










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.2 San José Jun. 2011
Ultraestructura de la epidermis foliar de Zeugites (Poaceae: Panicoideae)
Ana María Soriano1, Teresa Terrazas2 & Patricia Dávila3
1. Red de Biodiversidad y Sistemática, Instituto de Ecología, A.C. Carretera antigua a Coatepec 351, El Haya, Xalapa 91070, Veracruz, México; ana.soriano@inecol.edu.mx2. Departamento de Botánica, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado Postal 70-367, 04510, Distrito Federal, México; tterrazas@ibiologia.unam.mx
3. Unidad de Biología Tecnología y Prototipos (UBIPRO), Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México. Avenida de los Barrios No. 1, Los Reyes Iztacala, 54090 Tlalnepantla, Estado de México, México; pdavilaa@servidor.unam.mx
Dirección para correspondencia
Abstract
Key words: key, leaf blade, epidermis, macrohairs, Poaceae, Paniceae, prickles, ultrastructure, Zeugites.
Resumen
El género Zeugites incluye once especies de pastos con distribución neotropical, en México, el Caribe, Centro y Sudamérica. En este trabajo se describe la epidermis foliar de las once especies, se utilizaron técnicas de microscopía electrónica de barrido y revisaron al menos tres especímenes de cada especie, algunas muestras proceden de ejemplares de herbario y otras fueron recolectadas por la primera autora. Se presenta una clave y descripciones para las especies del género, en las cuales se resaltan las características epidérmicas distintivas. Los caracteres que resultaron informativos, desde el punto de vista taxonómico fueron: la presencia o ausencia de aguijones y macropelos, la forma de las células cortas intercostales y la forma de los cuerpos de sílice. Con base en estas características, las especies de Zeugites se pueden separar en tres grupos: (1) especies que no presentan aguijones (Z. americana, Z. mexicana, Z. pringlei, Z. munroana y Z. sagittata) y que con excepción de Z. pringlei tampoco presentan macropelos; (2) especies que presentan sólo aguijones (Z. latifolia y Z. smilacifolia); y (3) especies que presentan aguijones y macropelos (Z. capillaris, Z. hackelii, Z pittieri y Z. sylvatica). Las características morfológicas de la epidermis foliar, sustentan la relación de las tribus Centotheceae y Paniceae.
Palabras clave: clave de identificación, epidermis de la lámina foliar, macropelos, Poaceae, Paniceae, aguijones, ultraestructura, Zeugites.
Diversos autores han resaltado la importancia de los caracteres de la epidermis en la taxonomía de las gramíneas (Prat 1932, 1936, Tateoka et al. 1959, Metcalfe 1960, Jacques-Felix 1962, Barkworth 1981, Renvoize 1986, Hilu 1984, Dávila & Clark 1990). En particular, destaca el trabajo de Dubé & Morisset (1996) quienes afirman que el estudio de la anatomía de la epidermis abaxial es una importante fuente de caracteres diagnósticos, porque a diferencia de lo que ocurre en la mayoría de las angiospermas, la epidermis de gramíneas es compleja y presenta varios tipos de células, mientras otros como Aiken & Lefkovitch (1984), sostienen que estos caracteres tienen mucha plasticidad, lo cual restringe su valor diagnóstico.
En las gramíneas, los caracteres de la epidermis que generalmente se toman en cuenta desde el punto de vista taxonómico, son la longitud y la forma de las células largas y cortas, la ondulación de la pared en las células largas, la amplitud máxima de las ondulaciones, la proporción y forma de las células cortas en la zona costal e intercostal, la posición de los aguijones, el tipo de estomas, la forma de las células subsidiarias y la presencia o ausencia de micropelos y macropelos (Dubé & Morisset 1996). Con base en estos caracteres algunos autores han intentado caracterizar los principales grupos de gramíneas. Por ejemplo, Twiss et al. (1969) reconocen cuatro clases de cuerpos de sílice, tres de los cuales corresponden a subfamilias de Poaceae: clase Panicoide, clase Festucoide y clase Chloridoide. Además, identifican la clase Elongata que no corresponde con una subfamilia en particular.
Por otra parte, Metcalfe (1960) mencionó que la distribución de los cuerpos de sílice es importante para reconocer algunos grupos taxonómicos, especialmente a nivel de subfamilia. Así, menciona que en gramíneas de tipo festucoide, los cuerpos de sílice son solitarios o pareados, mientras que en las de tipo panicoide se arreglan en hileras largas. Clifford & Watson (1977) encontraron que las células subsidiarias de los estomas con forma triangular se encuentran en aquellas gramíneas del tipo no festucoide y que las células en forma de domo o con los lados paralelos son características de las festucoide. Estos ejemplos hablan de las generalizaciones que se han realizado en las gramíneas y es un hecho que al menos, en ciertos grupos, el uso de caracteres epidérmicos ha resultado útil. El estudio de la anatomía de las hojas en las Poaceae se ha hecho tradicionalmente con microscopía de luz, pero se ha demostrado que el uso del microscopio electrónico de barrido puede ser complementario (Palmer 1976).
El género Zeugites pertenece a la tribu Centotheceae, la posición sistemática de la tribu ha sido controversial ya que se ha considerado como una tribu de las Bambusoideae (Watson & Dallwitz 1992), como una tribu con afinidades a las Arundinoideae (Renvoize 1986, Kellogg & Campbell 1987, Soderstrom & Ellis 1987, Barker et al. 1995, Mathews et al. 2000), y actualmente se considera una tribu de las Panicoideae (Clark et al. 1995, Hilu et al. 1999, Zuloaga et al. 2003, Sánchez- Ken & Clark 2007). Soriano et al. (2007), incluyen en el género Zeugites once especies: Z. smilacifolia Scribn. 1896, Z. capillaris (Hitchc.) Swallen 1943, Z. hackelii Swallen 1943, Z. latifolia (E. Fourn.) Hemsl. 1885, Z. sagittata Hartley 1941, Z. pittieri Hack. 1902, Z. munroana Hemsl. 1885, Z. americana Willd. 1805, Z. mexicana (Kunth) Trin. ex Steud. 1841, Z. pringlei Scribn. 1898 y Z. sylvatica (Soderstrom & Decker 1973). Estas son especies neotropicales, la mayoría tienen distribución restringida a México; algunas se extienden a Centroamérica (Z. munroana, Z. pittieri y Z. sylvatica), Sudamérica (Z. mexicana) y Z. americana se localiza en el Caribe, ocupan una gran variedad de hábitats que incluyen desde lugares sombreados, barrancas y laderas montañosas en el bosque de pino-encino, el bosque de Quercus, el bosque de Pinus, hasta la selva baja caducifolia y el bosque mesófilo de montaña. Se encuentran a una altitud entre 550 y 2 300m.s.n.m., sin embargo, las especies Z. hackelii, Z. latifolia, Z. sagittata y Z. sylvatica tienen distribución geográfica y ecológica restringida.
Considerando que no existen trabajos anatómicos completos del género Zeugites, en los cuales se revise en detalle la epidermis foliar, con excepción de Z. munrroana (McCoy 1934), Z. pringlei, Z. sylvatica y Z. pittieri (Renvoize 1986). En este trabajo se describe la epidermis foliar de los miembros del género Zeugites con base en observaciones realizadas con los microscopios electrónico de barrido y óptico.
Materiales y métodos
Para cada especie se revisaron al menos tres especímenes procedentes de colecciones de herbario y de material recolectado en el campo (Cuadro 1). Las preparaciones de la epidermis abaxial y adaxial de la lámina foliar se realizaron seleccionando porciones sanas de la parte media de la lámina en hojas maduras. Se utilizó la técnica de microscopía electrónica de barrido para la observación y caracterización de los taxones, que se complementaron mediante observaciones con microscopía de luz usando campo claro y contraste de fases.
Las muestras previamente fijadas en FAA (formol, ácido acético y alcohol), se deshidrataron con etanol al 50, 60, 70, 80, 90 y 100% por 1h. Posteriormente se pasaron a una cámara de desecación durante 2h a 42°C. Las muestras fueron adheridas a una base de aluminio usando una cinta de carbón conductivo y se cubrieron con una capa de oro de 40 mA. Se montaron dos porciones, una exponiendo la superficie abaxial y la otra la adaxial. Las observaciones se hicieron en un microscopio electrónico de barrido Hitachi S-2460N. Las fotografías se tomaron con una cámara Reflex Pentax Z-10 y un film Kodak 100 Tmax. El material que se analizó con el microscopio óptico se preparó utilizando la técnica sin tinción propuesta por Metcalfe (1960).
Las descripciones se realizaron siguiendo la terminología propuesta por Ellis (1979) y Palmer & Tucker (1981). En la descripción genérica se incluyeron todas las características de la epidermis observadas en las diferentes especies del género y en las descripciones de las especies se señalan los caracteres distintivos de cada taxón.
Resultados
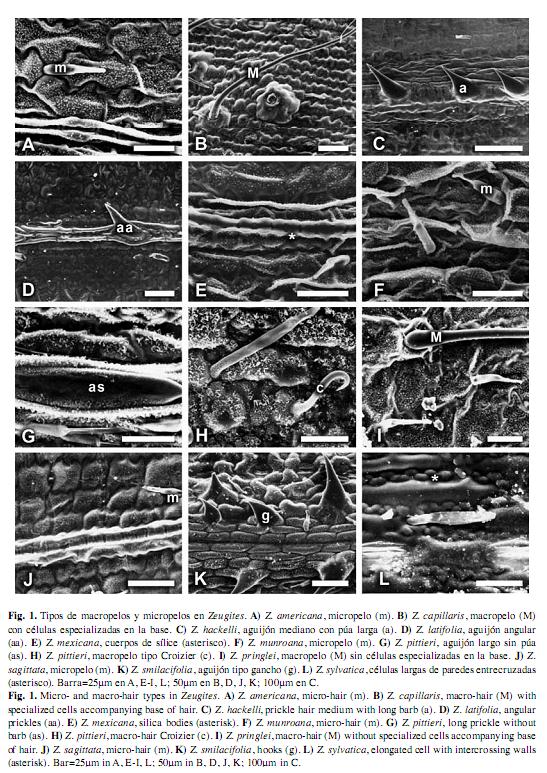
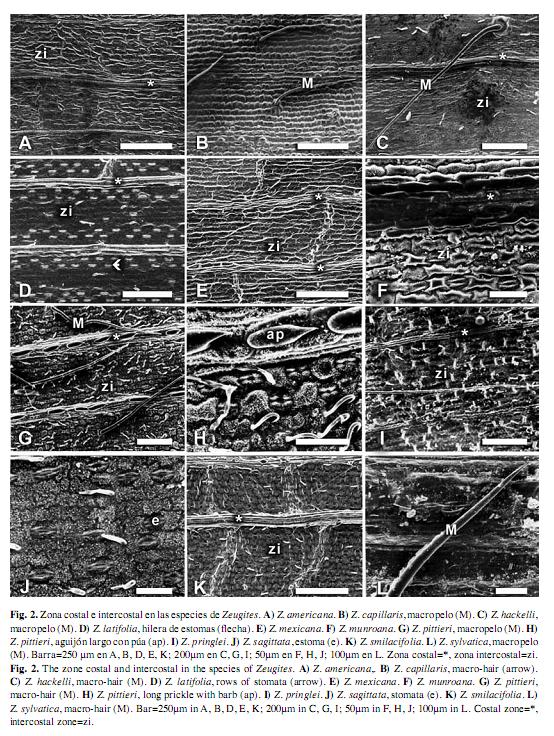
Zeugites: Células largas intercostales: largas y delgadas (usualmente tres veces más largas que anchas o más), rectangulares, cuadradas o ligeramente más altas que anchas; paredes irregulares, moderada a fuertemente onduladas o entrelazadas, moderada o fuertemente gruesas. Estomas: abundantes en la superficie adaxial, arreglados en dos a ocho hileras, escasos a frecuentes en la superficie abaxial; células subsidiarias triangulares con el domo alto u ovoides con el domo bajo. Células interestomáticas: rectangulares o cuadradas, 1-3(5) células o micropelos sucesivos entre estomas, bordes de las células ligeramente cóncavos (forma de U). Células cortas intercostales: ausentes, escasas o abundantes, solitarias o en pares: redondeadas a elípticas o alargadas verticalmente, con el contorno liso o irregular; cuando se encuentra en pares, una es de sílice con forma redondeada a elíptica y otra es de corcho en forma de media luna (crecentic) que rodea a la célula de sílice. Macropelos: a veces ausentes, cuando presentes de dos tipos, un tipo de 370-970μm de largo, frecuentes en la superficie adaxial y abaxial, formando una hilera en la zona media, generalmente suaves, con o sin células epidérmicas especializadas asociadas a la base del macropelo. El otro tipo de 29- 63μm de largo, rígidos con el ápice curvo (crozier), sin células epidérmicas especializadas asociadas a la base del macropelo. Células cortas costales: a veces ausentes, cuando presentes cuadradas o rectangulares, con paredes sinuosas. Cuerpos de sílice: agrupados en hileras, en forma de cruz o halteriformes, con la porción central ancha y los bordes redondeados o con la porción central constreñida y los bordes planos o redondeados. Aguijones: a veces ausentes, cuando presentes escasos o abundantes en la superficie adaxial, abaxial o en ambas, largos (base al menos dos veces más larga que un estoma), medianos (base tan larga como alta, casi del mismo tamaño que un estoma), púa a veces ausente, cuando presente corta (más corta que la base) o larga (más larga que la base). Micropelos: bicelulares, comunes en la zona intercostal de la superficie abaxial y adaxial (infrecuentes en la zona costal), célula basal más corta o igual en tamaño que la célula apical, ápice ahusado.
Zeugites americana (Fig. 1.A; Fig. 2.A): Células largas intercostales: largas y delgadas, rectangulares; paredes moderada o fuertemente onduladas. Estomas: abundantes en ambas superficies, arreglados en 2-5 hileras, células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células y micropelos sucesivos entre estomas. Células cortas intercostales: escasas, solitarias, redondeadas a elípticas, con el contorno liso y en pares, una de sílice y otra de corcho, la de corcho con forma de media luna rodeando a la célula de sílice, ésta última redondeada a elíptica. Macropelos: ausentes. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central ancha, bordes redondeados. Aguijones: ausentes. Micropelos: la célula basal casi tan larga como la apical.
Zeugites capillaris (Fig. 1.B; Fig. 2.B): Células largas intercostales: rectangulares, cuadradas, algunas ligeramente más altas que anchas; paredes fuertemente onduladas. Estomas: abundantes en la superficie adaxial, arreglados en 3-5 hileras, escasos a frecuentes en la superficie abaxial; células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células sucesivas entre estomas. Células cortas intercostales: ausentes. Macropelos: 370-571μm de largo, frecuentes en ambas superficies, una a dos hileras en la zona media, unicelulares, suaves, rectos, con células epidérmicas especializadas asociadas a la base del macropelo. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central constreñida, bordes planos. Aguijones: en la zona costal, infrecuentes, en ambas superficies, medianos con púa corta o largos con púa corta o sin púa; en la zona intercostal, ausentes. Micropelos: la célula basal más corta que la apical.
Zeugites hackelii (Fig. 1.C; Fig. 2.C): Células largas intercostales: ligeramente más altas que anchas, paredes fuertemente onduladas. Estomas: abundantes en ambas superficies, arreglados en 2-4 hileras, células subsidiarias triangulares con el domo alto. Células interestomáticas: una a tres células sucesivas entre estomas. Células cortas intercostales: escasas, solitarias, alargadas verticalmente con el contorno liso. Macropelos: 520-968μm de largo, frecuentes en ambas superficies, arreglados en una hilera en la zona media,unicelulares, suaves, rectos, con células epidérmicas especializadas asociadas a la base del macropelo. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central constreñida, bordes planos. Aguijones: en la zona costal, abundantes, en ambas superficies, medianos con púa corta o largos con púa corta o larga; en la zona intercostal, ausentes. Micropelos: la célula basal casi tan larga como la apical.
Zeugites latifolia (Fig. 1.D; Fig. 2.D): Células largas intercostales: rectangulares a cuadradas, paredes fuertemente onduladas. Estomas: abundantes en ambas superficies, arreglados en 5-7 hileras, células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células sucesivas entre estomas. Células cortas intercostales: ausentes. Macropelos: ausentes. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central constreñida, bordes redondeados. Aguijones: en la zona costal, abundantes en la superficie abaxial, medianos, con púa corta o larga y angulares con púa tan larga como la base; en la zona intercostal ausentes. Micropelos: la célula basal más corta que la apical.
Zeugites mexicana (Fig. 1.E; Fig. 2.E): Células largas intercostales: largas y delgadas, rectangulares; paredes moderada o fuertemente onduladas. Estomas: abundantes en ambas superficies, arregladas en 2-7 hileras, células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células y micropelos sucesivos entre estomas. Células cortas intercostales: escasas, solitarias, alargadas verticalmente con el contorno liso. Macropelos: ausentes. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central ancha, bordes redondeados. Aguijones: ausentes. Micropelos: la célula basal casi tan larga como la apical.
Zeugites munroana (Fig. 1.F; Fig. 2.F): Células largas intercostales: largas y delgadas, rectangulares, paredes moderada o fuertemente onduladas. Estomas: abundantes en ambas superficies, arreglados en 2-4 hileras, células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células y micropelos sucesivos entre estomas. Células cortas intercostales: escasas, solitarias, de redondeadas a elípticas, con el contorno liso. Macropelos: ausentes. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central constreñida, bordes redondeados. Aguijones: ausentes. Micropelos: la célula basal casi tan larga como la apical.
Zeugites pittieri (Fig. 1.G,H; Fig. 2.G,H): Células largas intercostales: largas y delgadas, rectangulares; paredes moderada o fuertemente onduladas. Estomas: abundantes en la superfície adaxial, arreglados en 2-3 hileras, escasos a frecuentes en la superficie abaxial; células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células sucesivas entre estomas. Células cortas intercostales: escasas, solitarias, redondeadas a elípticas o alargadas verticalmente, con el contorno liso o irregular. Macropelos: unicelulares, frecuentes en ambas superficies, arreglados en una hilera en la zona media, de dos tipos: un tipo, 542-560μm de largo, suaves, rectos, con cιlulas epidérmicas especializadas asociadas a la base del macropelo, el otro tipo (crozier), 29-63μm de largo, rígidos, ápice curvo, sin células epidérmicas especializadas asociadas a la base del macropelo. Células cortas costales: cuadradas o rectangulares, con paredes sinuosas. Cuerpos de sílice: halteriformes, porción central constreñida, bordes redondeados. Aguijones: en la zona costal, infrecuentes, en ambas superficies, largos con púa corta o sin púa; en la zona costal, ausentes. Micropelos: la célula basal más corta que la apical.
Zeugites pringlei (Fig. 1.I; Fig. 2.I): Células largas intercostales: largas y delgadas, rectangulares; paredes moderada o fuertemente onduladas. Estomas: abundantes en ambas superficies, arreglados en 2-4 hileras, células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células y micropelos sucesivos entre estomas. Células cortas intercostales: ausentes. Macropelos: 428-552μm de largo, frecuentes en la superficie adaxial, arreglados en una hilera en la zona media, unicelulares, suaves, sin células epidérmicas especializadas asociadas a la base del macropelo. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central ancha, bordes redondeados. Aguijones: ausentes. Micropelos: la célula basal casi tan larga como la apical.
Zeugites sagittata (Fig. 1.J; Fig. 2.J): Células largas intercostales: rectangulares a cuadradas, paredes fuertemente onduladas. Estomas: abundantes en la superficie adaxial, arreglados en 2-6 hileras, escasos en la superficie abaxial; células subsidiarias triangulares con el domo alto. Células interestomáticas: una a cinco células sucesivas entre estomas. Células cortas intercostales: escasas, solitarias, redondeadas a elípticas, con el contorno liso. Macropelos: ausentes. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, porción central constreñida, bordes planos. Aguijones: ausentes. Micropelos: la célula basal casi tan larga como la apical.
Zeugites smilacifolia (Fig. 1.K; Fig. 2.K): Células largas intercostales: rectangulares, cuadradas o ligeramente más altas que anchas; paredes moderada o fuertemente onduladas. Estomas: abundantes en ambas superficies, arreglados en 5-8 hileras, células subsidiarias triangulares con el domo alto. Células interestomáticas: una a dos células sucesivas entre estomas. Células cortas intercostales: escasas, solitarias, redondeadas a elípticas, con el contorno liso. Macropelos: ausentes. Células cortas costales: ausentes. Cuerpos de sílice: halteriformes, con la porción central constreñida, bordes planos. Aguijones: en la zona costal, en ambas superficies, angulares, medianos con púa igual o más larga que la base; en la zona intercostal, ganchos con la base redondeada, medianos, con la púa tan larga como la base. Micropelos: la célula basal casi tan larga como la apical.
Zeugites sylvatica (Fig. 1.L; Fig. 2.L): Células largas intercostales: largas y delgadas, rectangulares, paredes entrelazadas. Estomas: abundantes en la superficie adaxial, arreglados en 1-2 hileras, escasos a frecuentes en la superficie abaxial; células subsidiarias ovoides con el domo bajo. Células interestomáticas: una a tres células sucesivas entre estomas. Células cortas intercostales: abundantes, redondeadas a elípticas, con el contorno liso, o alargadas verticalmente con el contorno liso o irregular. Macropelos: 670-800μm de largo, frecuentes, en la superficie abaxial, arreglados en una hilera en la zona media, unicelulares, suaves, rectos, con células epidérmicas especializadas asociadas a la base del macropelo. Células cortas costales: cuadradas o rectangulares, con paredes sinuosas. Cuerpos de sílice: en forma de cruz con cuatro ápices no redondeados, perpendiculares. Aguijones: en la zona costal, infrecuentes en la superficie abaxial, largos con púa corta o sin púa; en la zona costal, ausentes. Micropelos: la célula basal más corta que la célula apical.
En el género Zeugites existe una gran variación en la morfología de la epidermis foliar, en especial, en la presencia y ausencia de aguijones y macropelos, en las formas de las células cortas intercostales y en los cuerpos de sílice. Estas características han resultado útiles para caracterizar las especies (ver clave para el género).
Con base en las características de los aguijones y los macropelos, las especies de Zeugites se pueden separar en tres grupos. En el primer grupo están las especies Z. americana, Z. mexicana, Z. pringlei, Z. munroana y Z. sagittata, las cuales no presentan aguijones. Z. pringlei presenta macropelos largos y suaves en toda la lámina, carácter que la distingue de Z. americana y Z. mexicana que llegan a presentar macropelos en una hilera cerca de la nervadura central; éstas tres especies, fueron consideradas variedades de Z. americana (MacVaugh 1983). Las especies en este grupo se distinguen además, por la forma de los cuerpos de sílice, en Z. sagittata y Z. munroana, éstos son halteriformes con la porción central constreñida, mientras que en Z. americana, Z. mexicana y Z. pringlei, los cuerpos de sílice también son halteriformes, pero con la porción central ancha. Por su parte, Z. americana es la única especie del género que además de presentar células cortas intercostales solitarias, también presenta pares de células, una de sílice y otra suberosa. La principal diferencia entre Z. mexicana y Z. munroana radica en la forma de las células cortas intercostales. En la primera, estas células están alargadas verticalmente, mientras que en Z. munroana son redondeadas a elípticas.
En el segundo grupo están las especies Z. latifolia y Z. smilacifolia que sólo tienen aguijones y no presentan macropelos. En ambas especies, los aguijones están tanto en la superficie abaxial como en la adaxial, pero en Z. latifolia se encuentran únicamente en la zona costal, mientras que en Z. smilacifolia se encuentran tanto en la zona costal como en la intercostal. Además, las células cortas intercostales están ausentes en Z. latifolia y presentes en Z. smilacifolia.
En el tercer grupo, se encuentran las especies que se caracterizan por tener aguijones y macropelos (Z. hackelii, Z. capillaris, Z pittieri y Z. sylvatica). En estas especies los macropelos tienen en la base un conjunto de células especializadas (células diferentes a las epidérmicas), con excepción de Z. pittieri en la cual dichas células están ausentes. Los macropelos son suaves y rectos de 360-968μm de largo. En Z. pittieri además, se encuentra un segundo tipo de macropelos curvos, de 29-63μm de largo, es la ϊnica especie del género que presenta esta característica. Macropelos similares se han encontrado en Orthoclada laxa ((Rich.) P. Beauv. 1812), otro miembro de la tribu Centotheceae (Lersten & Phol 1969, Renvoize 1986).
Zeugites sylvatica fue reconocida como Calderonella sylvatica (Soderstrom 1981, Soderstrom & Decker 1973), sin embargo, los resultados del análisis filogenético del grupo utilizando datos morfológicos y secuencias nucleares (ITS) y del cloroplasto (trnL) (Soriano et al. 2007), apoyaron la inclusión de C. sylvatica dentro de los límites taxonómicos del género Zeugites. Z. sylvatica se distingue de las otras especies por tener las paredes engrosadas y entrelazadas, los cuerpos de sílice tienen forma de cruz, con los cuatro ápices no redondeados y las células subsidiarias ovoides con el domo bajo. Algunos de estos atributos los comparte con otros miembros de la familia Panicoideae. Por ejemplo, la forma de las paredes es similar a Orthoclada laxa y los cuerpos de sílice son semejantes a Thysanolaena maxima (Roxb.) Kuntze 1891, de la tribu Centotheceae. Por su parte, las células subsidiarias son parecidas a las que presentan Digitaria sanguinalis (L.) Scop. 1771, de la subtribu Digitariinae y Echinochloa crus-galli (L.) P. Beauv. 1812 y Panicum virgatum L. 1753, de la subtribu Panicinae (A. Soriano, en prep.).
Las especies Z. hackelii y Z. capillaris se distinguen principalmente por la abundancia y forma de los aguijones y los macropelos. Z. hackelii presenta aguijones abundantes, únicamente en la zona costal. En el caso de Z. capillaris los aguijones se presentan tanto en la zona costal como en la intercostal. En Z. hackelii los aguijones son más abundantes, mientras que en Z. capillaris son los macropelos los elementos dominantes. Además, en Z. hackelii las células cortas intercostales están presentes mientras que en Z. capillaris dichas células están ausentes.
La presencia de tricomas se observa en hojas de plantas que viven en hábitat xéricos como un mecanismo para reducir la transpiración (Esau 1977, Fahn 1986, Andersen et al. 2006). En este sentido las especies de Zeugites presentan este patrón, ya que las que no tienen aguijones ni macropelos (Z. americana, Z. mexicana y Z. munrroana) habitan a orillas de ríos, lagos, o en bosques de niebla. En contraste, las especies que presentan aguijones o macropelos fueron recolectadas en ambientes que aún y cuando pueden ser muy húmedos en una buena parte del día quedan expuestos al sol. La excepción a este patrón es Z. sagittata que no presenta ni aguijones ni macropelos y puede encontrarse en lugares muy perturbados.
Las especies de Zeugites comparten una serie de características tales como la forma de las células largas, estomas, células subsidiarias, células cortas, macropelos, cuerpos de sílice y micropelos, Sin embargo, algunos de sus miembros presentan características similares con otros taxones, que podrían indicar relaciones, como son los macropelos curvos de Z. pittieri que son semejantes a los que presenta Orthoclada laxa de la tribu Centotheceae. Otros caracteres que relacionan directamente al género con la subfamilia Panocoideae son el tipo de micropelos y los cuerpos de sílice en hilera (Metcalfe 1960). Además, de las características que presenta Z. sylvatica que comparte con miembros de la tribu Paniceae, al parecer es el miembro de Zeugites que más relaciona al género con miembros de la subfamilia Panicoideae.

Agradecimientos
Se agradece a los curadores de los herbarios que facilitaron el material analizado, así como a Berenit Mendoza y a Julio César Montero del Instituto de Biología, UNAM, por la asistencia técnica en el microscopio electrónico de barrido y por el trabajo gráfico respectivamente.
Referencias
Aiken, S.G. & L.P. Lefkovitch. 1984. The taxonomic value of using epidermal characteristics on the Canadian rough fescues complex (Festuca altica, Festuca campestris, Festuca halii, "Festuca scabrella"). Can. J. Bot. 62: 1864-1870. [ Links ]
Andersen, A., F.F. Lucchini, J. Moriconi & E.A. Fernández. 2006. Variabilidad en la morfo-anatomía foliar de Lippia turbinata (Verbenaceae) en la provincia de San Luis (Argentina). Phyton 75: 137-143. [ Links ]
Barker, N.P., H.P. Linder & E.H. Harley. 1995. Polyphyly of Arundinoideae (Poaceae): Evidence from rbcL sequence data. Syst. Bot. 20: 423-435. [ Links ]
Barkworth, M.E. 1981. Foliar epidermis and taxonomy of North American species of Stipa (Gramineae). Syst. Bot. 6: 136-152. [ Links ]
Clark, L.G., W. Zhang & J.F. Wendel. 1995. A phylogeny of the grass family (Poaceae) based on ndhF sequence data. Syst. Bot. 20: 436-460. [ Links ]
Clifford, H.T. & L. Watson. 1977. Identifying grasses: data, methods and illustrations. St. Lucia, Queensland, Australia. [ Links ]
Dávila, P. & L.G. Clark. 1990. Scanning electron microscopy of leaf epidermis of Sorghastrum (Poaceae: Andropogoneae). Amer. J. Bot. 77: 499-511. [ Links ]
Dubé, M. & P. Morisset. 1996. L’emploi des caractères épidermiques dans l’étude taxonomique du Festuca rubra sensu lato (Poaceae). Can. J. Bot. 74: 469-485. [ Links ]
Ellis, R.P. 1979. A procedure for standardizing comparative leaf anatomy in the Poaceae, II. The epidermis as seen in surface view. Bothalia 12: 641-671. [ Links ]
Esau, K. 1977. Anatomy of seed plants. John Wiley, Nueva York, EEUU. [ Links ]
Fahn, A. 1986. Structural and functional properties of trichomes of xeromorphic leaves. Ann. Bot. 57: 631-637. [ Links ]
Hilu, K.W. 1984. Leaf epidermis of Andropogon sect. Leptopogon (Poaceae) in North America. Syst. Bot. 9: 247-257. [ Links ]
Hilu, K.W., L.A. Alice & H. Liang. 1999. Phylogeny of Poaceae inferred from matK sequences. Ann. Missouri Bot. Gard. 86: 835-851. [ Links ]
Holmgren, P.K., W. Keuken & E.K. Schofield. 2004. Index herbariorum. International Association for Plant Taxonomy. The New York Botanical Garden, Nueva York, Nueva York, EEUU. [ Links ]
Jacques-Felix, H. 1962. Les Graminées (Poaceae) d´ Afrique tropicale. Institut of Recherches Agronomiques Tropicales et des Culture Vivieres, París, Francia. [ Links ]
Kellogg, E.A. & C.S. Campbell. 1987. Phylogenetic analyses of the Gramineae, p. 310-322. In T.R. Soderstrom, K.W. Hilu, C.S. Cambpell & M.E. Barkworth (eds.). Grass systematics and evolution. Smithsonian Institution, Washington, EEUU. [ Links ]
Lersten, N.R. & R.W. Pohl. 1969. Anatomical and chromosomal observations on Orthoclada laxa (Centotheceaea; Gramineae). Amer. J. Bot. 56: 1054-1057. [ Links ]
Mathews, S., R.C. Tsai & E.A. Kellogg. 2000. Phylogenetic structure in the grass family (Poaceae): Evidence from the nuclear gene phytochrome B. Amer. J. Bot. 87: 96-107. [ Links ]
McCoy, R.W. 1934. The anatomy of the leaf of Zeugites munroana, an anomalous grass. Bull. Torrey Bot. Club 61: 429-436. [ Links ]
McVaugh, R. 1983. Flora Novo Galiciana. 14. Gramineae. The University of Michigan, Michigan, EEUU. [ Links ]
Metcalfe, C.R. 1960. Anatomy of the monocotyledons. I. Gramineae. Oxford University, Oxford, Reino Unido. [ Links ]
Palmer, P.G. 1976. Grass cuticle: A new paleoecological tool for East African Lake sediments. Can. J. Bot. 54: 1725-1734. [ Links ]
Palmer, P.G. & A.E. Tucker. 1981. A scanning electron microscope survey of East African grasses. I. Smithsonian Contrib. Bot. 49: 1-84. [ Links ]
Prat, H. 1932. L’épiderme des Graminées. Étude anatomique et systématique. Ann. Sci. Nat. Bot. 14: 117-324. [ Links ]
Prat, H. 1936. La Systématique des Graminées. Ann. Sci. Nat. Bot. 18: 165-258. [ Links ]
Renvoize, S.A. 1986. A survey of leaf-blade anatomy in grasses. IX. Centothecoideae. Kew Bull. 41: 339-342. [ Links ]
Sánchez-Ken, J.G. & L.G. Clark. 2007. Phylogenetic relationships within the clade Centothecoideae+Panicoideae (Poaceae), based on ndhF and rpl16 sequences and morphological data, p. 467-482. In J.T. Columbus, E.A. Friar, J.M. Porter, L.M. Prince & G. Simpsom (eds.). Monocots: comparative biology and evolution - Poales. Aliso, Rancho Santa Ana Botanic Garden, California, EEUU. [ Links ]
Soderstrom, T.R. 1981. The grass subfamily Centothecoideae. Taxon 30: 614-616. [ Links ]
Soderstrom, T.R. & H.F. Decker. 1973. Calderonella, a new genus of grasses, and its relations to the centostecoid genera. Ann. Missouri Bot. Gard. 60: 427-441. [ Links ]
Soderstrom, T.R. & R.P. Ellis. 1987. The position of bamboo genera and allies in a system of grass classification, p. 225-238. In T.R. Soderstrom, K.W. Hilu, C.S. Campbell & M.E. Barkmorth (eds.). Grass systematics and evolution. Smithsonian Institution, Washington, EEUU. [ Links ]
Soriano, M.A.M., G.A. Salazar & P. Dávila A. 2007. Phylogenetic relationships of Zeugites (Poaceae: Centothecoideae) inferred from plastid and nuclear DNA sequences and morphology. Syst. Bot. 32: 722-730. [ Links ]
Tateoka, T., S.S. Inoue & S. Kawano. 1959. Notes on some grasses. IX. Systematic significance of bicellar microhairs of leaf epidermis. Bot. Gaz. 124: 264-270. [ Links ]
Twiss, P.C., E. Suess & R.M. Smith. 1969. Morphological classification of grass phytoliths. Soil Sci. Soc. Am. J. 33: 109-115. [ Links ]
Watson, L. & M.J. Dallwitz. 1992. The grass genera of the world. CAB International, Wallingford, Reino Unido. [ Links ]
Zuloaga, F.O., O. Morrone, G. Davidse, T.S. Filgueiras, P.M. Peterson, R.J. Soreng & E.J. Judziewicz. 2003. Catalogue of new world grasses (Poaceae): III. Subfamilies Panicoideae. Aristidoideae, Arundinoideae, and Danthonioideae. Contr. U.S. Natl. Herb. 46:1-662. [ Links ]
Correspondencia a: Ana María Soriano. Red de Biodiversidad y Sistemática, Instituto de Ecología, A.C. Carretera antigua a Coatepec 351, El Haya, Xalapa 91070, Veracruz, México; ana.soriano@inecol.edu.mx
Teresa Terrazas.Departamento de Botánica, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado Postal 70-367, 04510, Distrito Federal, México; tterrazas@ibiologia.unam.mx
Patricia Dávila. Unidad de Biología Tecnología y Prototipos (UBIPRO), Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México. Avenida de los Barrios No. 1, Los Reyes Iztacala, 54090 Tlalnepantla, Estado de México, México; pdavilaa@servidor.unam.mx
Recibido 15-VI-2010. Corregido 16-XII-2010. Aceptado 25-I-2011.


{kind=link}
{kind=link}
{kind=link}