










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.2 San José Jun. 2011
Macrophytes in the upper Paraná river floodplain: checklist and comparison with other large South American wetlands
Fernando Alves Ferreira1, Roger Paulo Mormul1, Sidinei Magela Thomaz2, Arnildo Pott3 & Vali Joana Pott3
1. Postgraduate Program in Ecology of Continental Aquatic Environments, Universidade Estadual de Maringá- UEM, Av. Colombo 5790, bloco H-90, CEP 87020-900, Maringá, Paraná, Brazil; ferreirabot@gmail.com, roger.mormul@gmail.com2. Research Group in Limnology, Ichthyology and Aquaculture-Nupelia, Biological Science Department, Universidade Estadual de Maringá-UEM, Av. Colombo 5790, bloco H-90, CEP 87020-900, Maringá, Paraná, Brazil; smthomaz@nupelia.uem.br
3. Biological Science Department, Universidade Federal do Mato Grosso do Sul - UFMS CEP 79.070-900, Caixa - Postal 549, Campo Grande, MS, Brazil; arnildo.pott@gmail.com, vali.pott@gmail.com
Dirección para correspondencia
Abstract
Key words: floristic survey, plant diversity, aquatic plants, Brazil.
Resumen
Los ecosistemas acuáticos neotropicales tienen una rica flora acuática. En este informe, hemos hecho una lista de la flora acuática de diversos hábitats de la alta planicie de inundación del río Paraná mediante la compilación de datos de la literatura y los registros de nuestras colecciones propias realizadas durante el período 2007-2009. Nuestros principales objetivos fueron evaluar la riqueza de macrófitos en la llanura aluvial del Paraná, para compararlo con otros humedales de América del Sur y evaluar si el número de especies registradas en los inventarios suramericanos ya han alcanzado una asíntota. Se registraron un total de 153 especies de macrófitas en la llanura de inundación del Río Alto Paraná, pertenecientes a 100 géneros y 47 familias. En nuestro análisis comparativo, se mostró una clara división florística de otros humedales de América del Sur, con excepción del Pantanal, que es el más cercano a los humedales de la planicie de inundación del Paraná y, por tanto, podría considerarse una extensión florística del Pantanal. La curva de acumulación de especies demuestra que los esfuerzos de muestreo deben ser reforzados con el fin de elaborar un censo de la flora de macrófitos para América del Sur. La alta disimilitud entre los humedales de América del Sur, junto con la falta de una asíntota en nuestra curva de acumulación de especies, indica que el esfuerzo de muestreo debe ser mayor para dar cuenta de la riqueza real de las especies de macrófitos en esta región.
Palabras clave: Inventario florístico, diversidad de plantas, plantas acuáticas, Brasil.
One of the main ecological characteristics of South America is the existence of large wetlands (Neiff 2001). Inventories of wetlands provide an indication of the sites with the highest biological diversity and productivity (Taylor et al. 1995), and the information collected through inventories is a necessary prerequisite for conservation policies (Pressey & Adam 1995).
Approximately 50% of the inventoried wetlands in South America are located in Brazil (Naranjo 1995). However, specific information related to aquatic macrophytes is extremely scarce. Diegues (1994) performed the first inventory of wetlands in Brazil, and his work provided valuable data for evaluating ecological and economic aspects of these regions. According to Neiff (1978, 1986), aquatic macrophytes are important in shallow ecosystems, such as river-floodplain ecosystems, where they colonize extensive areas and exhibit high rates of primary productivity. In addition, macrophytes are a key component of riverfloodplain ecosystems because they enhance nutrient cycling, increase habitat heterogeneity and provide food for a variety of organisms (Esteves 1998).
Floodplains are known as ecosystems with a high diversity of habitats and aquatic and terrestrial species (Junk et al. 2000). Due to their high complexity and seasonal changes in physico-chemistry, these ecosystems are characterized by a variety of assemblages, which differ in richness and composition according to the water level. In the Upper Paraná River floodplain, for example, the vegetation is highly conditioned by geomorphology (Souza-Filho 1993); trees dominate the more elevated areas (levees), and shrubs colonize less elevated areas that remain flood- free most of the year, while aquatic macrophytes grow in permanently inundated areas of the wetlands.
Despite the importance of these macrophytes in the Upper Paraná River floodplain, a stretch of this river that is key in maintaining the biodiversity of Brazilian inland waters, information about the aquatic vegetation in this region is scattered among several different papers and reports (Bini 1996, Kita & Souza 2003, Thomaz et al. 2004, Thomaz et al. 2009); most of these studies emphasized that the flood pulse and changes in water physico-chemistry are important factors controlling macrophyte populations and communities.
In the present study, we first addressed the number of macrophyte species in the main habitats of the Upper Paraná River and its floodplain (herein only Paraná floodplain), using records gathered since 1997 and intensive collections performed between 2007 and 2009. Secondly, we used this dataset to compare the species richness and similarity of this area with other South American wetlands. Finally, using species accumulation curves, we examined whether the number of species described in South America is reaching an asymptote, or if more sampling efforts are still necessary to accomplish a comprehensive inventory of the rich aquatic flora of this area.
To accomplish these objectives, we adopted the conceptualization of aquatic macrophytes proposed by Cook (1996), in which the author includes plants which photosynthetically active organs are either permanently, or for several months of the year, total or partially submersed in freshwater or floating in aquatic habitats. More recently, Chambers et al. (2008) also included Charophytes within the definition of macrophytes. To avoid any confusion, we did not use in any part of our text the term "vascular plants" but, instead, consistently used the term "aquatic macrophytes".
Materials and methods
Study area: The floodplain of the Paraná River is located downstream from the Porto Primavera Reservoir. This stretch has a length of 160km and is the last region of the river that remains not dammed in Brazilian territory. Thus, it is of key importance to the conservation of the aquatic biodiversity of the Paraná Basin (Agostinho & Zalewiski 1995).
According to the Köppen system, the climate in this region is classified as tropical and sub-tropical, with warm summers (mean annual temperature 22oC) and a mean annual rainfall of 1 500mm (Maack 2002).
The compiled list of taxa was based on records of samplings conducted in the floodplain since 1997. In addition, we utilized several other studies (published and unpublished) that had been conducted in the floodplain. Macrophytes were collected in a variety of habitats, such as the river main channel, lateral channels (anabranches), temporary and permanent lakes, and in the aquatic-terrestrial transition zone (ATTZ, sensu Junk et al. 1989). We also analyzed and revised specimens deposited in the Laboratory of Macrophytes and in the Herbarium (HUEM) of the University of Maringá. To complement the list of species recorded in previous investigations, we carried out additional samplings between 2007 and 2009 in six habitats that are being monitored in the Brazilian Long Term Ecological Research Program (site 6; Thomaz et al. 2009).
In each lake, the aquatic macrophytes were analyzed by boat at a slow speed along the entire shoreline. In the ATTZ, samplings were carried out on foot. We used a grapple attached to a line to record submersed species. Because ponds and lakes have small areas (from 0.006 to 113.8ha) and samplings were carried out on the entire shores, we considered the recorded species as the actual richness of these habitats, and did not correct the results to account for sampling effort (rarefaction curves, for example).
Identification followed comparative morphology and a specialized bibliography (e.g., Hoehne 1948, Cook 1996, Pott & Pott 2000, Amaral et al. 2008, Bove & Paz 2009). The list of taxa contains families and genera according to the "Angiosperm Phylogeny Group-APG II" (2003) for Magnoliophyta (Angiospermae), Willis (1973) for Pteridophyta, and Crandall-Stotler (1980) for Hepatophyta.
Plant life forms were chosen according to Pedralli (1990), and we followed Tur (1972) for epiphytic forms. Plants growing in wet soils (marshes locally known as "varjão" or "várzea") were included in the category of amphibious (Irgang & Gastal 1996).
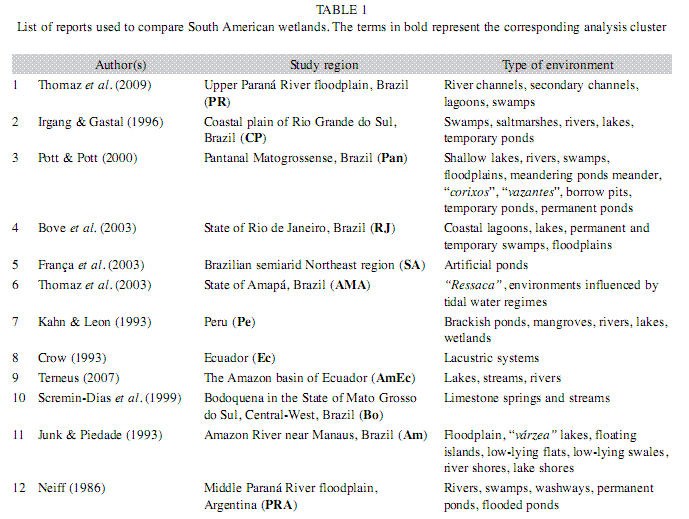
In order to make comparisons among South American wetlands, we used the following lists of macrophytes obtained in long-term surveys (Table 1).
All of the investigations and the lists of species that we used were carried out by specialists and included several types of habitats (Fig. 1). Although there are several other papers describing single habitats, we did not use these studies. It is difficult to guarantee that all studies follow the same methodology, but we believe that they are similar enough to at least contribute a first tentative of comparison of Neotropical wetlands to make inferences about the richness of macrophytes in this region.
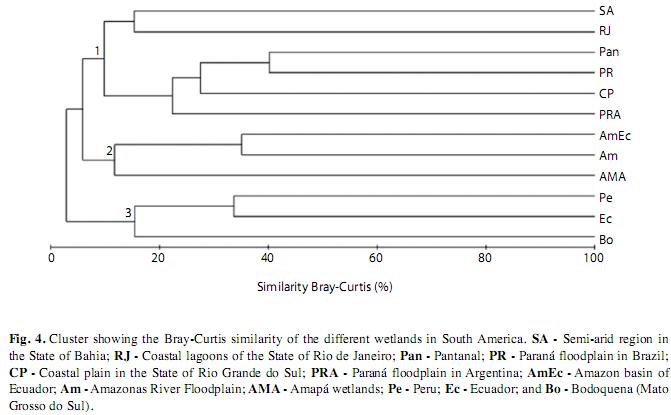
To find similarity among these surveys, we first converted all data into a large matrix containing species occurrence presence/absence. A matrix of similarity was built using the Bray-Curtis distance coefficient (Krebs 1999). To compare all surveys, we used the method of complete linkage (Sneath & Sokal 1973). A dendrogram of similarities was built using the PRIMER v. 6 software, Plymouth Routines in Multivariate Ecological Research (Clarke & Gorley 2006).
Using the entire dataset, which included all wetlands, we assessed whether the richness of macrophytes in South America reaches an asymptote, or if there are many species yet to be found. The expected species accumulation curve was calculated according to a Mao Tau function. Using an accumulative curve with "studies" as units of sampling effort, an asymptote would indicate whether almost all species have already been recorded; however, the lack of an asymptote would indicate that the number of aquatic macrophyte species found until now is still far from the real total of these species. We also estimated the richness of macrophytes using a first-order Jacknife estimator (Jack1) with the objective to assess the extent to which the number of macrophyte species in South America remains underestimated. Accumulation and estimation curves were constructed using the EstimateS program (Colwell 2009).
Results
A total of 153 species of macrophytes was recorded in the Upper Paraná River floodplain. These species were distributed in 100 genera and 37 families (Appendix 1), representing a variety of taxonomic groups (Charophyta Bryophyta, Pteridophyta, Basal Angiospermae and Angiospermae).
Sixteen of the recorded species are cryptogams and are classified as follows: two charophytes, two hepatophytes and 12 pteridophytes. Of the Angiospermae, Poales exhibited the highest number of taxa (40), followed by Alismatales (17), Myrtales and Lamiales (12 species each). The families with the highest numbers of species were Poaceae (21), Cyperaceae (17), Pontederiaceae (8), Hydrocharitaceae (7), Polygonaceae and Onagraceae (6) and Fabaceae (5). Araceae, Alismataceae, Commelinaceae, Amaranthaceae and Plantaginaceae were represented by four species each and the other families by three or fewer species.
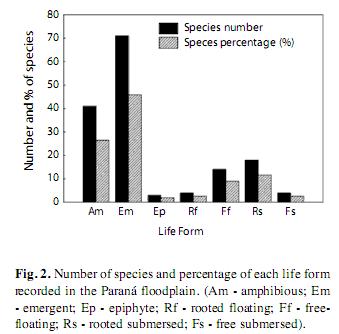
All life forms were found in the area, and emergent and amphibious types were the most representative macrophytes, contributing 45% and 26% of the species, respectively (Fig. 2). Rooted submersed (11%) and free-floating species (9%) were also important, while the lowest numbers of species were found for rooted floating and free-submersed types (2%) and epiphytes (1%).
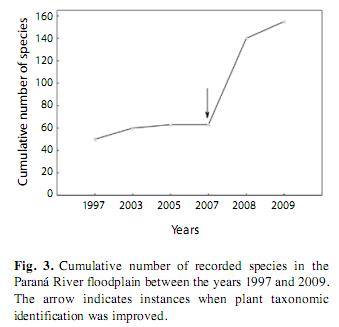
The number of species recorded in the Paraná River and its floodplain consistently increased over time (Fig. 3). The greatest increase occurred between 2007 and 2009, due to intensified sampling efforts and refined taxonomic identification, which led to the addition of 105 species to the recorded flora of the Upper Paraná River floodplain.
The Paraná floodplain exhibits the third highest richness of macrophytes (153 species) of the 12 areas for which we have data in South America. The coastal area of the State of Rio Grande do Sul (Brazil) ranked first (321 species), and the Pantanal Matogrossense, one of the largest wetlands in the world, ranked second (247 species).
A dendrogram built using the Bray-Curtis coefficient of distance showed that South American wetlands are dissimilar with respect to macrophyte assemblages (Fig. 4). We can roughly recognize three groups of wetlands. The first includes the seasonal ponds (Northeast Brazil), coastal lagoons of the State of Rio de Janeiro, Pantanal, Paraná floodplain in Brazil, the coastal plain in the State of Rio Grande do Sul and the Paraná floodplain in Argentina, with a similarity of 11.7% (Fig. 4). These areas share only two species in common, Polygonum ferrugineum and Nymphoides indica, both of which are widespread hydrophytes. In this first group, the most similar areas were the Paraná floodplain and the Pantanal Matogrossense with 40.2% similarity and 79 species in common (9% of the total species)The second group included the Amazon basin of Ecuador, Amazon River floodplain and Amapá wetlands, with a similarity of 13% and sharing four species, Eichhornia azurea, Hymenachne amplexicaulis, Salvinia auriculata and Utricularia foliosa. Within this group, the aquatic flora of the Amazon River floodplain and the Amazon Basin of Ecuador share 13 species (2% of the total species) and exhibit 35.1% similarity. Finally, group three was formed by Peru, Ecuador and Bodoquena (Mato Grosso do Sul), with a similarity of 18% and sharing 11 species; within this group, Peru and Ecuador had the highest similarity of 33.6% and sharing 35 species (4% of all species).
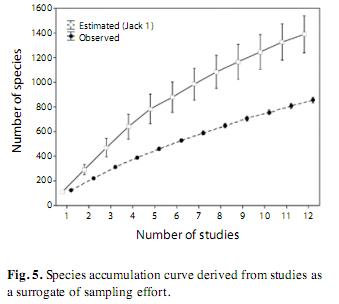
Considering all of the surveys that we found for South America together with the survey we conducted in the Paraná floodplain, a total of 854 species of macrophytes was compiled. However, the species accumulation curve produced using these studies as a surrogate of sampling effort, did not reach an asymptote (Fig. 5). In fact, the number of species estimated through Jack1 was 1 388, indicating an approximate underestimation of 534 species.
Discussion
According to Chambers et al. (2008), the Neotropical region has the highest number of macrophyte species in the world (984 species). The number of species of macrophytes recorded in the Upper Paraná River and its floodplain (153 species; 16% of the Neotropical region) can be considered high due to the small relative area of this ecosystem (2 500km2) compared to other aquatic areas from this region, such as the Amazon and the Pantanal. Even considering the higher species richness found in the Pantanal Matogrossensse (247 species), our survey still indicates that the Paraná floodplain is highly diverse because the Pantanal is 55 times larger in area, extending over approximately 138 183km2. Thus, the conservation units contained inside this stretch of the Paraná River can be considered important for the conservation of aquatic macrophyte diversity. Again, we emphasize that the number of species we recorded does not represent the real species richness because there are a number of habitats not yet investigated in this stretch.
The comparison of the region investigated in this study with other dammed stretches suggests the importance of the Paraná floodplain as a hotspot of macrophyte species diversity in this basin. In a survey of 18 reservoirs of the Paraná River and some of its main tributaries, Martins et al. (2008) found only 39 species of macrophytes. Even in the Itaipu Reservoir, which is dendritic (and thus, favorable for macrophyte colonization), a long-term dataset showed a total of 110 species (Mormul et al. 2010). The same conclusion can be made when we compare our data with reservoirs from other basins; for example, only 23 species of macrophytes were recorded in the Guri Reservoir, Venezuela (Vilarrubia & Cova 1993). The great variety of habitats found in the Paraná floodplain, together with the natural disturbance caused by seasonal oscillations in the water level, might explain these differences in relation to different reservoirs. On the other hand, the sampling effort was not controlled in these different surveys, and thus the results should be viewed with caution. However, this pitfall might be minimized because all of the investigations that we included in this report were floristic surveys, which tend to maximize the sampling within a region. In addition, there may be differences in the definition of macrophytes used in different surveys. Considering species composition, the assemblages of the Paraná floodplain can be considered as a sample of the aquatic flora from the Pantanal, as was previously pointed out by Thomaz et al. (2009). In fact, the most representative families in number of species are largely the same for both ecosystems (Poaceae and Cyperaceae, Onagraceae, Pontederiaceae, Plantaginaceae and Fabaceae). The families with few species in both ecosystems are also the same (Typhaceae, Cucurbitaceae, Maranthaceae, Haloragaceae, Solanaceae and Orchidaceae). Our cluster analysis also indicated that these three wetlands are the most similar amongst all ecosystems in our dataset, which could be due to their geographical proximity and to hydrological similarities (all areas are subjected to seasonal variation in the water level and also include a great variety of habitats). Furthermore, both wetlands belong to the same larger Paraná basin.
However, some families differ considerably in the number of species between these two wetlands. For example, there are only two species of Nymphaeaceae in the Paraná floodplain, whereas there are eight in the Pantanal that occur mainly in rain- fed shallow ponds and seasonal standing or slow flowing water in addition to in the river floodplain, except for Victoria amazonica, which grows in oxbow lakes. Similarly, 10 species of Characeae were found in the Pantanal, which is attributed to the alkaline and brackish waters in the Southwestern Pantanal (Bueno 1993, Pott & Pott 1997), but only two were identified in the Paraná floodplain, where acid soils predominate and charophytes do not thrive. The importance of the type of habitat in determining species composition can also be observed if we compare a survey carried out on lakes, reservoirs and wetlands in the Southern Paraná State (Cervi et al. 2009), only 200km away, that shares only 23% of macrophytes with the Paraná floodplain.
The low richness of aquatic epiphytes reported in the Paraná floodplain is related to the small sampling effort that has been carried out on floating meadows. Epiphytes usually colonize advanced stages of aquatic succession (Pott & Pott 2003). For example, surveys carried out by Tur (1972) and Neiff (1982) in the Middle Paraná (Argentina) identified 70 species of epiphytes. Even though both regions are on the Paraná River, the flood pulses differ between the Middle and the Upper Paraná basins, which may influence the accumulation of organic matter and, thus, the formation of floating-substrates, as well as the displacement of these islands, and this may explain the differences in the richness of epiphytes found in these wetlands.
The species number increase over time reported for the Paraná floodplain can be mainly attributed to the refinement of the taxonomic searches and identification carried out. In addition to this effect, we also considered a higher sampling effort in the Paraná floodplain, with collections made in habitats not previously investigated (e.g., rocks in the Paraná channel) and the arrival of new species (e.g. Hydrilla verticillata). The high level of species richness that we found indicates that the Paraná floodplain is still in a good conservation state, despite the strong anthropogenic pressures in the region related to changes in hydrometric levels, nutrient cycling and suspended solid loadings (Souza-Filho 2009).
However, despite the good status of conservation with respect to the aquatic flora, we contend that there is a concern related to the presence of two invasive species, H. verticillata and Urochloa subquadripara. The first is a submersed species native to Asia and the North of Africa, that colonizes the Paraná main channel and has a high competitive ability, threatening native species due its rapid regeneration following hydrological disturbances (Sousa et al. 2009, Thomaz et al. 2009). Its success in the Paraná main channel is associated with the same effect leading to an increase in the colonization by submersed species, i.e., the increase in water transparency and propagule pressures originating in the upstream reservoirs (Thomaz et al. 2009). Hydrilla verticillata has not yet colonized lakes in either the Baía or Ivinhema river habitats (Sousa et al. 2009). The second species, U. subquadripara, belongs to the family Poaceae, which contributes with several invasive species (Petenon & Pivello 2008). Although U. subquadripara has been rarely recorded in the Paraná floodplain, it reduced significantly the diversity of macrophytes in a lake close to the Baía River, the only place where it occurs with high biomass in this floodplain (Michelan et al. 2010). Disturbances associated with the oscillation in water levels may explain why this species is so rare in most habitats in the floodplain, but in light of its severe threat to macrophyte diversity, its monitoring is a priority, especially in the best preserved areas of this region.
The results of our cluster analysis indicate that Neotropical wetlands are different regarding macrophyte composition. Thus, we infer that such differences may be due to multiple factors, such as climate, flood regime and geography.
In fact, the most similar areas (Pantanal and Paraná floodplain) share many similarities: they are both large floodplains located from 80-160 m.a.s.l., have a great variety of habitats and are subjected to seasonal water level fluctuations (Agostinho & Zalewiski 1995, Vila da Silva 1995). However, the cluster analysis also shows that South American wetlands are diverse regarding macrophyte assemblages, and even ecosystems located in the same basin may differ considerably (e.g., the upper and middle/lower Paraná floodplains). The differences observed for these two floodplains may be accounted for by differences in their nutrient regimes (the Argentinean floodplains receive high phosphorus inputs from the Andes tributaries) and also to the types of habitats investigated (e.g., the widespread occurrence of floating meadows in Argentina with a high richness of epiphytes). As previously mentioned, the groups formed by the cluster analysis suggest that, though they represent geographically distant environments, such as the Amazonian floodplains and the Argentina plain, the sampled landscapes are determinant in forming groupings.
The accumulation curve reflects the differences found in the cluster analysis. In other words, the great differences among the South American surveys included in the cluster analysis indicate a high beta-diversity, leading to a lack of an asymptote in the accumulation curve. The total number of species found in all 12 surveys represents 87% of the number found by Chambers et al. (2008) for the Neotropical region. Despite the fact that our findings are close to the total number of Neotropical macrophytes, the lack of an asymptote, together with the high underestimation of true richness suggested by the Jack1 estimator, indicates that we are still far from describing the actual richness for this region. The number of plants to be described in Brazil, what may reflect the situation of South America, is considered very high (Pimm et al. 2010). In fact, there has been a clear lack of investigations conducted in pristine habitats in South America, such as in parts of the Amazon and the Andes, which are areas of high biodiversity. Future investigations at these sites, together with the description of new species (e.g., Bove et al. 2006, Amaral & Bittrich 2008), will certainly increase the number of species of Neotropical macrophytes recorded and give a better idea of the biodiversity provided by the great variety of ecosystems of this biogeographical region.
Our results reinforce the hypothesis of Irgang & Gastal (1996) that Uruguay, North Argentina, Paraguay and South Brazil form a phytogeographic unit, and therefore, the sampled number of species does not closely correlate with other evaluated areas. There are many other large wetlands in South America that should be included in this analysis but that were not included because of insufficient floristic inventories, such as Guaporé and Ilha do Bananal.
In summary, this report highlights the flora of different wetlands of South America and indicates that the actual species richness of macrophytes of this continent is far from being well understood. Our hypothesis sustains that macrophyte records, together with existing surveys, indicate a continuous need for carrying out increasing numbers of collections in new areas in the upper Paraná river-floodplain system and in other South American wetlands, as the number of species so far reported remains far from the predicted total. The checklist generated in this study is intended to support other research in wetlands and, in particular, to assure the continuity of ongoing long-term ecological programs, and it reveals a rich flora that is practically unknown to botanists and ecologists.
Acknowledgments
This study was funded by the Brazilian Council of Research (CNPq) through a Long Term Ecological Program (site number 6). S. M. Thomaz received a CNPq Productivity Research Grant and acknowledges this agency for long-term funding. A. Pott received a CNPq and CAPES Productivity Research. Additionally, F. A. Ferreira and R. P. Mormul acknowledge CNPq and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for furnishing Ph.D. fellowships, respectively.
References
Agostinho, A.A. & M. Zalewski. 1995. The dependence of fish community structure and dynamics on floodplain and riparian ecotone zone in Parana River, Brazil. Hydrobiologia 303: 141-148. [ Links ]
Amaral, M.C., V. Bittrich, A.F. Faria, L.A. Anderson & L.Y.S. Aona 2008. Guia de campo para plantas aquáticas e palustres do Estado de São Paulo. Holos, Ribeirão Preto, São Paulo, Brazil. [ Links ]
APG II. 2003. An update of the Angiosperm Phylogeny Group classification for orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 141: 399-436. [ Links ]
Bini, L.M. 1996. Influência do pulso de inundação nos valores de fitomassa de três espécies de macrófitas aquáticas. Braz. Arch. Biol. Techn. 39: 715-721. [ Links ]
Bove, C.P., C.T. Philbrick & A.R. Novelo R. 2006. A new species of Cipoia (Podostemaceae) from Minas Gerais, Brazil. Syst. Bot. 31: 822-825. [ Links ]
Bove, C.P. & J. Paz. 2009. Guia de campo das plantas aquáticas do Parque Nacional da Restinga de Jurubatiba. Museu Nacional, Rio de Janeiro, Rio de Janeiro, Brazil. [ Links ]
Bueno, N.C. 1993. Characeae do Pantanal de Mato Grosso do Sul, Brasil, Levantamento Florístico. Master Thesis. Universidade Federal do Paraná, Curitiba, Paraná, Brazil. [ Links ]
Cervi, A.C., C. Bona, M.C.C. Moço & L. Linsingen. 2009. Macrófitas aquáticas de General Carneiro, Paraná, Brasil. Biota Neotrop. 9: 215-222. [ Links ]
Chambers, P.A., P. Lacoul, K.J. Murphy & S.M. Thomaz. 2008. Global diversity of aquatic macrophytes in freshwater. Hydrobiologia 595: 9-26. [ Links ]
Clarke, K.R. & R.N. Gorley. 2006. Primer v6: User Manual/Tutorial, Primer-E, Plymouth, United Kingdom. [ Links ]
Colwell, R.K. 2009. EstimatesS: statistical estimation of species richness and shared species from samples EstimateS. Version 8.2.0. (Downloaded: February 08, 2010, http://viceroy.eeb.uconn.edu/estimates). [ Links ]
Cook, C.D.K. 1996. Aquatic plant book. Amsterdam and New York, SPB, The Hague, The Netherlands. [ Links ]
Crandall-Stotler, B. 1980. Morphogenetic desings and theory of bryophyte origins and divergence. BioScience 30: 580-585. [ Links ]
Cronk, J.K. & M.S. Fennessy 2001. Wetland plants: biology and ecology. Lewis, Florida, USA. [ Links ]
Diegues A.C. 1994. An Inventory of Brazilian Wetlands. IUCN, Gland, Switzerland. [ Links ]
Esteves, F.A. 1998. Fundamentos de Limnologia. Interciência/FINEP, Rio de Janeiro, Brazil. [ Links ]
Hoehne, F.C. 1948. Plantas aquáticas. Secretaria da Agricultura, São Paulo, São Paulo, Brazil. [ Links ]
Irgang, B.E. & C.V.S. Gastal Jr. 1996. Macrófitas Aquáticas da planície costeira do RS. CPG - Botânica/ UFRGS, Porto Alegre, Rio Grande do Sul, Brazil [ Links ]
Junk, W.J., P.B. Bayley & R.E. Sparks. 1989. The flood pulse concept in river- floodplain systems. Can. J. Fish. Aquat. Sci. 106: 110-127. [ Links ]
Junk, W.J., J.J. Ohly, M.T.F. Piedade & M.G.M. Soares. 2000. The Central Amazon floodplain: Actual use and options for sustainable management. Backhuys, Leiden, The Netherlands. [ Links ]
Kita, K.K. & M.C. Souza. 2003. Levantamento florístico e fitofisionomia da lagoa Figueira e seu entorno, planície alagável do alto rio Paraná, Porto Rico, Estado do Paraná, Brasil. Acta Sci. Biol. Sci. 25: 145-155. [ Links ]
Krebs, C.J. 1999. Ecological methodology. A. Wesley Longman, New York, New York, USA. [ Links ]
Maack, R. 2002. Geografia física do Estado do Paraná. Curitiba, Paraná, Brazil. [ Links ]
Martins, D., N.V. Costa, M.A. Terra & S.R. Marchi. 2008. Caracterização da comunidade de plantas aquáticas de dezoito reservatórios pertencentes a cinco bacias hidrográficas do estado de São Paulo. Planta Daninha 26: 17-32. [ Links ]
Michelan, T.S., S.M. Thomaz, R.P. Mormul & P. Carvalho. 2010. Effects of an exotic invasive macrophyte (tropical signalgrass) on native plant community composition, species richness and functional diversity. Freshwat. Biol. 55: 1315-1326. [ Links ]
Mobot, Missouri Botanical Garden. W Tropicos. (Downloaded: February 08, 2010, http://mobot.mobot.Org/W3T/search/vast.html). [ Links ]
Mormul, R.P., F.A. Ferreira, P. Carvalho, T.S. Michelan, M.J. Silveira & S.M. Thomaz. 2010. Aquatic macrophytes in the large, sub-tropical Itaipu Reservoir. Brazil Rev. Biol. Trop. 58: 1437-1452. [ Links ]
Naranjo, L.G. 1995. An evaluation of the first inventory of South American wetlands. Vegetatio 118: 125-129. [ Links ]
Neiff, J.J. 1978. Fluctuaciones de la vegetacion acuatica en ambientes del valle de inundacion del Paraná medio. Physis 38: 41-53. [ Links ]
Neiff, J.J. 1982. Esquema sucesional de la vegetacion en Islas Flotantes del Chaco Argentino. Bol. Soc. Arg. Bot. 21: 325-341. [ Links ]
Neiff, J.J. 1986. Aquatic plants of the Paraná System, p. 557-571. In B.R. Davies & K.F. Walker (eds.). The Ecology of River Systems. Dr. W. Junk, Dordrecht, Netherlands. [ Links ]
Neiff, J.J. 2001. Diversity in some tropical wetland systems of South America. In B. Gopal, B. Junk, W.J. & J.A. Davies (eds.). Biodiversity in wetland: assessment, function and conservation. Backhuys, Leiden, Netherlands. [ Links ]
Pedralli, G. 1990. Macrófitos aquáticos: técnicas e métodos de estudos. Est. Biol. 26: 5-24. [ Links ]
Petenon, D. & V.R. Pivello. 2008. Plantas invasoras: representatividade da pesquisa dos países tropicais no contexto mundial. Natureza & Conservação 6: 65-77. [ Links ]
Pimm, S.L., C.N. Jenkins, L.N. Joppa, D.L. Roberts & G.J. Russell. 2010. How many endangered species remain to be discovered in Brazil? Natureza & Conservação 8: 71-77. [ Links ]
Pott, V.J. & A. Pott. 1997. Checklist das macrófitas aquáticas do Pantanal, Brasil. Acta Bot. Bras. 11: 315-327. [ Links ]
Pott, V.J. & A. Pott. 2000. Plantas aquáticas do Pantanal. Embrapa, Brasília, Brazil. [ Links ]
Pott, V.J. & A. Pott. 2003. Dinâmica da vegetação aquática do Pantanal, p. 145-162. In S.M. Thomaz & L.M. Bini (eds.). Ecologia e manejo de macrófitas aquáticas. EDUEM, Maringá, Paraná, Brazil. [ Links ]
Pressey, R.L. & P. Adam. 1995. A review of wetland inventory and classification in Australia. Vegetatio 118: 81-101. [ Links ]
R Development Core Team, 2010. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. (Downloaded: February 08, 2010, www.R-project.org). [ Links ]
Sneath, P.H. & R.R. Sokal. 1973. Numerical Taxonomy: The principles and practice of numerical classification. W.H. Freeman, San Francisco, USA. [ Links ]
Souza Filho, E.E. 1993. Aspectos da geologia e estratigrafia dos depósitos sedimentares do rio Paraná entre Porto Primavera (MS) e Guaíra (PR). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil. [ Links ]
Souza-Filho, E.E. 2009. Evaluation of the Upper Paraná River discharge controlled by reservoirs. Braz. J. Biol. 69: 707-716. [ Links ]
Sousa, W.T.Z., S.M. Thomaz, K.J. Murphy, M.J. Silveira & R.P. Mormul. 2009. Environmental predictors of the occurrence of exotic Hydrilla verticillata (L.f.) Royle and native Egeria najas Planch. in a sub-tropical river floodplain: the Upper River Paraná, Brazil. Hydrobiologia 632: 65-78. [ Links ]
Taylor, A.R.D., G.W. Howard & G.W. Begg. 1995. Developing wetland inventories in Southern Africa: A review. Vegetatio 118: 57-79. [ Links ]
Thomaz, S.M., T.A. Pagioro, L.M. Bini & D.C. Sousa. 2004. Aquatic macrophytes from the upper Paraná river floodplain: species list and patterns of diversity in large scale. In A.A. Agostinho, L. Rodrigues, L.C. Gomes, S.M. Thomaz & L.E. Miranda (eds.). Structure and functioning of the Paraná River and is floodplain. EDUEM, Maringá, Paraná, Brazil. [ Links ]
Thomaz, S.M., P. Carvalho, A.A. Padial & J.T. Kobayashi. 2009. Temporal and spatial patterns of aquatic macrophyte diversity in the Upper Paraná River floodplain. Braz. J. Biol. 69: 617-625. [ Links ]
Tur, N.M. 1972. Embalsados y Camalotes de la Región Isleña Del Paraná Médio. Darwiniana 7: 397-407. [ Links ]
Vila da Silva, J.S. 1995. Elementos fisiográficos para delimitação do ecossistema Pantanal: Discussão e proposta, p. 439-458. In F.A. Esteves (ed.) Oecologia Brasiliensis, Estrutura, Funcionamento e Manejo de Ecossistemas Brasileiros. PPGEUFRJ, Rio de Janeiro, Brazil. [ Links ]
Vilarrubia, V.T. & M. Cova. 1993. Estudio sobre la distribucion y ecologia de macrofitos acuaticos en el embalse de Guri. Interciencia 18: 77-82. [ Links ]
Willis, J.C. 1973. A dictionary of the flowering plant and ferns. Cambridge University, Cambridge, England. [ Links ]
Correspondencia a: Fernando Alves Ferreira & Roger Paulo Mormul. Postgraduate Program in Ecology of Continental Aquatic Environments, Universidade Estadual de Maringá- UEM, Av. Colombo 5790, bloco H-90, CEP 87020-900, Maringá, Paraná, Brazil; ferreirabot@gmail.com, roger.mormul@gmail.com
Sidinei Magela Thomaz. Research Group in Limnology, Ichthyology and Aquaculture-Nupelia, Biological Science Department, Universidade Estadual de Maringá-UEM, Av. Colombo 5790, bloco H-90, CEP 87020-900, Maringá, Paraná, Brazil; smthomaz@nupelia.uem.br
Arnildo Pott & Vali Joana Pott Biological Science Department, Universidade Federal do Mato Grosso do Sul - UFMS CEP 79.070-900, Caixa - Postal 549, Campo Grande, MS, Brazil; arnildo.pott@gmail.com, vali.pott@gmail.com
Received 04-VI-2010. Corrected 30-XI-2010. Accepted 07-I-2011.


{kind=link}
{kind=link}
{kind=link}