










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.2 San José Jun. 2011
Phenotypic plasticity of Vaccinium meridionale (Ericaceae) in wild populations of mountain forests in Colombia
Gustavo A. Ligarreto, Maria del Pilar Patiño & Stanislav V. Magnitskiy
Facultad de Agronomía, Universidad Nacional de Colombia, Ciudad Universitaria, Av. Carrera 30 N° 45-03, Bogotá, Colombia; galigarretom@unal.edu.co, mppatinog@unal.edu.co, svmagnitskiy@unal.edu.co
Dirección para correspondencia
Abstract
Vaccinium meridionale is a promising crop for the Andean region of South America and is currently available only in the wild. Spontaneous populations of this plant are found across the Colombian mountains, but very few published records on this plant morphology are available. A zonification study of V. meridionale was conducted in four principal areas of a low mountain forest of Colombia (Provinces of Boyacá, Cundinamarca, Santander and Nariño) in 2007. A total of 20 populations and 100 plants of V. meridionale were individually characterized and surveyed, using a list of 26 characters of morphological variables (9 quantitative and 17 qualitative characters). Our results indicated that natural populations of V. meridionale might be found in the tropical forest under a highly heterogeneous climate and microclimate conditions, at different mountain regions between 2 357 and 3 168masl. The shrubs of V. meridionale exhibited a high level of intra- population variation in several quantitative (plant height, stem diameter) and qualitative (growth habit, ramification density, presence of anthocyanins in stems) morphological characters, suggesting an environmentally induced phenotypic plasticity. Plant height, stem diameter and foliar density were the most variable morphological traits, with coefficients of variation higher than 50%. However, several quantitative characters of its reproductive potential, such as berry dimensions, rachis length and number of flowers per inflorescence, resulted with low plasticity with coefficients of variation lower than 30.2%, indicating that these characters were genetically determined. The highest correlation coefficients (p<0.05) resulted to be between fruit length and fruit width (0.90), leaf length and leaf width (0.78), plant height and stem diameter (0.60), and inflorescence length and flowers number per inflorescence (0.57). The results suggest that an important genetic resource exists for this species in the wild. Low variation in fruit size, which constitutes a target trait for plant breeders, could be useful for selection of cultivars of V. meridionale. The results of this study could also be applied in conservation programs aimed to protect these diverse populations in the mountain forests of Colombia. Rev. Biol. Trop. 59 (2): 569-583. Epub 2011 June 01.
Key words: plant morphological characters, Vaccinium meridionale, tropical forest, in situ characterization, Andean region, Colombia.
Resumen
Vaccinium meridionale es una planta promisoria para la región Andina de Sudamérica y está disponible actualmente sólo en forma silvestre. Las poblaciones espontáneas de esta planta se encuentran en las montañas colombianas y existen muy pocos reportes publicados respecto a su morfología. Se realizó un estudio de zonificación de V. meridionale en cuatro áreas principales de un bosque de baja montaña de Colombia (Provincias de Boyacá, Cundinamarca, Santander y Nariño) en 2007. Un total de 20 poblaciones y 100 plantas de V. meridionale fueron individualmente caracterizadas usando una lista de 26 descriptores de variables morfológicas (9 caracteres cuantitativos y 17 caracteres cualitativos). Nuestros resultados indicaron que las poblaciones naturales de V. meridionale podrían ser encontradas en el bosque tropical bajo unas condiciones de clima y microclima muy heterogéneas, en regiones de montaña diferentes entre 2 357 y 3 168masl. Los arbustos de V. meridionale presentaron un nivel alto de la variación intrapoblacional en varios caracteres morfológicos cuantitativos (altura de planta, diámetro de tallo) y cualitativos (hábito de crecimiento, densidad de ramificación, presencia de antocianinas en tallos), sugiriendo una plasticidad fenotípica ambientalmente inducida. La altura de planta, el diámetro de tallo y la densidad foliar fueron los rasgos morfológicos más variables, con coeficientes de variación superior al 50%. Sin embargo, varios caracteres cuantitativos de su potencial reproductivo, como dimensiones de baya, longitud de raquis y número de flores por inflorescencia, presentaron plasticidad baja con coeficientes de la variación inferior al 30.2%, indicando que estos caracteres fueron genéticamente determinados. Los coeficientes de correlación más altos (p<0.05) fueron para longitud de fruto y ancho de fruto (0.90), longitud de hoja y ancho de hoja (0.78), altura de planta y diámetro de tallo (0.60), y longitud de inflorescencia y número de flores por inflorescencia (0.57). Los resultados sugieren que en condiciones naturales existe un recurso genético importante para esta especie. La poca variación en el tamaño de fruto, puede ser útil para la selección de cultivares de V. meridionale y se constituye un rasgo objetivo para fitomejoradores. Los resultados de este estudio también pueden ser aplicados en programas de conservación para proteger estas poblaciones diversas en los bosques de montaña de Colombia.
Palabras clave: caracteres morfológicos de planta, Vaccinium meridionale, bosque tropical, in situ caracterización, región Andina, Colombia.
The genus Vaccinium of the family Ericaceae includes about 450 species of trees, shrubs, subshrubs, and hemiepiphytes that are mainly distributed in the tropics of the Old World, Malaysia, and Southeast Asia (Luteyn 2002, Vander Kloet & Dickinson 2009, Vander Kloet & Avery, 2010). According to Luteyn (2002), the neotropics are part of the center of diversity for the family Ericaceae, with most species in Colombia and Ecuador. From the approximately 40 species of Vaccinium present in the neotropics (Luteyn 2002), five native species are reported in Colombia: V. meridionale Swartz, V. floribundum Kunth., V. corymbodendron Dunal., V. euryanthum A C Smith and V. singularis Salinas (Salinas & Betancur 2007). These species occur in Andean cloud forests and the regions of páramo Gentry 1994, Luteyn 2002), with an exception of V. euryanthum, which may be encountered in the lowlands of Guiana Shield (Salinas & Betancur 2007). Among these plants, V. meridionale and V. floribundum are the foremost species that have edible fruits, but these are underutilized and are not domesticated species (Ávila Diaz- Granados et al. 2009 and references therein, Vasco et al. 2009). V. meridionale, known by its local names as agraz, mortiño, Colombian blueberry, Andean blueberry, or Jamaican bilberry, is a wild evergreen shrub in the low mountain forests of Colombia, Ecuador, Peru, Venezuela and Jamaica (Tanner 1982, Gentry 1994, Izco et al. 2007). In mountain areas of Colombia, V. meridionale and V. floribundum may be frequently found growing in associations, in which the latter species is generally of smaller size, higher ramification and smaller fruits when compared to V. meridionale. The species of genus Vaccinium compose the group of clonal plants (Albert et al. 2005) that could be propagated both sexually and vegetatively, with vegetative progenitors well established rapidly through rhizomes or stolons (Wilbur & Luteyn 2008). V. meridionale is known to be partially self-fertile (Tanner 1982), and, in Colombia, the European honey-bee (Apis mellifera) may be important for its crosspollination (Arjona 2001, pers. comm.), however vegetative growth is considered to be a principal strategy of the species propagation in natural habitats (Tanner 1982, McDonald et al. 2003, Ávila Diaz- Granados et al. 2009).
The fruits of V. meridionale have significant economic potential for the international markets due to the high contents of polyphenolic compounds that are known for their beneficial effects for human health including anticancerigenic properties. The average contents of total anthocyanins and total phenols in fruits of V. meridionale were measured 201 cya-3-glu mg/100 g FW and 609 gallic ac. mg/100 g FW, respectively; whereas total antioxidant activity (FRAP) reached 58.1 TE mkM/g (Gaviria et al. 2009). The values comparable to or exceeded those reported for the wild berries of V. myrtillus L. (Giovanelli & Buratti 2009) or V. ovalifolium Smith (Lee et al. 2004). In the Andean region, consumption of fruits of V. meridionale over the centuries maintained importance due to their nutritional and organoleptic qualities However, all activities related to plant usage are limited to recollection of berries in the wild and selling these on the market places or roads nearby to recollection sites as well as artisanal methods of fruit processing for juices, vines and marmalades. Tree damage made through inadequate practices of fruit harvest, tree lodging and branches recollection for charcoal in a low mountain forest (which constitutes a natural habitat of V. meridionale), turns the species currently under risk of genetic erosion (Hernández et al. 2009).
In South America, V. meridionale is currently accessible only in the wild and, in Colombia, a wide variation in the plant morphology is anticipated across the country. According to Ligarreto (2009), the quantitative characteristics of the species in a single Colombian metapopulation, exhibited a broad phenotypic diversity among the studied plants, with coefficients of variation ranging between 18 and 34%. This range implies that phenotypic features are affected by the environment and may be considered as an indicator of genetic variability, especially in variables of broad dispersion of values. Additionally, a wide qualitative variation, with detection of polymorphism in 16 qualitative variables in plants of V. meridionale, was reported (Ligarreto 2009). This type of variation, in general, refers to a high heritability h2 (Hill et al. 1998) that corresponds, according to Van Hintum (1995), to an alleles expression of specific genes, due to which these qualitative variables could be employed as genetic markers.
Colombia is one of the main natural centers where V. meridionale might be encountered in the wild, nevertheless, the distribution of its natural populations in the country is poorly characterized. To our knowledge, the populations of V. meridionale in the North East (Provincy of Santander) and South Pacific (Provincy of Nariño) mountain regions of the country were neither assessed nor characterized, and the knowledge on their possible existence in these areas was based essentially on local reports. Additionally, almost no published records on plant morphology in locations where the spontaneous populations were earlier found (Provinces Cundinamarca and Boycá) are available (Medina & Lobo 2004, Ligarreto 2009). Thus, the goal of this study was to characterize in situ the basic aspects of phenotypic plasticity of V. meridionale within its natural populations in four principal Provinces of Colombia: Cundinamarca, Boyacá, Santander and Nariño; occupied with a tropical mountain forest. The morphological variation degree among wild populations of V. meridionale, constitutes a fundamental question that should be addressed in order to: a) conservation efforts to the diverse populations of V. meridionale in situ and ex situ and b) to obtain plant material for breeding purposes, and promote its cultivation in world agriculture.
Materials and methods
Plant material: Information from previous field studies and reported cases of V. meridionale Swartz from the Colombian mountain areas were studied using herbaria of Instituto de Ciencias Naturales, Universidad Nacional de Colombia as well as publications (Luteyn 2002) and personal communications. These data further was used to systematize the reports and to elaborate maps of possible location of the species in the wild.
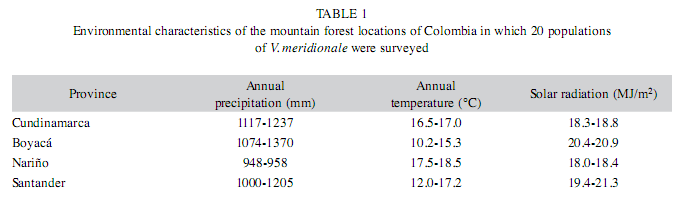
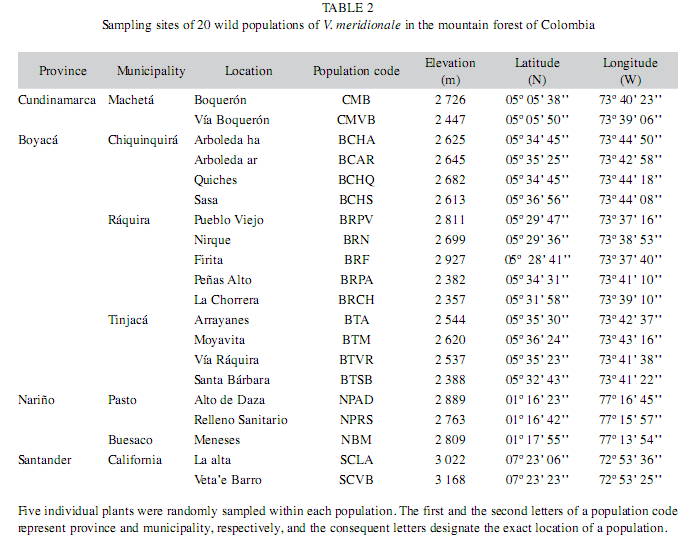
Field survey: According to the preliminary field studies conducted in a pilot zone in Cundinamarca, the presence of V. meridionale populations was tested in the similar habitats through the Provinces, making this study one of the first ones to make a geographic distribution of this species in Colombia. Spontaneous populations of V. meridionale were surveyed in the Provinces of Boyacá, Cundinamarca, Santander and Nariño located in a low mountain area (between 2 100 and 3 200m elevation) in Colombia (Table 1). Each plant was labeled with an identification code, where the first and the second letters represent province and municipality, respectively, the subsequent letters designate the exact location of a population, and a number indicates the plant organism assessed in the population (Table 2). The field studies were conducted during May-June 2007, that corresponded to the dry season, when berries of V. meridionale are usually collected in the mountain areas across the country. The populations were assessed randomly and the sampling scheme was intended to cover the range of precipitation and temperature regimes anticipated for the species (Luteyn 2002), and also taking into account that, in Colombia, V. meridionale is scarcely reported growing at elevations above 3 200m and below 2 300m. The population was considered a group of plants in a reproductive state growing under free solar exposure or in understory. Data were taken on populations located at 5km minimal distance one from another. In total 100 plants of V. meridionale were characterized individually in 20 spontaneous populations located in the mountain forests of four Provinces. For a given population, each morphological character was accessed in 5 randomly selected plants to evaluate intra-population variability and raw data were then averaged for the statistical analyses.
Soil samples that included the whole topsoil were collected for chemical analysis in all surveyed sites and soil pH (soil:water 1:1) was measured in the Laboratory for Soil Analyses of the Faculty of Agronomy, National University of Colombia, Bogotá campus. The climate data were obtained from the meteorological stations of IDEAM, Colombia.
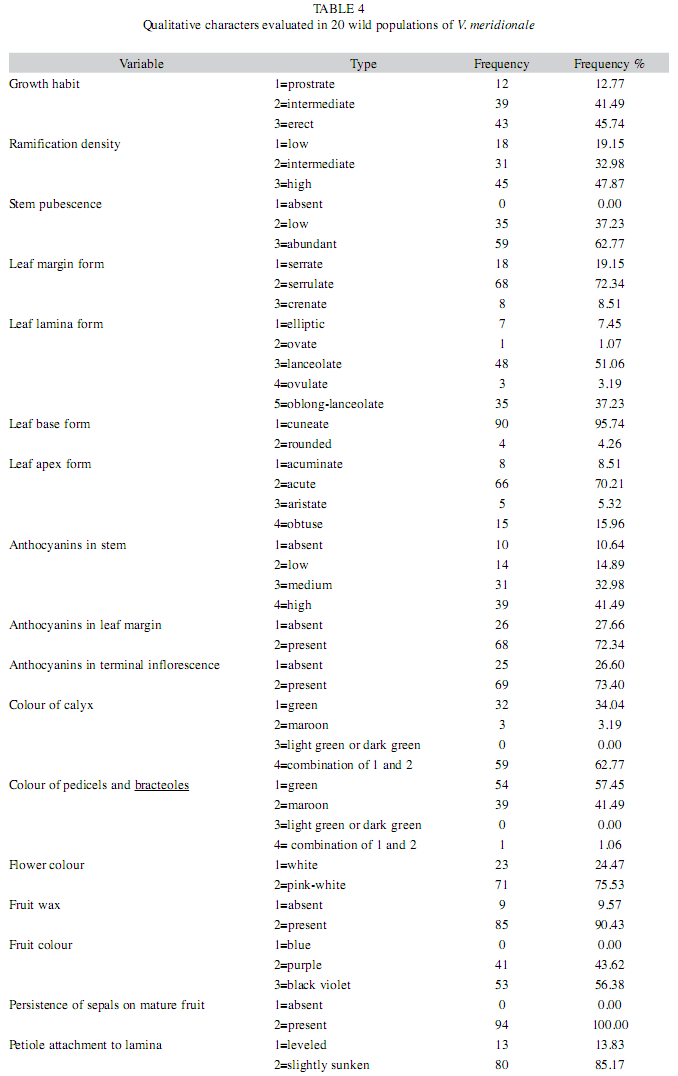
Morphological measurements: Description of plant morphology in situ was performed using a list of variables consisting of 9 quantitative and 17 qualitative characters according to Lopera (2005). Quantitative variables, excluding plant height, were measured with a digital caliper in mm to two decimal places. Plant height (PH) was measured as a distance between the soil level and the highest point of a principal orthotropic shoot. For measurements of stem diameter (SD), the mid-section of a principal stem was evaluated in each plant. In order to measure leaf density (LD), number of leaves was counted on 10cm segments of a single branch taken from the mid-part of the plant. Equally, leaf length (LL) and leaf width (LW) were measured in the mid-section of fully expanded leaves. For quantitative fruit variables, diameter of mature fruits was measured in the longest (fruit length, FL) and the shortest (fruit width, FW) sections of the fruits. Number of flowers per inflorescence (FI) and inflorescence length (IL) were counted on a terminal inflorescence located at the same branch, where the fruit characters were evaluated. The stem, leaf, and floral qualitative characters were evaluated subjectively as following: growth habit (1=prostrate, 2=intermediate, 3=erect), ramification density (1=low, no branches; 2=intermediate, 1 or 2 orders of ramification; 3=high, 3 or 4 orders of ramification), stem pubescence (1=absent, 2=low, 3=abundant), form of leaf margin on fully expanded leaves (1=serrate, number of points: >12/cm; 2=serrulate, number of points: 12-6/cm; 3=crenate, number of points: <6/ cm), form of leaf lamina (1=elliptic, 2=ovate, 3=lanceolate, 4=ovulate, 5=oblong-lanceolate), form of leaf base (1=cuneate, 2=rounded), form of leaf apex (1=acuminate, 2=acute, 3=aristate, 4=obtuse), anthocyanins in stem (1=absent, 2=low, 3=medium, 4=high), anthocyanins in leaf margin (1=absent, green margins; 2=present, maroon margins), anthocyanins in terminal inflorescence (1=absent, 2=present), colour of calyx (1=green, 2=maroon, 3=light green or dark green, 4=combination of 1 and 2), colour of pedicels and bracteoles (1=green, 2=maroon, 3=light green or dark green, 4= combination of 1 and 2), flower colour (1=white, 2=pink-white), fruit wax (1=absent, 2=present), fruit colour (1=blue, 2=purple, 3=black violet), persistence of sepals on mature fruit (1=absent, 2=present), petiole attachment to lamina (1=leveled, 2=slightly sunken).
Summary statistics were used to describe the variation in morphology of V. meridionale, and a correlation analysis was conducted on the full set of quantitative morphological traits. Differences were considered significant at p<0.05 level. A Principal Component Analysis (PCA) was undertaken on quantitative characters using SPAD 4.5 program (Pagès 2004). Since distributions of all quantitative characters were approximately normal, no data transformation for PCA was required.
Results
Natural environments of the populations: Wild populations of V. meridionale were found in four mountain forest regions of Colombia, within the elevations 2 357 and 3 168m (Table 2). Based on the preliminary zonification studies conducted for this species in the Provincy of Cundinamarca, the wild populations of V. meridionale in Boyacá, Santander and Nariño were identified and characterized. In general, the populations were found in locations with slopes of 3 to 50%, and vegetation typical of life zones of lower dry montane and lower wet montane forests, that were rich in arrayán (Myrcianthes leucoxyla (Ortega) McVaugh), laurel (Myrica parvifolia Benth.), Ericaceae of genera Pernettya and Cavendishia, epiphytes (Bromeleaceae), and hemiepiphytes (Clusiaceae).
Most of the plants of V. meridionale in each population were found growing under free solar exposure; also, the plants were observed as a co-dominant species growing over the litter coverage and associated with oak (Quercus humboldtii Bonpl.) and pine patula (Pinus patula Schiede ex Schltdl. & Cham.), making this last association very common in all studied Provinces. Other frequent species in forest locations of V. meridionale were Dendropanax macrophyllum Cuatrec and Bombacopsis patinoi Dugand & Robyns (Nariño) and Alnus acuminata Kunth. (Cundinamarca, Boyacá and Santander). For Nariño locations, the third typical species were Ossaea macrophylla (Bentham) Cogniaux as well as various Rubiaceae species, especially Elaeagia utilis (Goudot) Wedd. and Rubus spp. Ground vegetation in all studied sites consisted mainly of mosses (Sphagnum spp.) and ferns, having total percentage coverage of more than 80%.
The climate factors that apparently affected the distribution of populations V. meridionale in the wild were temperature, precipitation, and radiation. Climate conditions, where the populations were found across the studied zones, highly differed in mean daily temperatures (8.4-23.6°C) and annual precipitation rates that ranged from 948mm (very dry areas) up to 1 237mm (very humid areas); the variation in solar radiation was relatively low, between 18.0 and 21.3MJ/m2 (Table 1).
A constant condition was the soil acidity, where pH ranging from 3.8 to 5.3, with pH 4.6 on average. All soils of the study zones presented good drainage and udic moisture regime. Plants were found growing in soils ranging from large A horizons that exceeded 50cm to very superficial soils, e.g. directly on the parental rock material, as in a particular case of populations BRPV and BRF located in Ráquira, Boyacá.
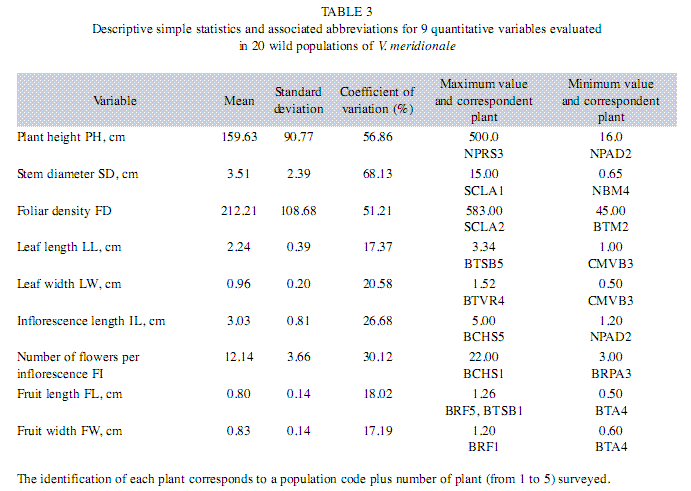
Quantitative characterization of species morphology in populations: Plant height, stem diameter and foliar density were the most variable quantitative morphological traits, with coefficients of variation higher than 50% (Table 3). The mean plant height of 159.63cm was characterized with a standard deviation of 90.77cm indicating high degree of data dispersion (Table 3). The stem diameter presented a mean value of 3.51cm with a standard deviation of 2.39cm, apparently affected by the presence of very dissimilar data, as may be seen in Table 3. High data variation was equally observed for foliar density, with a standard deviation of 108.68cm with respect to a mean value of 212.21cm (Table 3).
Leaf and fruit size characters, namely length and width, had narrow amplitude of variation, with coefficients of variation lower than 21%, indicating that these were relatively homogeneous characters of low phenotypic plasticity in all populations (Table 3). The leaves were on average 2.24cm long and 0.96cm wide with standard deviations of 0.39 and 0.20cm, respectively, while fruit length and fruit width presented mean values of 0.80 and 0.83cm with standard deviations of 0.18 and 0.17cm, respectively. Although the behavior of fruit size characters was of low plasticity in the studied metapopulation, in some habitats of the highest elevations, plants had a tendency to produce leaves and fruits of the smallest sizes. These patterns were also observed with increasing number of fruits per inflorescence, suggesting a competition among the fruits; however, on rare occasions fruit diameters were less than 6mm. Inflorescence length and number of flowers per inflorescence were characterized as more heterogeneous, with coefficients of variation of 26.68% and 30.12%, respectively (Table 3).
Among the characters observed, the highest correlation coefficients (p<0.05) were between fruit length and fruit width (0.90), leaf length and leaf width (0.78), plant height and stem diameter (0.60), and inflorescence length and number of flowers per inflorescence (0.57). No relationships between inflorescence length and fruit length or fruit width were found. The weak negative correlations were observed between foliar density, on one side, and leaf length (-0.20) or leaf width (-0.21), on the other side, indicating that, in a ramet with smaller number of leaves, these tended to be larger. Stem diameter had a positive correlation with foliar density (0.36). The leaf size characteristics were correlated with fruit size and inflorescence size, such as the positive correlations were established between leaf length, on one side, and fruit length (0.29), fruit width (0.38), or inflorescence length (0.20) on the other side. Leaf width had the positive correlations with fruit length (0.20), fruit width (0.28) or inflorescence length (0.21).
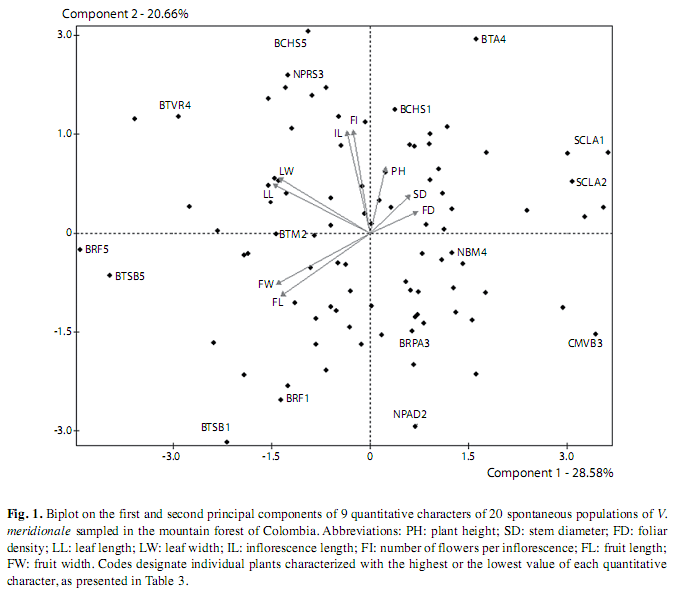
The PCA revealed that fruit size variables, namely fruit length and fruit width, contributed to the first component (F1) that accounted for 28.58% total variance (Fig. 1). Number of flowers per inflorescence and inflorescence length were the major loading characters to the second component (F2) that explained 20.66% total variance. It may be noticed that the plants with extreme (maximum or minimum) states of quantitative characters were observed within the same area as well as in different geographical locations (Fig. 1, Table 3). For example, the plants located in Pasto, Nariño presented both the highest (5.0m, NPRS3 that exceeded the height ranges typical for the shrubs of Vaccinioideae (Gentry 1994, Lens et al. 2004) and the lowest height (0.16m, NPAD2) within all populations evaluated. The latest individual also had the smallest inflorescence length (1.2cm), while the longest inflorescence (5.0cm) was observed on a plant in Chiquinquirá, Boyacá (BCHS5), located in the positive quadrant of F2, for which the variable of inflorescence length has the highest contribution (Fig. 1). The highest leaf length (3.34cm) was measured in a plant from Santa Bárbara, Boyacá (BTSB5) located in the negative quadrant of F1, whereas the smallest leaf length (1.0cm) was recorded for a plant CMVB3 from Vía Boquerón, Boyacá (Table 3), located in the opposite quadrant of the plot (Fig. 1). In general, the PCA revealed a high dispersion of characters without correlation with their location, which indicates a genetic diversity of plants within the metapopulation. Equally, the PCA demonstrated a high dispersion in quantitative characters within some populations, showing a high intrapopulational morphological variation in plants of V. meridionale.
Qualitative characterization of species morphology in populations: A qualitative description of V. meridionale in wild populations illustrated that plant growth habit varied among intermediate and erect, with similar frequencies of 41.9% and 45.74%, respectively. Ramification was predominantly dense with 47.87% individual plants located within this category; however, there were 32.98% plants of intermediate density (Table 4).
The variables of leaf form, such as forms of margin, apex, lamina and base had lower ranges of dispersion than the above-mentioned characters, so that the predominant leaf forms may be distinguished. In the 51.06% of studied plants, leaf lamina form was lanceolate followed by 37.23% oblong-lanceolate. Serrulate leaf margins constituted a distinctive feature of V. meridionale plants having a frequency of 72.34% in all populations. The majority of plants presented leaves with cuneate foliar base (95.74%) and acute apex (70.21%) (Table 4).
Anthocyanin contents in the mid-section of plant stems were mostly high (32.98%), medium (41.49%), low (14.89%) and only 10.64% of plants completely lack this product. Accumulation of anthocyanins in leaf margins and inflorescences was detected in more than 72% of evaluated plants. Calyx colour varied among green, maroon, and a combination of these, showing maroon on the margins and green on the central part of the calyces. The combination of green and maroon was the most frequent colour (62.77%) followed by the one with complete green calyx (34.04%). Colour of pedicels was predominantly green (57.45%), but maroon pedicels were also observed (41.49%). Colour of bell-shaped flowers was characteristically pink- white to almost fuchsia (75.53%), whereas some flowers were completely white (24.47%). Pubescence in mid-sections of stems was present for 62.77% plants and absent for 37.23% plants assessed in the study (Table 4).
Fruit colour was purple (43.62%) or black violet (56.38%), while no fruits were found with a the characteristically blue colour of V. corymbosum or V. myrtillus, that may vary from blue to almost black. However, some mature berries of V. meridionale had white and light rose colour, which may be the possible mutants of structural or regulatory genes involved in anthocyanin synthesis, as it was shown for V. myrtillus (Jaakola et al. 2002).
Some characters of fruit morphology, such as wax presence on fruit surface (90.43%) or the form of petiole insertion, and for which the major form was slightly sunken (85.17%), had very low variation. Persistence of sepals was a constant feature for mature fruits in all evaluated populations, with 100% plants grouped in the category "present" (Table 4).
Discussion
Our results indicated that natural populations of V. meridionale might be found under highly heterogeneous climate and microclimate conditions in a tropical mountain forest, in different geographic areas of Colombia. The location of wild populations of V. meridionale in the studied zones corresponded to the habitats previously described by Luteyn (2002) who mentioned preference of neotropical Vaccinium spp. to humid and undisturbed mountain habitats. The plants of V. meridionale were found growing under partial shade or free solar exposure as well as in canopy gaps suggesting high rates of adaptability to different microenvironments including those of solar radiation within a same study area.
Plant height, stem diameter and foliar density were highly variable morphological traits (Table 3), and, while plant height is a subject of phenotypic variation in wild clones of Vaccinium spp. (Gustavsson 2001), in our study, this variable might not provide a consistent comparison across the plants because their ages were unknown. High variability of the above mentioned descriptors may be due to the fact that, in wild populations, there were plants of various age stages, reproduction strategies, light, water, and nutrient regimes - all these differences, when combined with occasional distortion of plant habitats by tree lodging, generated populations highly heterogeneous in plant height, stem diameter and foliar density. On the contrary, a low variation in fruit length and fruit width of V. meridionale (Table 3), indicates a low phenotypic plasticity of these characters and could be attributed to a low environmental effect over the fruit size that, in turn, implies that characterization of fruit morphology in situ could be used to estimate the levels of genetic diversity (Vargas & Blanco 2000). Fruit size in Vaccinium sp. is known to be affected both by genetic and environmental factors; among the latest, pollination (Eck 1988) and irrigation regimes (Hicklenton 2000, Mingeau 2001); a reduction in leaf size with increasing altitude, which is commonly observed in deciduous Vaccinium spp., that could be due to withstanding nitrogen limitation and low temperatures (Woodward 1986). Inflorescence length and number of flowers per inflorescence might also serve as indicators of plant adaptation to certain environments, as in the case of V. corymbosum, for which the flower number was reduced under moderate water stress (Mingeau 2001). These characters describe a reproductive state of the species; however, as in case of fruit or leaf size, low or moderate variation in inflorescence length and number of flowers per inflorescence (CV<30.2%, Table 3) indicates their low phenotypic plasticity.
In general, different environments that represented microclimates, influenced the variation of quantitative characters. Our findings are consistent with the pioneer studies of Medina & Lobo (2004) of wild populations of V. meridionale in the Provincy of Antioquia, Colombia, who revealed that plant height and stem diameter were highly variable morphological characters. The positive correlation established between plant height and stem diameter might be explained by plant age. Additionally, in Ericaceae, an environment might have a significant effect over the stem diameter, such as the trends in decreasing stem diameter with increasing altitude of plant habitat (Woodward 1986, Merev & Yavuz 2000) or microclimate changes including low availability of soil water (Mingeau 2001).
The PCA revealed that fruit length, fruit width, number of flowers per inflorescence, and inflorescence length were the main loading characters that accounted for a total variance (Fig. 1). These characters, especially fruit size, are useful in characterization of sexual reproductive potential of V. meridionale, taking into account that the largest seeds (up to 1.9mm length and 1.4mm width) have the highest germination rates, and could be obtained from the largest fruits (Valencia & Ramírez 1993). The latest data may have an importance for sexual recruitment of the species in the wild, however, the rates of seedling establishment in natural habitats, in case of V. meridionale, are unknown, and, in case of other Vaccinium spp. (Eriksson 1989), might be negligible. However, it is worth mentioning that propagation by seeds, as well as somatic mutagenesis and initial genetic diversity in a given population, are the main factors that increase a genetic variation in clonal populations of Vaccinium spp. (Persson & Gustavsson 2001, Albert et al. 2005). The results of the PCA demonstrated a genetic diversity of V. meridionale within the metapopulation as well as an intrapopulational variation in plant morphology for some populations.
The morphological traits that, in the majority of plants, respond to the environmental stimuli changes are elongation of stem and foliar growth (Fordyce 2006). Thus, within a given population, the tallest plants of V. meridionale were frequently found in partial shade, while those of arbustive prostrate growth were typical of open sites; as shown for other Ericaceae (Dawson & Heenan 2004). Additionally, V. meridionale is a plant of clonal propagation through stolons and rhizomes (McDonald et al. 2003), and variation in form of growth in clonal plants also represents a particular case of plasticity (Navas & Garnier 2002) observable in other Vaccinium spp.; as for example, a reduction in layering stem growth under intense radiation (Moola & Mallik 1998) or during fruit production (Elle 1996, Gustavsson 2001). The fact that plants were studied in highly heterogeneous environments makes it feasible to conduct further studies on the contribution of phenotypic plasticity to morphological variability of V. meridionale.
Several qualitative characters of species morphology were highly variable, which at least partially, could be attributed to phenotypic plasticity. Phenotypic plasticity related to changes in plant height and shape may be directed, in particularly, to optimize light use efficiency in natural populations (Dawson & Heenan 2004, Wang et al. 2006). This could explain a high variability in height (plant height, stem diameter) and shape (growth habit, ramification density) characters of V. meridionale, and, indirectly, indicate that plants in the studied metapopulation experienced a certain environmental pressure. At the same time, some qualitative characters of fruit or leaf morphology had very low (wax presence on fruit surface, form of petiole insertion) or null (persistence of sepals on mature fruits) variation demonstrating that these characters were genetically determined.
The variables that characterize colour of stems, flowers and leaves in Vaccinium depend on anthocyanins contents in these plant organs (Eck 1988). Phenotypic variation in fruit colour, as the one shown in our study, is readily apparent among wild clones of lowbush blueberry (Kalt et al. 2001) and attributed to tissue levels of anthocyanins and other flavonoid compounds (Eck 1988). Synthesis of anthocyanins in leaves and fruits of Vaccinium is developmentally regulated (Witzell et al. 2003) and could be highly variable when stimulated by numerous environmental factors, including light, UV radiation, fungal infection, wounding, drought or low nitrogen status (Jaakola et al. 2004, Witzell & Shevtsova 2004). Direct sunlight activates the synthesis of anthocyanins in leaves, fruits and flowers of V. myrtillus (Jaakola et al. 2004 and references therein), so that tissue concentrations of anthocyanins tend to increase conferring a higher desiccation tolerance and photoprotection of cells, especially at low temperatures (Tahkokorpi et al. 2007), the normal conditions with increasing altitudes in plant habitat. While the current relationship may not be always recognizable (Rieger et al. 2008), in our study, the highest levels of anthocyanins in leaves, fruits and flowers were scored mostly on plants growing in the Provinces of Boyacá and Santander that are characterized with the highest solar radiation levels (Table 1). Flowers colour variation among the populations might also have an effect on the reproductive traits and genetic diversity. Thus, in temperate climates, plants of lingonberry with more pinkish corollas began flowering later than plants with white flowers (Gustavsson 2001); this feature, under dry-rainy seasonality conditions in a tropical mountain forest, might be used to synchronize flowering and fructification with cycles of pollinators and frugivores, respectively (Hilty 1980).
In general, our studies revealed a wide morphological variation in the metapopulation of V. meridionale across the study areas. Several quantitative (plant height, stem diameter) and qualitative (growth habit, ramification density, presence of anthocyanins in stem) descriptors of the species morphology were highly influenced by the environment. Our results suggested that an important genetic resource exists for V. meridionale in the wild that could provide a basis for introduction of this species into world agriculture. Thus, low variation in fruit size (CV<18.02%), which constitutes a target trait for plant breeders, might be useful for the development of breeding programs aiming to the selection of cultivars of V. meridionale. Additionally, the studies on genetic diversity in V. meridionale at the molecular level would help to implement a conservation strategy for this species. Alteration of the species habitat results in reduction of population size that, in turn, could diminish a degree the morphological variation within and among the populations, the process that would further determine ecotypes formation of V. meridionale. While some habitats of natural populations of V. meridionale in Santander and, especially, Nariño, remain undisturbed and practically inaccessible in the near future, fragmentation of a mountain forest, and reduction in populations size are increasing in other regions, such as Cundinamarca and Boyacá, so that the development of strategies to prevent genetic erosion of V. meridionale in its natural habitats should be addressed.
Acknowledgments
This research was conducted under the project "Estandarización de métodos de propagación y escalamiento de producción de plántulas de agraz (Vaccinium meridionale Swartz)" funded by The Ministry of Agriculture and Rural Development of Colombia and The National University of Colombia, Bogotá campus, Colombia. The authors express their gratitude to César Pacheco Chaparro (Annonaceae Research Project, Agronomy Faculty, National University of Colombia) for the help with data analyses and Alia Rodríguez (Agronomy Faculty, National University of Colombia) for review of the manuscript and help with the English translation.
References
Albert, T., O. Raspe & A.L. Jacquemart. 2005. Diversity and spatial structure of clones in Vaccinium uliginosum populations. Can. J. Bot. 83: 211-218. [ Links ]
Ávila Díaz-Granados, R.A., O.J. Orozco Silva, G. Ligarreto Moreno, S. Magnitskiy & A. Rodríguez. 2009. Influence of mycorrhizal fungi on the rooting of stem and stolon cuttings of the Colombian blueberry (Vaccinium meridionale Swartz). Inter. J. Fruit Sci. 9: 372-384. [ Links ]
Dawson, M.I. & P.B. Heenan. 2004. Morphological variation of the Leucopogon fraseri complex (Ericaceae: Styphelieae) in New Zealand, and recognition of a new species, L. nanum. New Zeal. J. Bot. 42: 537-564. [ Links ]
Eck, P. 1988. Blueberry science. Rutgers University, New Brunswick, New Jersey, USA. [ Links ]
Elle, E. 1996. Reproductive trade-offs in genetically distinct clones of Vaccinium macrocarpon, the American cranberry. Oecologia 107: 61-70. [ Links ]
Errikson, O. 1989. Seedling dynamics and life histories in clonal plants. Oikos 55: 231-238. [ Links ]
Fordyce, J.A. 2006. The evolutionary consequences of ecological interactions mediated through phenotypic plasticity. J. Exp. Biol. 209: 2377-2383. [ Links ]
Gaviria, C.A., N. Ochoa, C.I. Sánchez, C.I. Medina, M. Lobo, P. Galeano, A.J. Mosquera, A. Tamayo, Y. Lopera & B. Rojano. 2009. Actividad antioxidante e inhibición de la peroxidación lipídica de extractos de frutos de mortiño (Vaccinium meridionale SW). Boletín Latinoam. Caribe Plantas Medic. Aromát. 8: 519-528. [ Links ]
Gentry, A.H. 1996. A field guide to the families and genera of woody plants of northwest South America (Colombia, Ecuador, Peru) with supplementary notes on herbaceous taxa. The University of Chicago, Chicago, USA. [ Links ]
Giovanelli, G. & S. Buratti. 2009. Comparison of polyphenolic composition and antioxidant activity of wild Italian blueberries and some cultivated varieties. Food Chem. 112: 903-908. [ Links ]
Gustavsson, B.A. 2001. Genetic variation in horticulturally important traits of fifteen wild lingonberry Vaccinium vitis-idaea L. populations. Euphytica 120: 173-182. [ Links ]
Hernández, M.I., M. Lobo, C.I. Medina, J.R. Cartagena & O.A. Delgado. 2009. Comportamiento de la germinación y categorización de la latencia en semillas de mortiño (Vaccinium meridionale Swartz). Agron. Colomb. 27: 15-23. [ Links ]
Hicklenton, P.R., J.Y. Reekie & R.J. Gordon. 2000. Physiological and morphological traits of lowbush blueberry (Vaccinium angustifolium Ait.) plants in relation to posttransplant conditions and water availability. Can. J. Plant Sci. 80: 861-867. [ Links ]
Hill, J., H.C. Becker & P.M.A. Tigerstedt. 1998. Quantitative and ecological aspects of plant breeding. Chapman and Hall, London, United Kingdom. [ Links ]
Hilty, S.L. 1980. Flowering and fruiting periodicity in a premontane rain forest in Pacific Colombia. Biotropica 12: 292-306. [ Links ]
Izco, J., Í. Pulgar, Z. Aguirre & F. Santin 2007. Estudio florístico de los páramos de pajonal meridionales de Ecuador. Rev. Peru. Biol. 14: 237-246. [ Links ]
Jaakola, L., K. Määttä, A.M. Pirttilä, R. Törrönen, S. Kärenlampi & A. Hohtola. 2002. Expression of genes involved in anthocyanin biosynthesis in relation to anthocyanin, proanthocyanidin, and flavonol levels during bilberry fruit development. Plant Physiol. 130: 729-739. [ Links ]
Jaakola, L., K. Määttä-Riihinen, S. Kärenlampi & A. Hohtola. 2004. Activation of flavonoid biosynthesis by solar radiation in bilberry (Vaccinium myrtillus L.) leaves. Planta 218: 721-728. [ Links ]
Kalt, W., D.A.J. Ryan, J.C. Duy, R.L. Prior, M.K. Ehlenfeldt & S.P. Vander Kloet. 2001. Interspecific variation in anthocyanins, phenolics, and antioxidant capacity among genotypes of highbush and lowbush blueberries (Vaccinium Section Cyanococcus). J. Agric. Food Chem. 49: 4761-4767. [ Links ]
Lee, J., C.E. Finn & R.E. Wrolstad. 2004. Anthocyanin pigment and total phenolic content of three Vaccinium species native to the Pacific Northwest of North America. HortScience 39: 959-964. [ Links ]
Lens, F., J.L. Luteyn, E. Smets & S. Jansen. 2004. Ecological trends in the wood anatomy of Vaccinioideae (Ericaceae s.l.). Flora 199: 309-319. [ Links ]
Ligarreto, G.A. 2009. Selección de poblaciones in situ de agraz (Vaccinium meridionale Swartz) por atributos fenotípicos reproductivos. Pp. 69 XI Congreso de Asociación Colombiana de Fitomejoramiento y Producción de Cultivos. Octubre 28- 30 2009, Palmira, Colombia. [ Links ]
Lopera, S.A. 2005. Colecta, conservación y caracterización de diversas poblaciones de Vaccinium meriodionale (mortiño) presentes en los bosques altoandinos de la jurisdicción de Corantioquia, para promover su utilización sostenible. Tesis, Universidad Católica de Oriente, Rionegro, Antioquia, Colombia. [ Links ]
Luteyn, J. 2002. Diversity, adaptation, and endemism in neotropical Ericaceae: Biogeographical patterns in the Vaccinieae. Botan. Rev. 68: 55-87. [ Links ]
McDonald, M.A., A. Hofny-Collins, J.R.T. Healey & C.R. Goodland. 2003. Evaluation of trees indigenous to the montane forest of the Blue Mountains, Jamaica for reforestation and agroforestry. Forest Ecol. Manag. 175: 379-401. [ Links ]
Medina, C.I. & M. Lobo. 2004. Informe final técnico del proyecto "Colecta, conservación y caracterización de diversas poblaciones de Vaccinium meridionale (mortiño), presentes en los bosques altoandinos de la jurisdicción de Corantioquia para promover su utilización sostenible. I Fase". Rionegro, Antioquia, Colombia. [ Links ]
Merev, N. & H. Yavuz. 2000. Ecological wood anatomy of Turkish Rhododendron L. (Ericaceae). Intraspecific variation. Turk. J. Bot. 24: 227-237. [ Links ]
Mingeau, M., C. Perrier & T. Améglio. 2001. Evidence of drought-sensitive periods from flowering to maturity on highbush blueberry. Sci. Hortic. 89: 23-40. [ Links ]
Moola, F.M. & A.U. Mallik. 1998. Morphological plasticity and regeneration strategies of velvet leaf blueberry (Vaccinium myrtilloides Michx) following canopy disturbance in boreal mixedwood forests. Forest Ecol. Manag. 111: 35-50. [ Links ]
Navas, M.L. & E. Garnier. 2002. Plasticity of whole plant and leaf traits in Rubia peregrina in response to light, nutrient and water availability. Acta Oecol. 23: 375-383. [ Links ]
Pagès, J. 2004. Analyse factorielle de donnees mixtes: principe et exemple d’application. Laboratoire de mathématiques appliquées. Agrocampus, Rennes cedex, France. [ Links ]
Persson, H.A. & B.A. Gustavsson. 2001. The extent of clonality and genetic diversity in lingonberry (Vaccinium vitis-idaea L.) revealed by RAPDs and leaf-shape analysis. Molec. Ecol. 10: 1385-1397. [ Links ]
Rieger, G., M. Müller, H. Guttenberger & F. Bucar. 2008. Influence of altitudinal variation on the content of phenolic compounds in wild populations of Calluna vulgaris, Sambucus nigra, and Vaccinium myrtillus. J. Agric. Food Chem. 56: 9080-9086. [ Links ]
Salinas, R.N. & J. Betancur 2007. Novedades taxonómicas de las Ericaceaes del suroccidente de Colombia. Caldasia 29: 51-58. [ Links ]
Tahkokorpi, M., K. Taulavuori, K. Laine & E. Taulavuori. 2007. After-effects of drought-related winter stress in previous and current year stems of Vaccinium myrtillus L. Environ. Exp. Bot. 61: 85-93. [ Links ]
Tanner, E.V.J. 1982. Species diversity and reproductive mechanisms in Jamaican trees. Biol. J. Linn. Soc. 18: 263-278. [ Links ]
Valencia, M.L.C. de & F. Ramírez. 1993. Notas sobre la morfología, anatomía y germinación del Agraz (Vaccinium meridionale Swartz.). Agron. Colomb. 10: 151-159. [ Links ]
Vasco, C., K. Riihinen, J. Ruales & A. Kamal-Eldin. 2009. Chemical composition and phenolic compound profile of mortiño (Vaccinium floribundum Kunth). J. Agric. Food Chem. 57: 8274-8281. [ Links ]
Vander Kloet, S.P. & T.A. Dickinson. 2009. A subgeneric classification of the genus Vaccinium and the metamorphosis of V. section Bracteata Nakai: more terrestrial and less epiphytic in habit, more continental and less insular in distribution. J. Plant Res. 122: 253-268. [ Links ]
Vander Kloet, S.P. & T.S. Avery. 2010. Vaccinium on the edge. Edinburgh J. Bot. 67: 7-24. [ Links ]
Van Hintum, T.J.L. 1995. Hierachical approaches to the analisis of genetic diversity in crop plants, p. 23-34. In T.J.L. Van Hintum, A.H.D. Brown, C. Spillane & T. Hodgkin (eds.). Core collections of plant genetic resources. John Wiley and Sons, New York, USA. [ Links ]
Vargas, A. & D.A. Blanco. 2000. Fruit characterization of Cocos nuficera L. (Arecaceae) cultivars from the Pacific coast of Costa Rica and the Philippines. Genet. Resour. Crop Evol. 47: 483-487. [ Links ]
Wang, W., S.B. Franklin, Y. Ren & J.R. Ouellette. 2006. Growth of bamboo Fargesia qinlingensis and regeneration of trees in a mixed hardwood-conifer forest in the Qinling Mountains, China. Forest Ecol. Manag. 234: 107-115. [ Links ]
Wilbur, R.L. & J.L. Luteyn. 2008. A synopsis of the mexican and central American species of Vaccinium (Ericaceae). J. Bot. Res. Inst. Texas 2: 207-241. [ Links ]
Witzell, J., R. Gref & T. Näsholm. 2003. Plant-part specific and temporal variation in phenolic compounds of boreal bilberry (Vaccinium myrtillus) plants. Biochem. Systemat. Ecol. 31: 115-127. [ Links ]
Witzell, J. & A. Shevtsova. 2004. Nitrogen-induced changes in phenolics of Vaccinium myrtillus – Implications for interaction with a parasitic fungus. J. Chem. Ecol. 30: 1937-1956. [ Links ]
Woodward, F.I. 1986. Ecophysiological studies on the shrub Vaccinium myrtillus L. taken from a wide altitudinal range. Oecologia 70: 580-586. [ Links ]
Correspondencia a: Gustavo A. Ligarreto, Maria del Pilar Patiño & Stanislav V. Magnitskiy. Facultad de Agronomía, Universidad Nacional de Colombia, Ciudad Universitaria, Av. Carrera 30 N° 45-03, Bogotá, Colombia; galigarretom@unal.edu.co, mppatinog@unal.edu.co, svmagnitskiy@unal.edu.co
Received 18-V-2010. Corrected 15-XII-2010. Accepted 17-I-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}