










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.2 San José Jun. 2011
Genetic molecular analysis of Coffea arabica (Rubiaceae) hybrids using SRAP markers
Manoj Kumar Mishra, Narayana Suresh, Asha M. Bhat, Nayani Suryaprakash, Saya Satheesh Kumar, Anil Kumar & Jayarama
Central Coffee Research Institute, Coffee Research Station -577117, Chikmagalur- Dist, Karnataka, India; manojmishra.m@gmail.com, sureshccri@gmail.com, ashabhat_bt@yahoo.co.in, nayanirao@gmail.com, sayasath-eesh@gmail.com, anilsirsi@yahoo.com, ccrijaya@gmail.com
Dirección de correspondencia
Abstract
In Coffea arabica (arabica coffee), the phenotypic as well as genetic variability has been found low because of the narrow genetic basis and self fertile nature of the species. Because of high similarity in phenotypic appearance among the majority of arabica collections, selection of parental lines for inter-varietals hybridization and identification of resultant hybrids at an early stage of plant growth is difficult. DNA markers are known to be reliable in identifying closely related cultivars and hybrids. Sequence Related Amplified Polymorphism (SRAP) is a new molecular marker technology developed based on PCR. In this paper, sixty arabica-hybrid progenies belonging to six crosses were analyzed using 31 highly polymorphic SRAP markers. The analysis revealed seven types of SRAP marker profiles which are useful in discriminating the parents and hybrids. The number of bands amplified per primer pair ranges from 6.13 to 8.58 with average number of seven bands. Among six hybrid combinations, percentage of bands shared between hybrids and their parents ranged from 66.29% to 85.71% with polymorphic bands varied from 27.64% to 60.0%. Percentage of hybrid specific fragments obtained in various hybrid combinations ranged from 0.71% to 10.86% and ascribed to the consequence of meiotic recombination. Based on the similarity index calculation, it was observed that F1 hybrids share maximum number of bands with the female parent compared to male parent. The results obtained in the present study revealed the effectiveness of SRAP technique in cultivar identification and hybrid analysis in this coffee species. Rev. Biol. Trop. 59 (2): 607-617. Epub 2011 June 01.
Key words: Coffea arabica, SRAP molecular marker, hybrid analysis, genetic inheritance.
Resumen
En Coffea arabica (café arabica), el fenotipo y la variabilidad genética son bajos debido a la estrecha base genética y la autofecundación de la especie. Por su alta similitud fenotípica entre la mayoría de las colecciones de arábica, la selección de líneas parentales para hibridación entre variedades, y la identificación de los híbridos resultantes en una fase inicial de crecimiento, es difícil. Para la identificación de variedades estrechamente relacionadas y sus híbridos, los marcadores de ADN son confiables, pero los polimorfismos de amplificación de secuencias relacionadas (SRAP, por sus siglas en inglés) constituyen una nueva tecnología de marcadores moleculares basada en PCR. En este trabajo, sesenta progenies arábica híbridas, pertenecientes a seis cruces, fueron analizadas utilizando 31 marcadores altamente polimórficos. El análisis reveló siete tipos de perfiles de marcadores que son útiles en la discriminación de los progenitores y los híbridos. El número de bandas amplificadas por pares de cebadores estuvo entre 6.13 a 8.58 con un promedio de siete bandas. Entre las seis combinaciones de híbridos, el porcentaje de bandas compartidas entre híbridos y sus progenitores estuvo entre 66.29% y 85.71% con bandas polimórficas que variaron entre 27.64% y 60.0%. El porcentaje de fragmentos híbridos específicos obtenidos en diversas combinaciones híbridas varió entre 0.71% y 10.86% lo que se atribuye a la recombinación meiótica. Con base en el cálculo del índice de similitud, se observó que los híbridos F1 compartieron un número máximo de bandas con el progenitor femenino que con el masculino. Los resultados obtenidos en este estudio muestran la eficacia de la técnica de SRAP en la identificación de cultivos e híbridos de esta especie de café.
Palabras clave: Coffea arabica, marcadores moleculares SRAP, análisis de híbridos, herencia genética.
Coffee is one of the most important agricultural commodities in international trade in terms of volume and foreign exchange. It is grown in about 80 countries spanning over 10.2 million hectars land in the tropical and sub-tropical regions of the world, especially in Africa, Asia and Latin America. The genus Coffea belongs to the family Rubiaceae and consists of more than 100 species (Davis et al. 2006) of which only two species Coffea arabica (known as arabica coffee) and Coffea canephora (known as robusta coffee) are commercially cultivated. All the coffee species are diploids (2n=2x=22) except C. arabica which is an allotetraploid (2n=4x=44) species. In the consumer market, arabica coffee is preferred for its beverage quality, aromatic characteristics and low caffeine content compared to robusta, which is characterized by a stronger bitterness, and higher caffeine content. Arabica contributes towards 65% of global coffee production (Lecolier et al. 2009).
Arabica coffee is susceptible to many pest and diseases, which limits its production and productivity. Therefore, improvement of the arabica coffee for pest and disease tolerance is the main objective of plant breeding programs. One of the major constraints that limits the arabica coffee breeding is the selection of parental lines and identification of hybrids at an early stage of plant growth based on morphological traits. This is because most of the commercial arabica cultivars are morphologically identical and not easily distinguishable from each other. Uniformity for morphological traits in C. arabica could be attributed to origin of the species, the narrow genetic basis and self-fertile nature. In coffee, there are no reliable methods for identification of hybrid genotypes and only in certain progenies some of the phenotypic traits such as leaf and fruit characters might give some indication. However, as coffee is grown under shade and in different agro-climatic zones, the stability of phenotypic characters is critical because of genotype environment interaction. In view of the above, development of an alternative technique that is reliable, quick and that efficiently discriminates coffee hybrids and parents, will greatly help in the hybridization and selection process, and thereby speed up the coffee breeding programs. Among various markers available for genetic analysis in plants, molecular markers are more efficient, precise and reliable for discriminating closely related species and cultivars and therefore widely used in marker assisted breeding programs. Among many types of molecular markers, sequence related amplified polymorphism (SRAP) has been demonstrated to be a useful tool in genetic analysis of different plant species (Li & Quiros 2001, Ferriol et al. 2003, Esposito et al. 2007, Merotto et al. 2009). SRAP is a PCR based marker system that preferentially targets coding sequences randomly distributed throughout genome (Li & Quiros 2001). Forward and reverse primers used in SRAP, preferentially amplify exonic and intronic regions of the genome respectively and uncover polymorphic sequences resulting from variations in the length of introns, promoters and spacers among different populations and genotypes. Zaefizadeh & Golieb (2009) reported that SRAP markers possess multiloci and multi-allelic features, which make them potentially more efficient for genetic diversity analysis, gene mapping and fingerprinting of genotypes. Recently, SRAP markers were also used for identification of cultivars and hybrids (Liu et al. 2007, Hao et al. 2008, Xuan et al. 2008). SRAP is highly reproducible and comparatively less expensive than other types of markers (Cravero et al. 2007). However, the potential of SRAP marker has not yet been tested in coffee. In the present study, SRAP markers were employed for the genetic analysis of six arabica coffee hybrids to ascertain its suitability and efficiency.
Materials and Methods
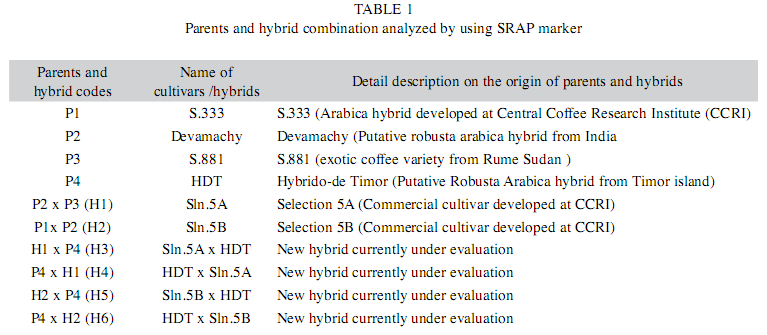
Plant materials: Sixty inter-varietals arabica F1 hybrids derived from six crosses and their respective parents constitute the material for the present study. These materials are grown at the experimental plots of Central Coffee Research Institute, India. The details of the plant material and their parentage is given in Table 1.
DNA extraction: Genomic DNA was extracted from fresh young leaves using a modified CTAB method (Murray & Thomson 1980). About 200 mg of leaf tissue was grounded to fine powder in liquid nitrogen, and then transferred to 5 mL extraction buffer (2% CTAB, 100mM Tris HCl (pH 8.0), 25mM EDTA, 2M NaCl, 0.1% beta-Mercaptoethanol). The suspension was incubated at 60°C for one hour with occasional shaking, and then cooled down to room temperature and centrifuged at 6 000rpm for 20min. The supernatant was transferred into a new tube and extracted twice with an equal volume of chloroform-isoamyl alcohol (24:1) followed by precipitation with 0.7 vol. of isopropanol at room temperature for 30min, and a centrifugation at 8 000rpm for 20min at 4°C. The pellet formed was washed with 75% (v/v) ethanol for 10min and the resuspended in TE buffer.
The concentration of DNA was measured using 0.8% w/v agarose gel stained with ethidium bromide and quantified by Uv spectrophotometry at 260nm and 280nm. Then purity of DNA was determined by calculatingthe ratio of absorbance at 260/280nm. For PCR templates, DNA were diluted to 10ng/µL and stored at -30°C until use.
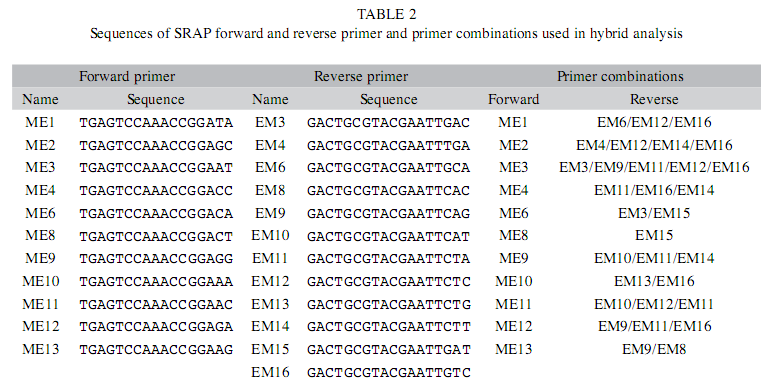
Amplification of SRAP markers: SRAP primers used in this study consisted of 13 forward and 16 reverse primers of Li & Quiros (2001) and their sequences are listed in Table 1. All the primers were synthesized by Sigma-Aldrich and 72 primer combinations were chosen for preliminary amplification (Table 2). DNA amplification was carried out in a 20µL reaction volume containing 2.0µL 10x PCR buffer (750mM Tris HCl (pH 8.8), 0.01%Tween 20, 200 mM NH4 (SO4)2), 30ng genomic DNA, 0.3µm of each primer, 2.5mM MgCl2, 200µM each dNTP and 1.0U/µL of Taq DNA polymerase. The thermal cycling profile used for all reactions was 96°C for 4 min, followed by five cycles at 94°C for one min, 35°C for 1.15min and 72°C for 2min, 30 cycles at 94°C for one min, 50°C for 1.15min and 72°C for 2min and a final extension at 72°C for 10min. The amplification products were analyzed by electrophoresis on a 2% (w/v) agarose gel using 1x TAE running buffer and detected by staining with ethidium bromide. The gel profiles were viewed under Uv-transilluminator (SYNGENE) and documented using the Gene Snap software program. SRAP analysis was repeated twice for each DNA sample.
SRAP Analysis: Thirty-one SRAP primer combinations, which showed polymorphism and consistent amplifications, were selected and are listed in Table 2. Amplified SRAP markers were scored as present (1) or absent (0) for each sample. Ambiguous bands that could not be easily distinguished were not scored. The data developed by manual scoring of the SRAP fingerprints were used for the genetic analysis of the F1 hybrids and parents. The similarity of samples was calculated as follows: Similarity=2NAB/NA+NB, NAB is the number of bands shared by individuals A & B &, NA & NB are the number of bands in individuals A & B respectively (Chapco et al. 1992, Wild et al. 1992).
Results
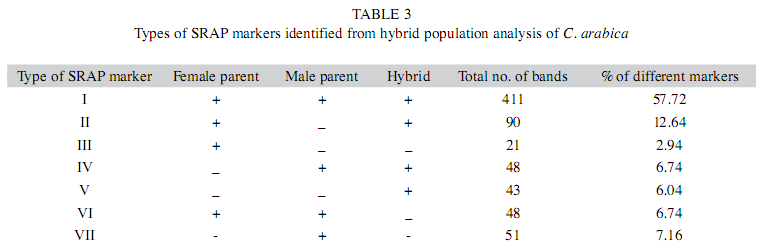
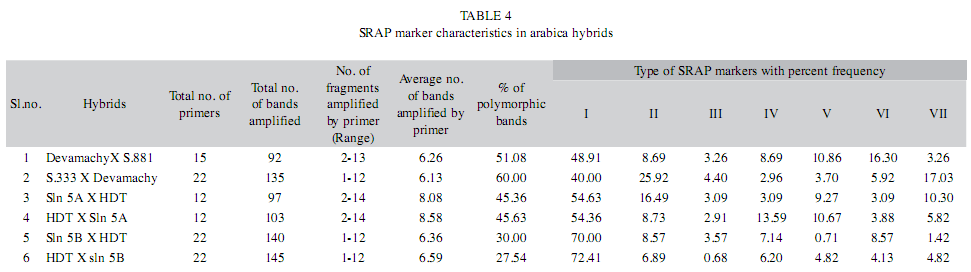
Extent of polymorphism: In the present study, 72 SRAP primer combinations were screened (data not shown) of which 31 pairs showed polymorphism between the parents and therefore were used for further analysis of hybrids (Table 2). These 31 pairs of primers amplified seven different types of SRAP markers at variable frequency in six arabica hybrids and their respective parents (Table 3). Among the seven different types of markers, Type I and Type II are more frequent comprising about 57.72% and 12.64% of the total markers respectively (Table 3). The number of primers used for genetic analysis of six arabica hybrids and their parents varies from 12 to 22 (Table 4). The average number of bands amplified per primer in different hybrids ranges from 6.13 to 8.58 with average number of seven bands amplified per primer pair (Table 4). In hybrid combination of HDT x Sln.5A and its reciprocal cross, the average number of bands amplified by a primer is higher than any other hybrids (Table 4). In different hybrid combinations, the percentage of polymorphic bands ranged from 27.54% (HDT x Sln. 5B) to 60% (S.333 x Devamachy) with a mean of 43.26%. Among the polymorphic marker types, Type II markers are comparatively more frequent (12.64%) than other types of polymorphic markers.
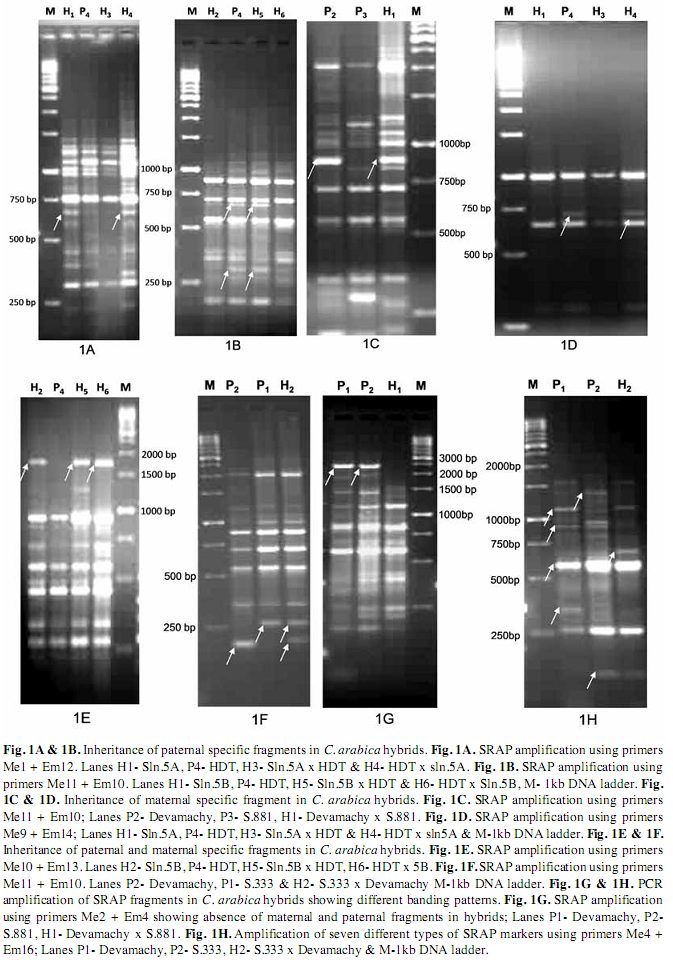
Parents and hybrid identification: The seven different types of SRAP markers obtained was used for discriminating the parents and hybrid authentication. Out of these seven types of markers, Types I, II and IV are good markers for hybrid identification. The Type IV marker is especially very effective and can unambiguously identify the true hybrid because of the presence of a male specific band in the hybrids. Similarly, Type III and Type VII are effective markers for identifying true female and male parents respectively in the hybrid population. The primer pairs ME1 and EM12 amplified a cultivar specific band in Sln.5A that is inherited as paternal fragment (Type IV) in HDT x Sln.5A hybrid but not in the reciprocal hybrid (Sln.5A x HDT) where it is used as female parent (Fig. 1A). Similarly, using the primer combinations of ME11 and EM10, specific SRAP fragment of HDT is inherited as paternal specific fragment (Type IV) in Sln.5B x HDT hybrid combinations (Fig. 1B).
In the present study, amplification of maternal specific fragment is also observed in Devamachy x S.881 and HDT x Sln.5A hybrid combinations (Type II) (Figs. 1C, 1D). In 5B x HDT and its reciprocal hybrid, cultivar specific band of 5B amplified by primer combination ME10 and EM13 was inherited as both maternal and paternal (Type II and Type IV) specific fragments (Fig. 1E). Similarly, the SRAP primer combinations of ME11 + EM10 amplified the cultivar specific band in Devamachy and S.333 which are inherited as paternal and maternal specific fragments in S.333 x Devamachy hybrid (Fig. 1F). Further, it is revealed that SRAP banding patterns in the hybrids is not completely additive as 2.94 % to 7.16% of SRAP markers from parents were not found in hybrid combinations (Fig. 1G). Interestingly, the primer combinations of ME4 and EM16 amplified all the seven different types of SRAP markers in S333 x Devamachy hybrid (Fig. 1H).
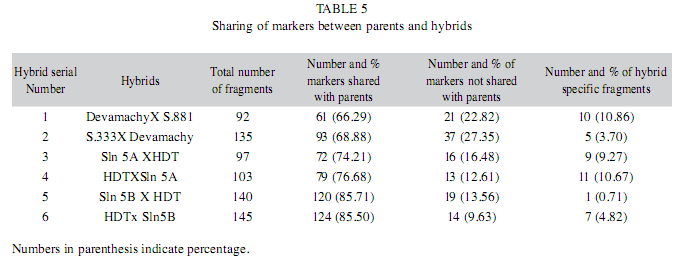
Genetic similarities between parents and hybrids: The presence and absence of marker profiles in six arabica hybrids and parents were computed and the sharing of bands between parents were calculated and presented in Table 5. It is observed that the percentage of markers shared between parents and hybrids varied from 66.29% (Devamachy x S.881) to 85.71% (Sln.5B x HDT). Interestingly, the majority of the F1 hybrids shared a maximum number of bands with the female parent compared to the male parent. In Devamchy x S. 881 hybrid, male and female parents shared equal number of bands with the hybrid, where as in HDT x Sln.5A, higher percentage of male specific bands is shared by the hybrid compared to the female specific bands. Based on the banding pattern, the similarity index was calculated between different hybrids and parents and presented in Table 6. In the hybrid population, parental bands at the range of 9.63% to 27.35% with an average of 17.07% are not displayed. However, all the F1 hybrids displayed several hybrid specific fragments, which are not present in any of the parents. The percentage of hybrid specific bands ranges from 0.7% in hybrid 5B x HDT to 10.86% in hybrid Devamachy x S.881 (Table 5).
Discussion
Due to the advancement in DNA fingerprinting techniques, diagnostic molecular markers are now increasingly available for use in molecular taxonomy, cultivar identification and marker assisted selection in plants. Among various molecular markers, SRAP is a relatively new PCR marker extensively used for germplasm characterization, cultivar identification, molecular mapping and gene cloning in plants during recent years (Ferriol et al. 2004, Gulsen et al. 2005, Sun et al. 2006, Vandemark et al. 2006, Esposito et al. 2007, Mutlu et al. 2008). However, only a few reports are available on the utility of SRAP marker in identification and authentication of hybrids in plants (Hale et al. 2006, Liu et al. 2007, Xuan et al. 2008). Liu et al. (2007) suggested that co dominant markers are best suited for hybrid analysis. Xuan et al. (2008) demonstrated the effectiveness of SRAP marker in genetic analysis and hybrid identification of Zoysia hybrids. However, SRAP being a highly informative and reproducible marker has not been employed in coffee for cultivar identification and hybrid analysis studies. In this study, genotype/variety specific SRAP markers were identified in S.333, S.881, Devamachy, Sln.5A, Sln.5B and HDT. Identification of genotype specific SRAP marker has helped in identification of intervarietal hybrids including the reciprocal ones which was otherwise difficult to identify using phenotypic markers.
The percentage of polymorphic bands amplified by primers in different arabica hybrids ranged from 27.54 % to 60% with an average of 43%. The polymorphism observed is relatively low compared to other plants such as alfalfa (Vandemark et al. 2006), paeonia (Hao et al. 2008), pinus (Feng et al. 2009), mulberry (Zhao et al. 2009) using SRAP markers. The origin of C. arabica cultivars from a narrow genetic base coupled with self-compatible mating system and inbreeding are the major factors for the observed low genetic polymorphism. In general, the low level of polymorphism in C. arabica is consistent with the earlier observation using RAPD markers (Lashermes et al. 1996).
Based on the presence/absence of bands in parents and hybrids, SRAP marker could be classified into seven types (Table 3). Among the seven different types of SRAP markers obtained, Type I, Type II are more frequent compared to other types of marker and together they account for more than 70% of total fragments. In Type I marker, common fragments of both maternal and paternal parents are inherited and displayed by the hybrids. This is expected because of the combination of parental genomes in hybrids. Similarly the high percentage of type II marker (12.64%) showing the maternal fragments in hybrids could be related to the nature of hybrids in coffee showing greater influence of maternal genome in hybrids. All the three Type I, Type II and Type IV are good markers for hybrid identification especially the Type Iv that is very effective and can unambiguously identify the true hybrid because of the presence of male specific band in the hybrids. Similarly, Type III and Type VII are very effective markers for identifying true female and male parents respectively in the hybrid population.
The sharing of SRAP fragments between parents and hybrids of C. arabica ranged from the lowest 66.29% in Devamachy x S.881 hybrid to highest 85.71% in Sln.5B x HDT. The high percentage of sharing of bands between parents and hybrids is due to the close similarity in banding patterns between the male and female parents involved in the generation of hybrids as well as the inheritance of the common bands in hybrids. In chrysanthemum hybrids, maximum percent of band sharing between parents and hybrids was observed to be 48.9% using RAPD marker (Huang et al. 2000).
It is revealed that SRAP banding patterns in the hybrids is not completely additive as 9.63 % to 27.65% of SRAP markers from parents, which constitute of Type III, Type VI and Type VII, were not found in hybrid combinations. Using the SRAP markers, Xuan et al. (2008) observed a similar type of phenomenon in Zoysia hybrids. The non-inheritance of parental bands in hybrids was observed in Chrysanthemum (Huang et al. 2000) and cotton (Ali et al. 2008) using RAPD markers and ascribed to the segregation of heterozygous chromosomes during meiosis. Polymorphism in SRAP assay may result from small insertions and deletions or changes in nucleotide sequences (Li & Quirus 2001). Therefore, differences in banding pattern from parents to offspring may be the result of DNA recombination, mutation or random segregation of chromosomes in meiosis during hybridization (Darnell et al. 1990, Huchett & Botha 1995). It is also suggested that crossing over of chromosomes during meiosis could result in the loss of priming sites in the hybrids and therefore markers are present in parents but not in offspring (Smith et al. 1996).
The bands of non-parental origin (Type V) were detected in a range of 0.71 % to 10.86% in different hybrids (Table 4). As discussed earlier, these hybrid specific bands might have been resulted due to recombination and/or mutational events during meiosis following hybridization (Darnell et al. 1990, Huchett & Botha 1995). However, the frequency of hybrid specific bands is low compared to the earlier studies (Hunt & Page 1992, Ayliffe et al. 1994, Novy & vorsa 1996) using RAPD marker. Another important observation made is the relatedness among the parents and their hybrids based on the marker profiles. From the similarity matrix of six hybrid combinations of C. arabica, it was observed that majority of the hybrids such as H2, H3, H5 and H6 were close to the female parent than the male parent. However, H1 hybrid share equal number of bands with male and female parent. The H4 (HDT x Sln.5A) hybrid is more similar to the male parent in banding pattern than that of the female parent. A close look in to the similarity matrix has revealed that both hybrids H4 (HDT x Sln.5A) and H3 (Sln.5A x HDT) are closer to Sln.5A irrespective of that it is used as male or female parent. However, in both H5 (Sln.5B x HDT) and H6 (HDT x Sln.5B) hybrids which are reciprocal to each other, hybrids are closer to the female parent. These clearly indicate that the impact of recombination during meiosis varies in different hybrids and determine the SRAP profile pattern.
The identification of cultivars and breeding lines is very important for plant variety registration and protection of plant breeder’s right. In coffee, morphological parameters are very often used to discriminate the varieties and hybrids. However, this exercise is not straight; need lot of field experience and very often misleading. Furthermore, coffee is a perennial plant and need at least 5-7 years for attaining reproductive maturity for evaluation of both vegetative and reproductive characters. Therefore it is critical to identify suitable marker, which can identify cultivars/hybrids at early stage of plant growth. The present study has revealed SRAP marker approach is highly efficient and reproducible not only for identification and authentication of hybrids but also useful for marker inheritance and hybrid progeny analysis in coffee.
References
Ali, M.A., M.T. Seyal, S.I. Awan, S. Niaz, S. Ali & A. Abbas. 2008. Hybrid authentication in upland cotton through RAPD analysis. Aus. J. crop Sci. 2: 141-149. [ Links ]
Ayliffe, M.A., G.J. Lawrence, G.J. Ellis & A.J. Pryor. 1994. Heteroduplex molecules formed between allelic sequences cause nonparental RAPD bands. Nucleic Acids Res. 22: 1632-1636. [ Links ]
Chapco, W., N.W.Ashton, R.K.B. Martel & N. Antonishyn. 1992. A feasibility study of the use of random amplified polymorphic DNA in the population genetics and systematics of grasshoppers. Genome 35: 569-574. [ Links ]
Cravero, V., E. Martin & E. Cointry. 2007. Genetic diversity of Cynara cardunculus determined by Sequence Related Amplified Polymorphism Markers. J. Amer Soc. Hort. Sci. 132: 1-5. [ Links ]
Darnell, J.E., D. Baltimore & H.F. Lodish. 1990. Molecular cell biology. Scientific American Books, New York, USA. [ Links ]
Davis, A.P., R. Govaerts, D.M. Bridson & P. Stoffelen. 2006. An annotated checklist of the genus Coffea L. (Rubiaceae). Bot. J. Linn. Soc. 152: 465-512. [ Links ]
Esposito, M.A., E.A. Martin, V.P. Cravero & E. Cointry. 2007. Characterization of pea accessions by SRAP’s markers. Sci. Hort. 113: 329-335. [ Links ]
Feng, F., M. Chen, D. Zhang, X. Sui & S. Han. 2009. Application of SRAP in the genetic diversity of Pinus koraiensis of different provenances. Afr. J. Biotech. 8: 1000-1008. [ Links ]
Ferriol, M., B. Pico & F. Nuez. 2003. Genetic diversity of a germplasm collection of Cucurbita pepo using SRAP and AFLP markers. Theor. Appl. Genet. 107: 271-282. [ Links ]
Ferriol, M., B. Pico, P.F. De Cordova & F. Nuez. 2003. Genetic diversity of a germplasm collection of Squash (Cucurbita moschata) determined by SRAP and AFLP markers. Crop Sci. 44: 653-664. [ Links ]
Gulsen, O., R.C. Shearman, K.P. Vogel, D.J. Lee, P.S. Baenziger, T.M. Heng-Moss & H. Budak. 2005. Nuclear genome diversity and Relationships among Naturally Occurring Buffalograss genotypes Determined by Sequence-related amplified Polymorphism Markers. HortScience 40: 537-541. [ Links ]
Hale, A.L., M.W. Farnham & M.A. Menz. 2006. Use of PCR-based markers for differentiating broccoli inbreeds. J. Amer Soc. Hort. Sci. 131: 418-423.
[ Links ]
Hao, Q., Z.A. Liu., Q.Y. Shu, R. Zhang, J. De Rick & L.S. Wang. 2008. Studies on Paeonia cultivars and hybrids identification based on SRAP analysis. Hereditas 145: 38-47. [ Links ]
Huang, S.C., C.C. Tsai & C.S. Sheu. 2000. Genetic analysis of Chrysanthemum hybrids based on RAPD molecular markers. Bot. Bull. Acad. Sin. 41: 257-262. [ Links ]
Huchett, B.I. & F.C. Botha. 1995. Stability and potensial use of RAPD markers in asugarcane genealogy. Euphytica 86: 117-125. [ Links ]
Hunt, G.J & R.E. Jr. Page. 1992. Patterns of inheritance with RAPD molecular markers reveal novel types of polymorphism in the honey bee. Theor. Appl. Genet. 85: 15-20. [ Links ]
Lashermes, P., P. Trouslot, F. Anthony, M.C. Combes & A. Charrier. 1996. Genetic diversity for RAPD markers between cultivated and wild accessions of Coffea arabica. Euphytica 87: 59-64. [ Links ]
Lecolier, A., P. Besse, A. Charrier, T.N. Tchakaloff & M. Noirot. 2009. Unravelling the origin of Coffea arabica ‘Bourbon pointu’ from La Reunion: a historical and scientific perspective. Euphytica 168: 1-10. [ Links ]
Li, G. & C.F. Quirus. 2001. Sequence–related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica. Theor. Appl. Genet. 103: 455-461. [ Links ]
Liu, L., G. Liu, Y. Gong, W. Dai, Y. Wang, F. Yu & Y. Ren. 2007. Evaluation of genetic purity of F1 hybrid seeds in cabbage with RAPD, ISSR, SRAP and SSR markers. HortScience 42: 724-727. [ Links ]
Merotto, A., M. Jasieniuk & A.J. Fischer. 2009. Estimating the outcrossing rate of Cyperous difformis using resistance to ALS-inhibiting herbicides and molecular markers. Weed Res. 49: 29-36. [ Links ]
Murray, M.G. & W.F. Thomson. 1980. Rapid isolation of high molecular weight plant DNA. Nucl. Acids. Res. 8: 4321-4326. [ Links ]
Mutlu, N., F.H. Boyaci, M. Gocmen & K. Abak. 2008. Development of SRAP, SRAP-RGA, RAPD andSCAR markers linked with a Fusarium wilt resistance gene in egg plant. Theor. Appl Genet 117: 1303-1312. [ Links ]
Novy, R.G. & N. Vorsa. 1996. Evidence for RAPD heteroduplex formation in cranberry: implications for pedigree and genetic-relatedness studies and a source of co-dominant RAPD markers. Theor. Appl. Genet 92: 840-849. [ Links ]
Smith, J.F., C.C. Burke & W.L. Wagner. 1996. Interspecific hybridization in natural populations of Cyrtandra (Gesneriaceae) on the Hawaiian islands: Evidence from RAPD markers. Pl. Syst. Evol. 200: 61-77. [ Links ]
Sun, S.J., W. Gao, S.Q. Lin, J. Zhu, B.X. Xie & Z.B. Lin. 2006. Analysis of genetic diversity of Ganoderma population with a novel molecular marker SRAP. Appl. Microbiol. Biotechnol. 72: 537-543. [ Links ]
Vandemark, G.J., J.J. Ariss, G.A. Bauchan, R.C. Larsen & T.J. Hughes. 2006. Estimating genetic relationships among historical sources of alfalfa germplasm and selected cultivars with sequence related amplified polymophisms. Euphytica 152: 9-16. [ Links ]
Wilde, J., R. Waugh & W. Powel. 1992. Genetic fingerprinting of Theobroma clones using randomly amplified polymorphic DNA markers. Theor. Appl. Genet. 83: 871-877. [ Links ]
Xuan, C., G. Hailin, X. Dandan & L. Jianxiu. 2008. Identification of Zyosia Hybrids by SRAP Analysis. Molecular Plant Breeding 6: 1233-1238. [ Links ]
Zaefizadeh, M. & M. Goliev. 2009. Diversity and Relationships among Durum wheat Landraces (Subconvars) by SRAP and Phenotypic Marker Polymorphism. Res. J. Biol. Sci. 4: 960-966. [ Links ]
Zhao, W., R. Fang, Y. Yang, J.W. Chung, I.M. Chung & Y.J. Park. 2009. Analysis of genetic relationships of mulberry (Morus L. ) germplasm using sequence-related amplified polymorphism (SRAP) markers. Afr. J. Biotechnol. 8: 2604-2610. [ Links ]
Correspondencia a: Manoj Kumar Mishra, Narayana Suresh, Asha M. Bhat, Nayani Suryaprakash, Saya Satheesh Kumar, Anil Kumar & Jayarama: Central Coffee Research Institute, Coffee Research Station -577117, Chikmagalur- Dist, Karnataka, India; manojmishra.m@gmail.com, sureshccri@gmail.com, ashabhat_bt@yahoo.co.in, nayanirao@gmail.com, sayasath-eesh@gmail.com, anilsirsi@yahoo.com, ccrijaya@gmail.com
Received 09-VII-2010. Corrected 15-XII-2010. Accepted 18-I-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}