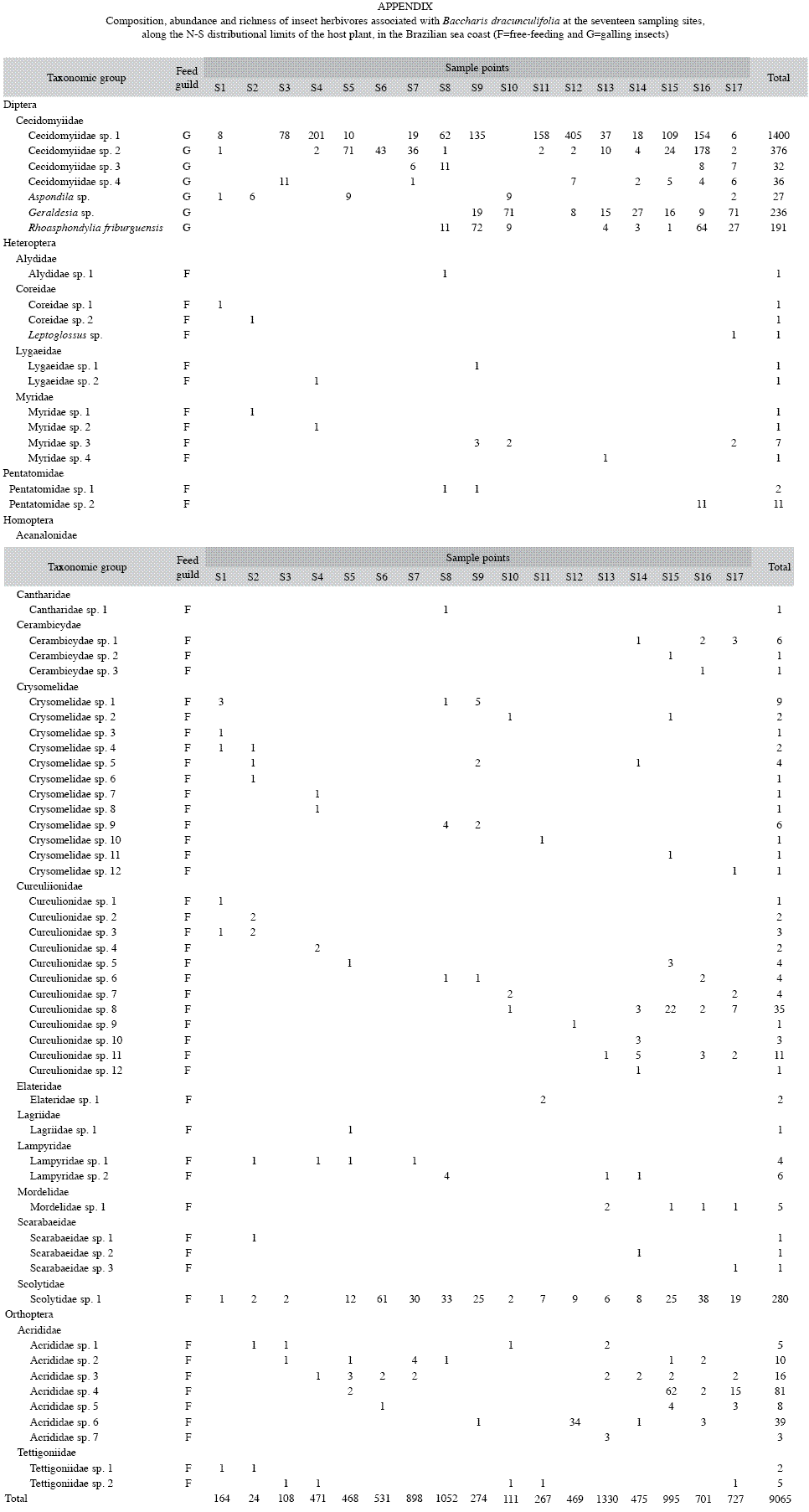
General patterns: A total of 8 201 galls and 864 free-feeding insect herbivores were collected from B. dracunculifolia at the seventeen sample points. The fauna of herbivorous insects associated with B. dracunculifolia was composed by 88 species from 28 families. The insect families with more species were Chrysomelidae (12 species), Curculionidae (12 species), Acrididae (7 species) and Cecidomyiidae (7 species). In general, free-feeding herbivore abundance and frequency were low, but an unidentified species of Scolytidae deserve further studies due to their high abundance and frequency in the samples. This species was commonly observed feeding on meristems and new leaves of host plant (Appendix).
The guild of free-feeding insects represented 90.99% of total insect herbivore species associated with B. dracunculifolia, while gall-forming contributed with 9.01% of the sampled species. On the other hand, gallforming insects represented the most abundant feeding guild (90.48%), whereas free-feeding insects represented just 9.52% of total herbivore insect abundance. In general, the richness of insects per sample site (local richness) was low (mean=14.47, range: 09-23) when compared to total richness (regional richness=88), indicating high species substitution among sample sites. This observed pattern was due mainly to free-feeding, rather than gall-forming insects (Appendix).
Latitudinal gradient hypothesis: The number of gall-inducing insect species associated with B. dracunculifolia decreased significantly towards the North, low latitude, limit of the host plant distribution (F=31.563, p=0.007, r2=67.78). However, the number of abundant species (F=1.469, p=0.245, r2=9.49), very abundant species (F=2.289, p=0.152, r2=14.05) and evenness (F=0.234, p=0.636, r2=1.642) did not vary significantly along the latitudinal gradient (Fig. 2). When free-feeding insect herbivores were analyzed, none of the measures of species diversity (specie richness: F=1.988, p=0.179, r2=11.70; number of abundant species: F=0.372, p=0.551, r2=2.42; number of very abundant species: F=0.005, p=0.944, r2=0.34 and evenness: F=0.234, p=0.636, r2=1.64) were significantly related to variations in latitude (Fig. 3).
Discussion
The number of gall-inducing insect species associated with Baccharis dracunculifolia decreased towards the equator. The observed result did not fit the more common pattern of increasing species diversity as latitude decreases (Willig et al. 2003, Lewinsohn & Roslin 2008). Factors such as geographic distribution, center of origin, taxonomic isolation and local diversity of the host plants have been used to justify the absence of correlation between species diversity of other insect herbivores and latitudinal variations (Cornell 1985, Leather 1986, Lewinsohn & Roslin 2008). We argue that our results are related to the evolutionary history of the host plant and the highly specialized feeding habit of gall-inducing insects. The genus Baccharis is more species-rich in the Southern region of Brazil, that probably corresponds to the genus origin center (Jarvis et al. 1991). Moreover, given the narrow host plant requirements of gall-inducing insects, it would be expected that exchange of gall-inducing insect species between closely related plant species would be easier than exchange between more distantly related plant species (Lawton & Schroder 1977, Leather 1986). Thus, it is possible that the galling insect associated with Baccharis genus radiated into the Southern region of Brazil, justifying the decrease of galling insect richness towards the North, limit of host plant observed in this study.
Species’ ranges may be constrained by both climatic tolerances and barriers to dispersal (e.g. mountains and rivers). In the former case, species can disperse to a new habitat but fail to become established, whereas in the latter case species might never have opportunity to reach the new habitat even though they have attributes needed to persist there (Buckley et al. 2010). In this study, physical barriers are uncommon among the sample sites, but the temperature results higher in Northern regions. Thereby, climatic intolerances should represent a constraint to some galling species establishment in the extreme Northern limit of host plant which would generate the observed pattern of high gall richness in the Southern region. In fact, recent studies indicate that climatic specialization and retention of climatic tolerances over time (niche conservatism hypothesis) might drive species distributions in space (Buckley et al. 2010, Kozak & Wiens 2010). However, adequate experimental design is need to test the effectiveness of niche conservatism to determine the regional distribution of galling species associated with B. dracunculifolia.
The variation in latitude did not influence the richness of free-feeding insect herbivores associated with B. dracunculifolia. This result supports the spatial heterogeneity hypothesis prediction because species richness should not change within structurally similar habitats along a latitudinal gradient. However, Dawidowitz & Rosenzweigh (1998) present four examples (ants, grasshopper, scorpions and mammals) where diversity trends within single biome types change along the latitudinal variation. In contrast, no significant correlations were found between the proportion of mining species and latitude (Sinclair & Hughes 2008). The authors argument that leaf mining is a widespread type of insect herbivory whose distribution patterns are more likely to be influenced by biotic than abiotic factors. Similarly, Andrew & Hughes (2005) found no consistent North to South (tropical to temperate) change in arthropod community structure associated with Acacia trees in Eastern Australia. The findings suggest that different mechanisms operate for different taxa and that it is not wise to generalize the effects of latitude on species diversity across taxa.
Despite of different patterns of species richness observed for gall-inducing and free feeding insects, neither the diversity indices represented by Hill’s number N1 and N2 or evenness of these two guilds were related to latitudinal variation. These patterns suggest that even though species composition and richness can vary among habitats, there is a general structure to the herbivore community associated with B. dracunculifolia (Fagundes et al. 1996, Andrew & Hughes 2005). In fact, there are strong evidences that bottom-up (Faria & Fernandes 2001, Espírito-Santo & Fernandes 2002) and top-down (Fagundes et al. 2005) forces operating in the system can influence the relative abundance of insect herbivores on B. dracunculifolia and regulate diversity at the local level.
The majority of tropical insect herbivores are relatively rare (Price et al. 1995). Generally, just a few groups of insect herbivores with specialized feeding habits, such as some macrolepidoptera (Price et al. 1995), Homoptera (Dansa & Rocha 1992) and galling insects (Fernandes et al. 1996), can be found predictably and at high densities on specific host plants in tropical regions. However, the composition of herbivorous families observed in this study corroborates the general insect composition patterns described in other tropical areas. Usually, Chrysomelidae and Curculionidae are folivores recurrently important components of free-feeding insect communities (Kitching et al. 1997, Basset 2001, Neves et al. 2010), while Cecidomyiidae is dominant among the galling insects (Carneiro et al. 2009).
The general pattern of low density and frequency, specially observed in free-feeding insect guild, probably may be related to high plant diversity and the more polyphagous feeding habits of these insects in the tropics (Strong et al. 1984, Lawton 1991, Lewinsohn & Roslin 2008). The majority of insect herbivores, especially free-feeding insects found associated with B. dracunculifolia, showed low population density and frequency. Low density could be associated with generalist habits of herbivores permitting migration to a variety of host plants, while low species frequency indicates high variation in insect fauna between sample sites. In fact, the majority of the insects found on B. dracunculifolia were restricted to a specific site rather than having a geographic distribution mirroring that of the host plant.
The variation in insect fauna among habitats is determined by their host specificity, interactions with natural enemies and their ability to follow host plant species in space and time and across different environments (Novotny 2009). Thus, it is probable that local events, such as biological interactions, regulate the population dynamics of herbivores associated with B. dracunculifolia. In contrast, the insect fauna collected at each sample site represent just a portion of the local pool of insect herbivores capable of colonizing B. dracunculifolia. Thereby, the insect fauna sample from each site, resulted of large scale events, as speciation, migration and coevolution, while at local level, the population of these insects is regulated by ecological forces which operate in the system. Understanding the core of the relative role of ecological and evolutionary processes will be essential to predict general community structure, and advance in new strategies for insect conservation (Perry et al. 1998, Lawes et al. 2000).
Acknowledgments
We thank B.G. Madeira, P.W. Price and T.M. Lewinsohn for insightful comments on earlier drafts of the manuscript, and two other anonymous referees who made valuable contributions to this paper. Financial support by the Fapemig (the Minas Gerais Research Foundation) and CNPq (the National Research Council of Brazil).
References
Andrew, N.R. & L. Hughes. 2005. Arthropod community structure along a latitudinal gradient: implications for future impacts of climate change. Aust. Ecol. 30: 281-287. [ Links ]
Barroso, G.M. 1976. Compositae – Subtribo Baccharidinae Hoffmann: estudo das espécies ocorrentes no Brasil. Rodriguésia 28: 3-273. [ Links ]
Basset, Y. 2001. Invertebrates in the canopy of tropical rain forests: How much do we really know? Plant Ecol. 153: 87-107. [ Links ]
Boldt, P.E. 1989. Baccharis (Asteraceae): a review of its taxonomy, phytochemistry, ecology, economic status, natural enemies and potential for its biological control in the Unites States. USDA, Agricultural Research Service Grassland, Temple, Texas, USA. [ Links ]
Buckley, L.B., T.J. Davies, D.D. Ackerly, N.J.B. Kraft, S.P. Harrison, B.L. Anacker, H.V. Cornell, E.I. Damschen, J. Grytnes, B.A. Hawkins, C.M. McCain, P.R. Stephens & J.J. Wiens. 2010. Phylogeny, niche conservatism and the latitudinal diversity gradient in mammals. Proc. Roy. Soc. Lond. B. Biol Sci. 277: 2131-2138. [ Links ]
Carneiro, M.A.A., R.A.X. Borges, A.P.A. Araújo & G.W. Fernandes. 2009. Insetos indutores de galhas da porção sul da Cadeia do Espinhaço, Minas Gerais, Brasil. Rev. Bras. Entomol. 53: 570-592. [ Links ]
Cornell, H.V. 1985. Local and regional richness of Cynipine gall wasps on California oaks. Ecology 66: 1247-60. [ Links ]
Cox, C.B. & P.D. Moore. 1993. Biogeography: an ecological and evolutionary approach. Blackwell, Berlin, Germany. [ Links ]
Dajoz, R. 2000. Insects and Forests: The role and diversity of insects in the forest environment. Lavoisier, Paris, France. [ Links ]
Dansa, C.V.A. & C.F.D. Rocha. 1992. An ant-membracidplant interaction in a cerrado area of Brazil. J. Trop. Ecol. 8: 339-348. [ Links ]
Davidowitz, G. & M.L. Rosenzweig. 1998. The latitudinal gradient of species diversity among North American grasshoppers (Acrididae) within a single habitat: a test of the spatial heterogeneity hypothesis. J. Biogeogr. 25: 553-560. [ Links ]
Dixon, A.F.G., P. Kindlman, J. Leps & J. Holman. 1987. Why there are so few species of aphids, especially in the tropics. Am. Nat. 129: 580-592. [ Links ]
Espírito-Santo, M.M. & G.W. Fernandes. 2002. Host plant effects on the development and survivorship of the galling insect Neopelma baccharidis (Homoptera: Psyllidae). Austral Ecol. 27: 249-57. [ Links ]
Espírito-Santo, M.M., B.G. Madeira, F.S. Neves, M.L. Faria, M. Fagundes & G.W. Fernandes. 2003. Sexual differences in reproductive phenology and their consequences to the demography of Baccharis dracunculifolia (Asteraceae), a dioecious tropical shrub. Ann.Bot. 91: 13-19. [ Links ]
Fagundes, M., F.S. Neves & G.W. Fernandes. 2005. Direct and indirect interactions involving ants, insect herbivores and parasitoids on the host plant Baccharis dracunculifolia (Asteraceae). Ecol. Entomol. 30: 18-25. [ Links ]
Fagundes, M., J.C. Zanuncio, F.S. Lopes & P. Marco Jr. 1996. Comunidades de lepidópteros noturnos desfolhadores de eucalipto em três regiões do cerrado de Minas Gerais. Rev. Bras. Zool. 13: 763-71. [ Links ]
Faria, M.L. & G.W. Fernandes. 2001. Vigour of a dioecious shrub and attack by a galling herbivore. Ecol. Entomol. 26: 37-45. [ Links ]
Fernandes, G.W. & P.W. Price. 1988. Biogeographical gradients in galling species richness: tests of hypotheses. Oecologia 76: 161-167. [ Links ]
Fernandes, G.W., M.A.A. Carneiro, A.C.F. Lara, L.R. Allain, G.I. Andrade, G. Julião, T.R. Reis & I.M. Silva. 1996. Galling insects on neotropical species of Baccharis (Asteraceae). Trop. Zool. 9: 315-32. [ Links ]
Frenzel, M. & R. Brandl. 1998. Diversity and composition of phytophagous insect guilds on Brassicaceae. Oecologia 113: 391-399. [ Links ]
Gaston, K.J. & P.H. Williams. 1996. Spatial patterns in taxonomic diversity, p. 202-229. In K.J. Gaston (ed.). Biodiversity: biology of numbers and differences. Blackwell Science, London, England. [ Links ]
Golden, D.M. & T.O. Crist. 1999. Experimental effects of habitat fragmentation on old-field canopy insects: community, guild and species responses. Oecologia 118: 371-380. [ Links ]
Hillebrand, H. 2004. On the generality of the latitudinal diversity gradient. Am. Nat. 163: 192-211. [ Links ]
Jarvis, B.B., N. Mokhtari-Rejali, E.P. Schenkel, C.S. Barros & N.I. Matzenbacher. 1991. Tricothecene mycotoxins from Brazilian Baccharis species. Phytochemistry 30: 789-797. [ Links ]
Julião, G.R., G.W. Fernandes, D. Negreiros, L. Bedê & R.C. Araújo. 2005. Insetos galhadores associados a duas espécies de plantas invasoras de áreas urbanas e
peri-urbanas. Rev. Bras. Entomol. 49: 97-106. [ Links ]
Kitching, R.L., H. Mitchell, G. Morse & G. Thebaud. 1997. Determinants of species richness in assemblages of canopy arthropods in rainforests, p. 131-150. In N.E. Stork, J. Adis & R.K. Didham (eds.). Canopy Arthropods. Chapman & Hall, London, United Kingdom. [ Links ]
Koricheva, J., S. Larsson & E. Haukioja. 1998. Insect performance on experimentally stressed woody plants: a meta-analysis. Annu. Rev. Entomol. 43: 195-216. [ Links ]
Kozak, K.H. & J.J. Wiens. 2010. Niche conservatism drives elevational diversity patterns in Appalachian salamanders. Am. Nat. 176: 40-54. [ Links ]
Lawes, M.J., H.T. Eeley & S.E. Piper. 2000. The relationship between local and regional diversity of indigenous forest fauna in KwaZulu-Natal Province, South Africa. Biodivers. Conserv. 9: 683-705. [ Links ]
Lawton, J.H. & D. Schroder. 1977. Effects of plant type, size of geographical range and taxonomic isolation on number of insect species associated with British plants. Nature 265: 137-140. [ Links ]
Lawton, J.H. 1991. Species richness, population abundances, and body sizes in insect communities: tropical versus temperate comparisons, p. 71-89. In P.W. Price, T.M. Lewinsohn, G.W. Fernandes & W.W. Benson (eds.). Plant-animal interactions: evolutionary ecology in tropical and temperate regions. Jonh Wiley and Sons, New York, New York, USA. [ Links ]
Leather, S.R. 1986. Insect species richness of the British Rosaceae: the importance of host range, plant architecture, age of establishment, taxonomic isolation and specie-area relationships. J. Anim. Ecol. 55: 841-860. [ Links ]
Lewinsohn, T.M. & T. Roslin. 2008. Four ways towards tropical herbivore megadiversity. Ecol. Lett. 11:398-416. [ Links ]
Ludwig, J.A. & J.F. Reynolds. 1988. Statistical ecology: a primer on methods and computing. John Wiley and Sons, New York, New York, USA. [ Links ]
Majer, J.D., R.L. Kitching, B.E. Heterick, K. Hurley & K.E.C. Brennan. 2001. North-South patterns within arboreal ant assemblages from rain forest in eastern Australia. Biotropica 33: 643-661. [ Links ]
Neves, F.S., L.S. Araújo, M. Fagundes, M.M. Espírito-Santo, G.W. Fernandes, G.A. Sánchez-Azofeifa & M. Quesada. 2010. Canopy herbivory and insect herbivore diversity in a dry forest-savana transition in Brazil. Biotropica 42: 112-118. [ Links ]
Novotny, V. & Y. Basset. 2005. Host specificity of insect herbivores in tropical forests. Proc. Roy. Soc. Lond. B. Biol Sci. 272: 1083-1090. [ Links ]
Novotny, V. 2009. Beta diversity of plant-insect food webs in tropical forest: a conceptual framework. Insect Conserv. Biodiv. 2: 5-9. [ Links ]
Otte, D. 1976. Species richness patterns of new world desert grasshoppers in relation to plant diversity. J. Biogeogr. 3: 197-209. [ Links ]
Perry, G., G.H. Rodda, T.H. Fritts & T.R. Sharp. 1998. The lizard fauna of Guam’s fringing islets: island Biogeography, phylogenetic history, and conservation implications. Global Ecol. Biogeogr. 7: 353-365. [ Links ]
Pianka, E.R. 1966. Latitudinal gradients in species diversity: a review of concepts. Am. Nat. 100: 33-46. [ Links ]
Price, P.W., I.R. Diniz, H.C. Moraes & E.S.A. Marques. 1995. The abundance of insect herbivore species in the tropics: the high local richness of rare species. Biotropica 27: 468-478. [ Links ]
Ribeiro, S.P., M.A.A. Carneiro & G.W. Fernandes. 1998. Free-feeding insect herbivores along environmental gradients in Serra do Cipó: basis for a management plan. J. Insect. Conservat. 2: 107-118. [ Links ]
Rohde, K. 1998. Latitudinal gradients in species diversity. Area matters, but so much? Oikos 82: 184-90. [ Links ]
Shepherd, U.L. 1998. A comparison of species diversity and morphological diversity across the North American latitudinal gradient. J. Biogeogr. 25: 19-29. [ Links ]
Sime, K.R. & A.V.Z. Brower. 1998. Explaining the latitudinal gradient anomaly in ichneumonid species richness: evidence from butterflies. J. Anim. Ecol. 67: 387-99. [ Links ]
Sinclair, R.J. & L. Hughes. 2008. Incidence of leaf mining in different vegetation types across rainfall, canopy cover and latitudinal gradients. Aust. Ecol. 33: 353-360. [ Links ]
Strong, D.R., J.H. Lawton & T.R.E. Southwood. 1984. Insect on Plants: Community Patterns and Mechanisms. Blackwell, London, England. [ Links ]
Terborgh, J. 1992. Diversity and the Tropical Rain Forest. Scientific American Library, New York, New York, USA. [ Links ]
Thomaz, S.M., D. Dibble, L.R. Evangelista, J. Higuti & L.M. Bini. 2008. Influence of aquatic macrophyte habitat complexity on invertebrate abundance and richness in tropical lagoons. Freshwat. Biol. 53: 358-367. [ Links ]
Willig, M.R., D.M. Kaufman & R.D. Stevens. 2003. Latitudinal gradient of biodiversity: patterns, process, scale, and synthesis. Annu. Rev. Ecol. Syst. 34: 273-309. [ Links ]
Whitehouse, M.E.A., S. Fardwick, B.C.G. Scholz, A.J. Annells, P.R. Grundy & S. Harden. 2009. Evidence of a latitudinal gradient in spider diversity in Australian cotton. Aust. Ecol. 34: 10-23. [ Links ]
Correspondencia a: Marcílio Fagundes. Laboratório de Biologia da Conservação DBG/ CCBS, Universidade Estadual de Montes Claros, Montes Claros, MG, Brazil, 39401-089; marcilio.fagundes@gmail.com
G. Wilson Fernandes. Ecologia Evolutiva & Biodiversidade/DBG/ICB/Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil; gwfernandes@gmail.com
Received 21-VII-2010. Corrected 07-I-2011. Accepted 08-II-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}