










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.59 n.1 San José Mar. 2011
Copulatory behaviour and the process of intromission in Anastrepha ludens (Diptera: Tephritidae)
R.D. Briceño1, Dina Orozco2, J. Luis Quintero2, Paul Hanson1 & Ma. del Refugio Hernández2
1. Escuela de Biología, Universidad de Costa Rica; rbriceno@biologia.ucr.ac.cr, phanson@biologia.ucr.ac.cr
2. Programa Moscamed Moscafrut-Desarrollo de Métodos Central Poniente No. 14 altos-Esq. 2ª Avenida Sur. CP 30700. Tapachula, Chiapas, México; dorozco1@prodigy.net.mx
Dirección para correspondencia
Abstract
Complex genitalia occur in many arthropods and in some species extreme female morphologies lead to serious mechanical difficulties for males. Tephritid flies offer examples of such complex genitalia. Because of their economic importance and the extensive use of sterile male releases for tephritid control in Texas and Mexico, studies have been done on various aspects of their basic reproductive biology, but the process of intromission has received little attention. The distiphallus of the male of Anastrepha ludens is complex. One membranous sac on the distiphallus is capable of rhythmic cycles of inflation and deflation. Inflations of the sac near the base of the distiphallus probably help propel the aedeagus deeper into the female along with stiffening of the basiphallus and may drive the genital rod (which does not transfer sperm) into the ventral receptacle. We were unable to establish an association between some of the behaviours displayed by males during mating and intromission process. Rev. Biol. Trop. 59 (1): 291-297. Epub 2011 March 01.
Key words: genitalia, penetration process, copulatory behavior, Anastrepha ludens, fruit flies.
Resumen
En muchos artrópodos, se observan genitales complejos; la morfología extrema en las hembras podría conducir a serios problemas mecánicos para los machos (Kamimura & Matsuo 2001). Las moscas tefrítidas tienen ejemplos de tales órganos genitales complejos. Debido a su importancia económica y la amplia utilización de machos estériles para el control biológico en Texas y México, se han realizado estudios sobre diversos aspectos de su biología reproductiva, pero el proceso de penetración ha recibido muy poca atención. El distiphallus de los machos de Anastrepha ludens es complejo. Un saco membranoso en el distiphallus es capaz de realizar ciclos rítmicos de inflación y deflación. El inflado de la bolsa cerca de la base del distiphallus, junto con la rigidez del basiphallus probablemente ayuda a impulsar el edeagus más profundamente en la hembra y orientar la vara genital (que no transfiere esperma) en la región del receptáculo ventral. No pudimos establecer una asociación entre algunas de las conductas mostradas por los machos durante el apareamiento y el proceso de intromisión.
Palabras claves: genitalia, proceso penetración, comportamiento copulatorio, Anastrepha ludens, mosca de la fruta.
Fruit flies (Tephritidae) encompass over 4 200 species, including the Mediterranean fruit fly, apple maggot, Mexican fruit fly, olive fruit fly and over 60 other species that damage seeds, fruits, stems, etc. They are found on a great variety of tropical, sub-tropical and temperate plants (White & Elson- Harris 1992). The Mexican fruit fly, Anastrepha ludens (Loew) is a polyphagous, frugivorous tephritid occurring from southern Texas to at least Costa Rica (Stone 1942).
Complex genitalia occur in many arthropods and in some species extreme female morphologies lead to serious mechanical difficulties for males (Kamimura & Matsuo 2001). Tephritid flies offer examples of such complex genitalia. Because of their economic importance and the extensive use of sterile male releases for tephritid control in Texas and Mexico, various aspects of their basic reproductive biology have been studied, but the process of intromission has received little attention. Intromission is mechanically challenging in many tephritids because the female vagina is a long, thin, S- shaped tube, a design that allows the female to extend her long ovipositor and lay eggs in sheltered sites. The male’s intromittent organ, his phallus, which includes a long, highly flexible, threadlike distiphallus, must penetrate deep into the female to deposit sperm near the ventral receptacle and the mouth of the spermatecal duct (Marchini et al. 2001, Fritz & Turner 2002). The process that allows such a long flexible structure to be threaded through the female cloaca and vagina to her bursa remains little studied and only in Ceratitis capitata this process is relatively well known (Eberhard 2005). Currently there are no studies on the functional morphology of male genitalia in Anastrepha ludens.
In this report we present a general description of the copulatory behaviour by the male with respect to the intromission process of the genitalia in Anastrepha ludens.
Material and Methods
Mass reared fertile and sterile flies were obtained as pupae from the Moscafrut facility in Mexico. This strain has been maintained for 13 years under mass-rearing conditions. Wild flies from Mexico were raised from larvae that emerged from infested oranges (Citrus aurantium) collected in the Soconusco region, in Chiapas, México. Costa Rican wild flies were collected from infested orange fruits at the Estación Experimental Fabio Baudrit near Alajuela. Third instar larvae were placed in vermiculite (Strong-lite®, Products Corp. Séneca lllinois) and maintained at 23±1°C during 14 days, the time required for pupal maturation. The pupae were later placed in wooden cages (30x30x30cm) covered with mesh (tull 2mm) until emergence. After emergence males and females were placed in separate cages and maintained in laboratory conditions under a photoperiod of 12L:12D (550±50lux), 25±1°C and 65±5% relative humidity. Adult flies from all strains were fed with a 3:1 mixture of sugar and hydrolyzed yeast.
Male-female pairs of mass reared flies, mass reared males-wild females and wild flies from Mexico were placed together and allowed to copulate. Laboratory fertile flies were 10 to 12 days old and wild flies were 18 to 20 days old. Copulating pairs were immobilized nearly instantaneously with a freezing spray (ethyl chloride) and fixed in 70% ethyl alcohol or immersed directly in alcohol. Whole mount preparations of the genital tracts of coupled pairs were examined with the light microscope. By freezing pairs after known periods of time (20, 40, 60, 80 and 100 minutes after the male mounted the female), it was possible to deduce the order of some events during copulation.
Pairs of flies in Costa Rica were videotaped at room temperature in 13.7cm diameter and 1.8cm deep mating chambers (clear Petri dishes) on a glass table using a Sony DCR. TRV80 digital camera equipped with +6 closeup lenses. The camera was below the table, allowing recording from below (most courtship occurred on the ceiling of the mating chamber). The pairs were frozen at different behavioural stages. Pairs were formed by transferring an unmated 30-day old male into a plexiglass cage with a 30-day old virgin female between 15:30-20:00h. If the male did not evert the pleural areas of his abdomen within 15 minutes, the flies were separated. For measurements of lengths and angles of the male genitalia we used Scion Image® software.
Results
Names for external structures are those of Aluja & Norrbom (2000) and the following description emphasizes only those parts that are critical in the process of intromission.
Male genitalia: The long aedeagus (5.2-6.1mm) arises from the ventral surface of the abdomen just anterior to the surstyli. The tip of the aedeagus consists of the complex distiphallus; the rest of the aedeagus is more or less cylindrical basiphallus. Nearly all of the dorsal side of the basiphallus, is strengthened by a pair of chitinous strips, while the ventral side is a stiff membrane with a series of transverse folds. The distiphallus is complex (Fig. 2c). At its basal end is a ventral basal lobe or eversible sac, with small, conical, scale-like teeth covering part of its surface (Fig. 2d). The tip of the distiphallus is covered by a relatively smooth cuticular sheet. In the chitinous sheath there is a trumpet-like sclerotized structure, which extends dorsally from the basal area. In addition, a single rod-like sclerotized structure extends ventrally in the opposite direction. The tip of this rod flares, somewhat like a bell.
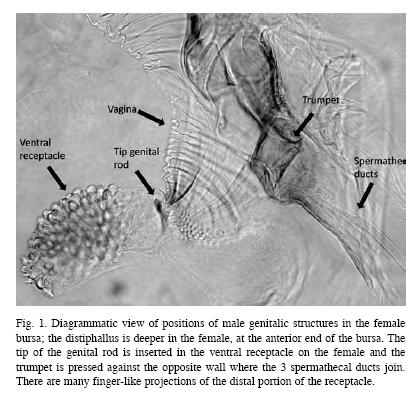
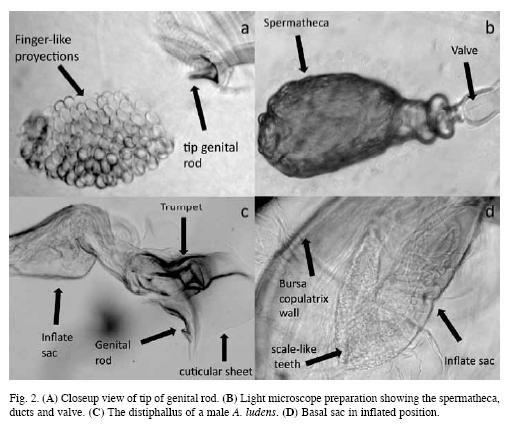
Notable differences in the diameter of the basal sac in slide preparations apparently indicate that it can inflate and deflate. Maximum inflation increased the diameter by an estimated 50-100% beyond that when the sac was minimally inflated. Observing inflated and deflated sacs suggest that the expansion of the basal sac causes a flexion in the region near the basal portion of the distiphallus, changing the direction it is projected. It appears that, as the distal sac is expanded, the trumpet apparently moves in the opposite direction and joins the spermathecal ducts.
Female genitalia: The ovipositor is the most posterior part of the female reproductive tract. The 7th segment is modified into a long, tubular segment within which the ovipositor lies when retracted. The 8th abdominal segment serves as a membranous hinge for extension and retraction of the ovipositor (Dodson 1978). The cloaca is located just anterior to the posterior edge of the sternite and thus lies slightly anterior to the posterior edge of the aculeus. Just prior to intromission, in the three pairs observed, the female extends the ovipositor so that the apical portion of the aculeus is exposed and the male clamps the tip of the aculeus with his surstyli.
Gross anatomical features of the female reproductive system of A. ludens include paired ovaries, lateral oviducts, common oviduct, bursa copulatrix, ventral receptacle, three spermathecae, accessory glands and ovipositor sheath. Two short lateral oviducts originate at the distal end of the ovaries and merge to form the median oviduct, which extends into the ovipositor. The bursa copulatrix is an expanded region that joins with the vagina. The vagina is the widest portion and serves as the point of insertion for the spermathecal ducts and accessory glands.
Projecting anteriorly from the inner upper end of the bursa is the ventral receptacle (fertilization chamber, Salinas & Nuzzaci 1984), a smaller blind- ended, cylindrical structure whose inner-most portion is expanded and covered with smooth, rounded projections. The lining of the bursa is more sclerotized in the area surrounding the entrance to the ventral receptacles and has a truncated conical shape (the receptacle cone) (Figs. 1, 2a).
Three spermathecae are present, a feature common among the higher flies (Chapman 1998). The spermatecae are cuticular vesicles surrounded by convoluted, transparent tissue. As described by Martínez & Hernández-Ortiz (1997) the surface of the spermatecal capsules under the light microscope appears to have numerous sharp, pointed spicules. The spermathecal ducts and those from the accessory glands open into the same small, raised area on the vagina, with the spermathecal ducts arising slightly anterior to the others. The ducts of the accessory glands appear longer than those of the spermatheca. The accessory glands are larger and nearly spherical in shape.
The spermathecae are spherical at their distal end with a narrowing pedicel that is continuous with the spermathecal duct. An expanded region at the base of the spermatheca houses a valve-like structure with a narrow pore; under the light microscope the valve is translucent and membranous (Fig. 2b), similar to that described by Fritz & Turner (2002) in A. suspensa.
Male copulatory behavior during intromission process: Preceding a leap, the male remained oriented toward the female (face to face) in a stationary position. Female orientation is probably the immediate cue triggering mounting attempts by males (Briceño & Eberhard 2002). The male leaps over the head of the female and onto her dorsum, turns and aligns himself, extends his mouth parts, attempts to insert his phallus into her ovipositor, all the while fanning his wings continuously and producing a relatively high intensity, energetic sound (Webb et al. 1976, Sivinsky et al. 1984, Briceño et al. 2009). Direct observations show that just prior to intromission, if the female raises her ovipositor and extends it so that the apical portion of the aculeus is exposed from the eversible membrane, the male can grasp it with his genitalic surstyli. After nipping her ovipositor with his genitalia, the male pulls the female ovipositor and then rubs the base of the female’s ovipositor with his hind legs (movement from bottom to top), an action that represents the first stage of mating. During copulation the position of the male’s legs is as follows: foretarsi along anterior thorax of female; mesotarsi along pleural region of midabdomen of female; metatarsi along oviscape or distal portion of female’s wings. The male also repeatedly pushed his mouthparts over the setae on the female’s head, including the antennal arista.
Typically, the male then appeared to pull posteriorly repeatedly, so that the female’s ovipositor was slightly extended. This extension probably caused the female’s normally S-shaped ovipositor to straighten and perhaps facilitated the movement of the male’s aedeagus toward her bursa. An alternative interpretation, that the female actively extends her ovipositor rather than having it pulled by the male, cannot be discarded, but pulling seems more likely because he often performed repeated "pulling" movements by raising the tip of the abdomen and moving it rearward. Throughout the copulation the male regularly inflates and deflates the anal membrane. The female genitalia have an enlarged dorso basal area of the eversible membrane that receives the enlarged lateral surstylus of the male. The medial surstylus is apparently used to hold the aculeus when she extends it.
We were unable to establish an association between some of the behaviours displayed by males during mating and the intromission process. Certain behaviours were displayed more or less continuously during mating, such as pushing with his mouthparts, rubbing the base of the female’s ovipositor or inflating and deflating the anal gland. One association we could make was that males that were fanning their wings continuously upon mounting the female always had their phallus outside the female’s ovipositor and the pair later separated (n=10). Males that were frozen a few seconds after they stopped vibrating their wings had their genitalia inside the ovipositor or the vagina (n=8).
The internal events associated with intromission were deduced from 161 different pairs frozen at different copulation times in which the aedeagus was at least partially inserted in the female’s vagina. Apparently the intromission process did not follow a strict time table. In 6 males (17%) frozen after 20 minutes the tip of the genital rod was inserted in the ventral receptacle on the female. In males frozen after 80 minutes of copulation 23% (8) were alf way from the receptacle, in two cases (6%) the tip was inserted in the receptacle, and in another two the basiphallus was not in the female bursa. In none of the pairings was a relationship between the length of copulation and distance traveled by the male genitalia into the female genitalia (LabxLab: mean 0.93±0.38mm, r=0.224, p=0.08, n=60; Labx- Wild: 0.82±0.37mm, r=0.06, p=0.52, n=60; WildxWild: 0.75±0.38mm, r=0.05, p=0.720, n=60).
In all 11 cases in which the male genitalia had not entered the bursa and were still inside the ovipositor, the sac of the distiphallus was not inflated. In only two cases where the trumpet and genital rod partially entered the bursa was the sac partially inflated in the widened base of the ovipositor. In all cases in which the male genitalia had entered the bursa, the genital rod was flexed between 1º and 90º. The degree of flexion was related to the degree of inflation of the sac. When it was fully inflated it was flexed 90º or more (48 pairs), moderately inflated between 30-45º (72) and inflated 5º or less (40).
Discussion
The deepest penetration of the male genitalia is to the anterior end of the bursa. The receptacle cone in the bursal lining may serve to guide the rod into the receptacle. Use of inflatable lobes or sacs to drive genitalic structures into the female is also thought to occur in other insects (Siva-Jothy 1988) and in other tephritids (Eberhard & Pereira 2005).
We suggest that sperm are ejaculated through the trumpet into the vagina where the spermatecal ducts arise (Fig. 1). Marchini et al. (2001) have shown in Ceratitis capitata that sperm is ejaculated through two gonopores at the top of the distiphallus and another at the base of the genital rod. The genital rod in A. ludens is a rigid cuticular structure which at its distal end forms just a very short cylinder, without any apparent distal opening through which discharge could occur.
The degree of male genitalic penetration was not directly related to the duration of copulation suggesting that the male genitalia reaching the appropriate site for depositing sperm depends on other factors, such as the interaction between male and female during mating. These results suggest that the duration of copulation is not necessarily a good indicator of how far the male has penetrated into the female genitalia and that differences probably are due to cryptic selection by the female. Another possibility is that the male periodically withdraws his genitalia at least partially from the female, then later introduces them deeper into her reproductive tract again.
Acknowledgments
We thank Eddy Camacho for helping with dissections and slide preparations, and the Universidad de Costa Rica, the Programa Moscamed Moscafrut, SENASICA, SAGARPA, the Mexican government and the International Atomic Energy Agency for financial support.
References
Aluja, M. & A. Norrbom. 2000. Phylogeny of the genera Anastrepha and Toxotrypana (Trypetinae: Toxotrypanini) based on morphology, p. 290-342. In A.L. Norrbom, R.A. Zucchi & V. Hernández-Ortiz. Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior. CRC, Boca Raton, USA. [ Links ]
Briceño, R.D., M.R. Hernández, D. Orozco & P. Hanson. 2009.Acoustic courtship songs in males of Anastrepha ludens (Tephritidae) associated with geography, mass rearing and courtship success. Rev. Biol. Trop. 57: 257-265. [ Links ]
Briceño, R.D. & W. Eberhard. 2002. Decisions during courtship by males and females medflies (Diptera:Tephritidae): coordinated changes in male behavior and female acceptance criteria in massreared flies. Fla. Entomol. 85: 14-31. [ Links ]
Chapman, R.F. 1998. The insects: structures and function, p. 268-321. Cambridge, Cambridge, England. [ Links ]
Dodson, G. 1978. Morphology of the reproductive system in Anastrepha suspensa (Loew) and notes on related species. Fla. Entomol. 61: 231-239. [ Links ]
Eberhard, W. & F. Pereira. 1995. The process of intromission in the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). Phyche 102: 99-119. [ Links ]
Eberhard, W. 2005. Threading a needle with reinforced thread: intromission in Ceratitis capitata (Diptera, Tephritidae). Can. Entomol. 137: 174-181. [ Links ]
Fritz, A.H. & F.R. Turner. 2002. A light and electron microscopical study of the spermathecae and ventral receptacle of Anastrepha suspensa (Diptera: Tephritidae) and implications in female influence of sperm storage. Arthrop. Struct. Dev. 30: 293-313. [ Links ]
Fritz, A.H. 2004. Sperm storage patterns in singly mated females of the Caribbean fruit fly, Anastrepha suspensa (Diptera: Tephritidae) Ann. Entomol. Soc. Am. 97: 1328-1335. [ Links ]
Kamimura, Y. & Y. Matsuo. 2001. A "spare" compensates for the risk of destruction of the elongated penis of earwigs (Insecta: Dermaptera). Naturwissenschaften 88: 468-471. [ Links ]
Marchini, D., G del Bene, L.F. Falso & R. Dallai. 2001. Structural organization of the copulation site in the medfly Ceratitis capitata (Diptera: Tephritidae) and observations on sperm transfer and storage. Arthrop. Struct. Dev. 30: 39-54. [ Links ]
Martínez, M.I. & V. Hernández-Ortiz. 1997. Anatomy of the reproductive system in six Anastrepha species and comments regarding their terminology in Tephritidae (Diptera). Proc. Entomol. Soc. Wash. 99: 727-743. [ Links ]
Salinas, M. & G. Nuzzaci. 1984. Functional anatomy of Dacus oleae Gmel. Female genitalia in relation to insemination and fertilization processes. Entomologica-Bari 19: 135-165. [ Links ]
Siva-Jothy, M. 1988. Sperm "repositioning" in Crocothemis erythraea, a libellulid dragonfly with a brief copulation. J. Insect. Behav. 1: 235-245. [ Links ]
Sivinsky, J., T. Burk & J.C. Webb. 1984. Acoustic courtship signals in the Caribbean fruit fly, Anastrepha suspense (Loew). Anim. Behav. 32: 1011-1016. [ Links ]
Stone, A.1942. The fruit flies of the genus Anastrepha. Department of Agriculture, Washington, USA. [ Links ]
Webb, J.C., J.L. Sharp, D.L. Chambers, J.J. McDow & J.C. Benner. 1976. Analysis and identification of sounds produced by the male Caribbean fruit fly, Anastrepha suspensa. Ann. Entomol. Soc. Am. 69: 415-420. [ Links ]
White, I.M. & M.M. Elson-Harris. 1992. Fruit flies of economic significance: their identification and bionomics. Cambridge, Cambridge, England. [ Links ]
Correspondencia a: R.D. Briceño & Paul Hanson. Escuela de Biología, Universidad de Costa Rica; rbriceno@biologia.ucr.ac.cr, phanson@biologia.ucr.ac.cr
Dina Orozco, J. Luis Quintero & Ma. del Refugio Hernández. Programa Moscamed Moscafrut-Desarrollo de Métodos Central Poniente No. 14 altos-Esq. 2ª Avenida Sur. CP 30700. Tapachula, Chiapas, México; dorozco1@prodigy.net.mx
Received 22-III-2010. Corrected 28-IX-2010. Accepted 26-X-2010.

