










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.58 suppl.3 San José Oct. 2010
Temporal patterns in coral reef, seagrass and mangrove communities from Chengue bay CARICOMP site (Colombia): 1993-2008
Alberto Rodríguez-Ramírez1, Jaime Garzón-Ferreira2, Angélica Batista-Morales3, Diego L. Gil3, Diana Isabel Gómez-López3, Kelly Gómez-Campo3, Tomás López-Londoño3, Raúl Navas-Camacho3, María Catalina Reyes-Nivia1 & Johanna Vega-Sequeda3
1. The University of Queensland, St. Lucia, Brisbane, QLD 4072, Australia; alberto.rodriguez@uq.edu.au,catalina.reyes@uq.edu.au
2. Brewster Academy, 80 Academy Drive, Wolfeboro, NH 03894, USA; anisotremus@gmail.com
3. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; angelica_batista@invemar.org.co, diego.gil@invemar.org.co, digomez@invemar.org.co, kelly_gomez@invemar.org.co, tomas_lopez@invemar.org.co, rnavas@invemar.org.co, co, johanna_vega@invemar.org.coAbstract
Key words: monitoring, coral reef, seagrass, mangrove, Chengue, CARICOMP, Colombian Caribbean.
Resumen
Palabras clave: Monitoreo, arrecifes coralinos, patos marinos, manglares, Chengue, CARICOMP, Caribe colombiano.
Despite their importance to human welfare, these ecosystems have been strongly impacted by anthropogenic activities. Important portions of mangroves around the world are threatened, and present-day mangrove forest areas have declined substantially, with an average loss of 35% (Valiela et al. 2001). Destruction or loss of seagrass meadows have also been reported for most parts of the world; even if natural causes are often related with this condition (e.g. "wasting disease" or high energy storms), the destruction has been commonly associated with human activities due to coastal development and growing populations (Short & Coles 2001, Duarte et al. 2006). Ecological degradation has lead to a decrease between 30 and 60% of seagrass beds around the world (Lotze et al. 2006, Waycott et al. 2009), and locally, some areas of the Colombian Caribbean have lost even more than 90% of seagrasses in the last 70 years (Díaz & Gómez 2003). During the last three decades of the past century, coral reefs have been facing unprecedented changes and mortality events when compared with geological timescales (Aronson et al. 2004, Pandolfi & Jackson 2006, Pandolfi et al. 2006). An unequivocal sign of these changes is the loss of living coral cover which was estimated around 5% per year until 2001 in the Caribbean (Gardner et al. 2003, Côté et al. 2005), 2% per year until 2004 in the Indo-Pacific (Bruno & Selig 2007) and has been significant over the last 40 years in the GBR (Bellwood et al. 2004). Although Colombian coral reefs have not been exempt from this worldwide process, no clear patterns of coral decline have been observed in recent years by monitoring programs (Rodríguez-Ramírez et al. 2010, Zapata et al. 2010). An international cooperative research network to advance understanding of the interactions and importance of the three major coastal ecosystems of the Caribbean was formulated at the beginning of the 1990s. The Caribbean Coastal Marine Productivity Program (CARICOMP) was created by this initiative and monitoring activities began in many countries of the region in 1992 (CARICOMP 1997a, Kjerfve et al. 1998). The Institute of Marine and Coastal Research (INVEMAR) has represented Colombia and contributed to the CARICOMP program through provision of long term monitoring at Chengue Bay (Tayrona Natural Park) since 1992 (Garzón-Ferreira 1998). This site includes sampling stations in coral reef, seagrass and mangrove ecosystems. A second CARICOMP site was established in 1998 in Colombia at San Andrés Island under the responsibility of the Corporation for the Sustainable Development of the San Andrés and Providencia Archipelago (CORALINA). The experience obtained by INVEMAR while participating for several years in CARICOMP was essential for the creation and launch of the National Reef Monitoring System of Colombia (SIMAC). This program has been operating since 1998 without interruption and includes monitoring stations in the Caribbean and the Pacific (Garzón-Ferreira & Rodríguez-Ramírez 2010).
This paper presents a synthesis of the results obtained during 16 years of environmental monitoring in Chengue Bay. Temporal patterns displayed by each studied variable are analyzed and discussed for each of the three ecosystems: coral reef, seagrass beds and mangrove forest.
Materials and methods
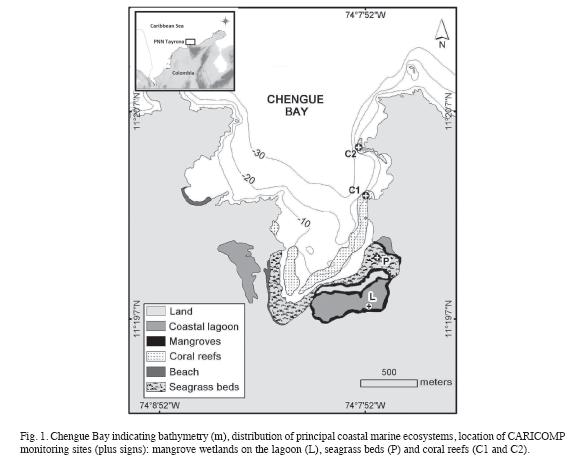
All five species of seagrass beds occurring in Colombia are found in Chengue. Thalassia testudinum beds (where CARICOMP monitoring is conducted) are the most extensive, although restricted to shallow waters in the southern portion of the bay (<3m depth) (Rodríguez-Ramírez & Garzón-Ferreira 2003). Within these beds are found patches of Syringodium filiforme and, less frequently, Halodule wrightii and Halophila baillonis. In deeper waters of the eastern bay wide stands of Halophila decipiens also occur. Thalassia beds grow over calcareous sediments, mainly coarse sand and coral rubble. The calcareous algae Halimeda opuntia is particularly abundant, and in many places, is the dominant live component covering the bottom of Thalassia beds (Garzón-Ferreira & Cano 1991). Chengue exhibits the most important mangrove formation in the national park, which is associated with a coastal lagoon (Fig. 1) surrounded mostly by Red mangrove Rhizophora mangle reaching heights of 13m and densities of 3800 trees/ha (Garzón-Ferreira & Cano 1991, Garzón-Ferreira 1998). Nearby, in less swampy terrain, three additional species are found: Avicennia germinans, Conocarpus erectus, and Laguncularia racemosa. More detailed information about coastal ecosystems in Chengue is provided by Rodríguez-Ramírez & Garzón-Ferreira (2003) and Rodríguez-Ramírez et al. (2004).
Coral reef sampling and data analysis:
The cover of benthic components was estimated annually along five 10 m permanent marked transects on two reef sites (Fig. 1). Transects were sampled using a light chain; components beneath each link were recorded following categories used by CARICOMP (2001). Hard corals were identified to the lowest possible taxonomic level (genus or species). Incidence of coral diseases and bleaching were estimated by examining every hard coral colony (>5cm) within a 2m wide band along each 10m transect. A PVC 1-m pipe marked every 5cm was used as a reference to estimate colony size and band width while swimming at each side of the transect line. Each colony was identified to species level and its surface examined carefully to record the presence of any disease or bleaching. The abundance of important mobile invertebrates (lobsters, octopuses, crabs and sea urchins) was also recorded along each belt transect (10x2m).
Prior to analyses, cover estimates were arcsine transformed. Overall means were calculated by averaging annual means values (averaging all transects per year) by each variable. Temporal trends of coral, algae (pooling turf, fleshy, encrusting and calcareous algae) and abiotic substrate were evaluated by repeated measures ANOVA. We used the univariate approach and the Greenhouse-Geisser corrected probability to test F (adjustment for multiple comparisons: Bonferroni).
Mean values of productivity (g/m2/day), shoot density (shoots/200cm2) and LAI were used to estimate temporal trends. A polynomial model was used to fit the data if R2≥0.70. Pearsons correlations were run to identify relationships between environmental variables and biological data.
Results
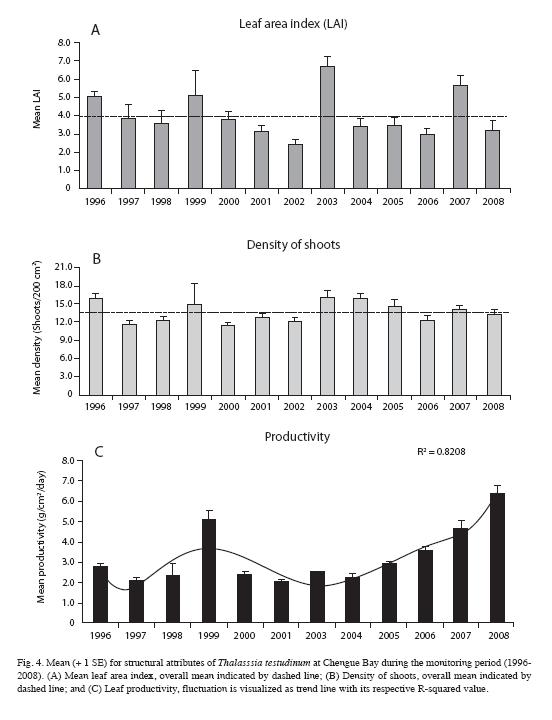
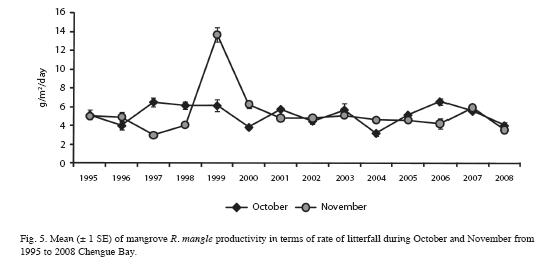
Seagrass beds: Structural parameters of T. testudinum beds on Chengue Bay showed different trends over the monitoring time (Fig. 4); thus: 1) LAI had periodic maximum peaks every 3-4 years, during 1996, 1999, 2003 and 2007; however, analysis showed no time trend (polynomial regression; R2<0.70). Overall mean value for LAI was 4.0±0.5, with fluctuations from 2.4± 0.3 (in 2002) to 6.7±0.5 (in 2003). 2) Density of shoots showed several fluctuations, but not as marked as LAI; maximum peaks were found in 1996, 1999, 2003, 2004 and 2005, and no time trend was evident (polynomial regression; R2 <0.70). Overall mean value for shoot density was 13.5±0.5 shoots/200cm2; with fluctuations from 11.3±0.5 (in 2002) to 16.1±1.1 (in 2003) shoots/200cm2. 3) Productivity was constant during most of the monitoring period (near 2.5 g/m2/day); however, during 1999 an extremely high value was evident, and since 2005 mean values showed a significant progressive increase (R2>0.70). No correlations were identified between environmental variables and biological data (Pearson, all r values between -0.390 and 0.135 and p values >0.05). Means of environmental data during the sampling periods are shown in Table 1.

Discussion
The results from this monitoring program are significant because they document the temporal variation of key structural and functional variables in local coral reefs, seagrass beds and mangroves over a 16 year monitoring period. Furthermore, they constitute a valuable baseline to assess the long-term dynamics of the most important marine ecosystems in the Caribbean.
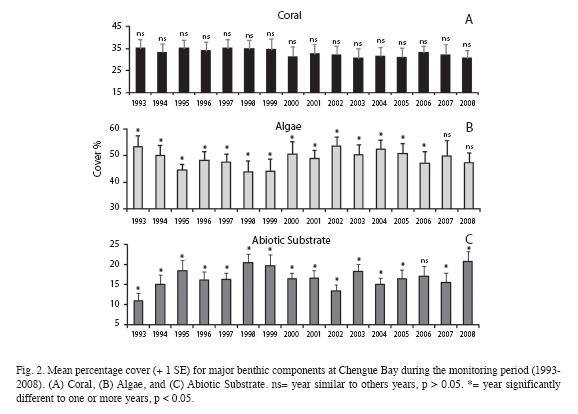
Our findings showed that there is a pattern of stability for coral cover but there is no clear trend in algal cover at monitoring sites between 1993 and 2008. Thus coral cover has remained fairly unchanged during the last 16 years. Previous temporal analyses of coral cover at Chengue found a similar trend between 1993 and 2000 (CARICOMP et al. 2002, Rodríguez- Ramírez & Garzón Ferreira 2003). Although this appears to conflict with two meta-analyses pointing out significant trends of decline in the Caribbean region, in particular the loss of coral cover and/or increase in algae abundance (Gardner et al. 2003, Côté et al. 2005), recent studies on coral reef dynamics in the Caribbean and Florida Keys have documented similar patterns of stability in coral cover at local scales (CARICOMP et al. 2002, Roggers & Miller 2006, Edmunds & Elahi, 2007, Somerfield et al. 2008, Rodríguez-Ramírez et al. 2010). The minor reduction registered in coral cover was due to Hurricane Lenny, which touched marginally the north coast of South America by November 14-16, 1999 (Rodríguez-Ramírez & Garzón-Ferreira 2003). However, the lack of changes in coral cover at the monitoring sites does not mean that these reefs may not have been under a process of decline. Temporal analyses of coral species show that significant losses have occurred in other reef assemblages in Chengue (Rodríguez-Ramírez et al. 2010).
Although previous analysis had revealed a decreasing trend in algal cover between 1993 and 1999 for monitoring sites in Chengue (CARICOMP et al. 2002, Rodríguez-Ramírez & Garzón-Ferreira 2003), our results showed that there is no overall pattern of decrease or increase from 1993 to 2008 (Fig. 2B). Clearly, algal variations were not related to changes in coral cover. Instead they reflected transitions with the abiotic substrate. According to Rodríguez-Ramírez & Garzón-Ferreira (2003) this concomitant variation between algae and abiotic substrate may represent some sampling bias laying out the chain on the substratum. On the other hand, algal variations could be associated with seasonal changes because surveys have been done during the rainy season (October-December) where algal communities experience noticeable changes in composition and abundance (Díaz-Pulido & Garzón-Ferreira 2002). However further analyses are required to explain fully the relationship between algal and abiotic substrate variations.
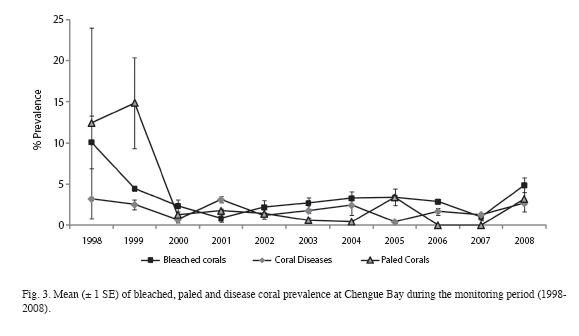
During the study period, Chengue reefs suffered from bleaching (in 1995, 1998, 1999, 2005 and 2008) and have been affected by coral diseases. Earlier studies recognized these stress factors caused minor coral mortality (CARICOMP 1997b, Rodríguez-Ramírez & Garzón-Ferreira 2003, Gil-Agudelo et al. 2006, Rodríguez-Ramírez et al. 2008). In general, the prevalence of bleached corals remained below 4% and paled corals below 2%, which is considered normal for the Caribbean (Weil et al. 2006). In 2005, a major bleaching event occurred in the Caribbean region (Wilkinson & Sauter 2008), but corals in Chengue Bay showed only a slight increase in the average of bleaching and paled corals (both close to 3.5%). This event did not have an effect in this area as large as in other places, probably due to local seasonal upwelling (Rodríguez-Ramírez et al. 2008). Regarding coral diseases, all the most important coral diseases reported for the Caribbean (Black Band Disease, White Band Disease, Yellow Band Disease; see Garzón- Ferreira et al. 2001, Weil et al. 2006) were observed in Chengue, but the most prevalent coral diseases were Dark Spots Disease and White Plague. The prevalence of coral diseases remained relatively low and stable during these 10 years of monitoring, usually staying below 2% and increasing to over 3% (in 1998 and 2001). Thus, the bleaching and diseases data suggest that, in comparison with other Caribbean coral reefs, Chengue reefs have exhibited an important degree of resistance to mass bleaching events and coral diseases.
Because there are no historical records about the abundance of mobile invertebrates of commercial value before the beginning of CARICOMP activities, it is not possible to assess if these organisms have suffered from overfishing or if their low density is a natural condition in Chengue. However, during the years of the monitoring program it has been possible to observe that Chengue is frequently visited by fishermen divers who capture lobsters, crabs, octopuses and other large invertebrates.
Sea urchins results are consistent with composition and density patterns found elsewhere in the Caribbean by CARICOMP (CARICOMP et al. 2002, Linton & Fisher 2004) and support previous findings for these invertebrates at Chengue (Rodríguez-Ramírez & Garzón-Ferreira 2003, Rodríguez-Ramírez et al. 2006). Although few urchins were recorded in the monitoring transects, important populations of these organisms (including D. antillarum) have been observed is shallow reefs of Chengue. This suggests a depth-related spatial distribution of urchins that is probably controlled by the competitive interactions with the damselfish Stegastes planifrons. The territories of this fish species are abundant at the depth interval where monitoring transects are located in Chengue (Santodomingo et al. 2002, Rodríguez-Ramírez et al. 2006). Herbivorous sea urchins, such as D. antillarum and Echinometra viridis, are aggressively excluded from damselfish territories (see Ceccarelli et al 2001 for review).
Since no significant correlation was foundbetween environmental data (temperature, water transparency and salinity) and biological parameters of T. testudinum beds (density of shoots, net above-ground productivity and LAI), it is assumed that fluctuations detected for some structural parameters are better explained by cyclical renewal tendencies more than a response of the community to interannual environmental changes. However samplings along the year are required to establish potential relationships between environmental variables and biological data because our results are constrained to a single period during the year.
Monitoring results may characterize the seagrass beds on Chengue Bay as a lush, healthy and productive ecosystem on the Colombian Caribbean. Its inclusion in a legally protected area with low human intervention, moderate continental influx, seasonal upwelling and its proximity to productive mangroves and coral reefs, are special characteristics that must favour the current condition of these seagrass beds.
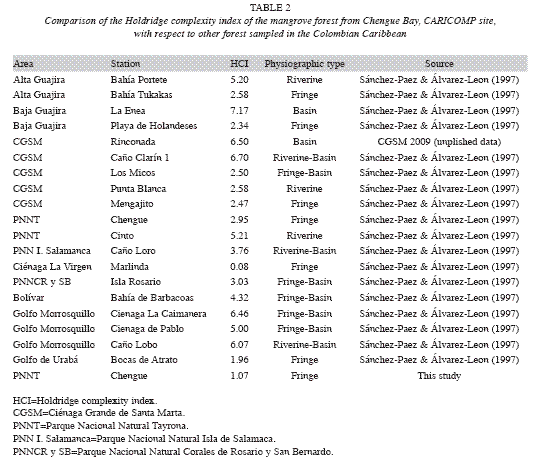
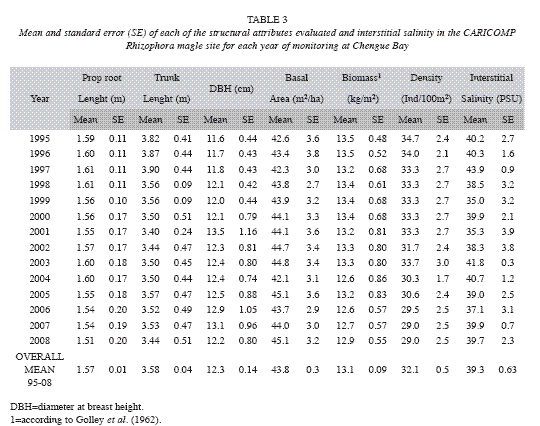
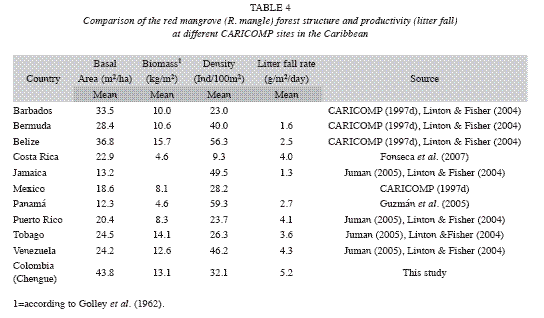
Mangrove: The Red mangrove forest in Chengue Bay has a low structural complexity compared with other mangrove forests along the Colombian Caribbean (Table 2). Relatively small tree sizes, poorly defined wood, very intricate and branched forest (Garzón-Ferreira 1998) are essential characteristics of the site. These features are related to deficiencies in water flow, high salt content and soil conditions (Table 3). In contrast with the mean structural attributes of some of these variables in other mangrove forests in the Caribbean (Table 4), most of the data in this study are close to the highest values reported for other CARICOMP network locations. The results showed that no major structural changes have occurred since 1995 (Table 3). However, variations in biomass, basal area and density, were due mainly to the death of 18% of trees assessed inside the plots. This may have been caused by insect damage (Rodríguez-Ramírez & Garzon-Ferreira 2003). Attacks from xylophagous insects and other pests of some trees are part of the natural mangroves cycles that do not have a major influence on mangrove populations (Villalobos et al. 1996). Human intervention could not have affected directly the structural development of the forest, considering that no evidence of human disturbance was observed on the sample plots. At global level, the mangrove ecosistema is severely threatened and has lost at least 35% of the area of mangrove forests in the past two decades. This is due partly to global changes, but mainly to human activities such as construction, mariculture and agriculture (Valiela et al. 2001, Halpern et al. 2007). In this context, it is suggested that the mangrove in Chengue differs from this scenario because it has not undergone major disturbances over the period of the study, with some alterations primarily influenced by natural degradation factors.
In summary, we did not find evidence of decline trends in the coral reef, seagrass and mangrove sites at Chengue over the monitoring period. Annual variations in coral reefs and mangroves were related to the effects of an isolated event, namely Hurricane Lenny. The observed patterns in seagrasses appear to characterize the natural variability of cyclical renewal processes. Therefore, our results suggest that monitoring sites at the three ecosystems have had a healthy development in the last 16 years. This information is a unique upport for management and conservation purposes at Tayrona Natural Park. As Chengue ecosystems can be critical for ecological processes and also tourism benefits in the region, an effective enforcement on the marine ecosystems inside the Tayrona Park is required.
Acknowledgments
This work has been funded in great part by grants from COLCIENCIAS, Ministerio del Medio Ambiente, Vivienda y Desarrollo Territorial (MAVDT), the Regional Coordinating Unit of the Caribbean Environment Programme of the United Nations Environment Programme (UNEP-CAR/RCU) and CARICOMP. The Instituto de Investigaciones Marinas y Costeras (INVEMAR) has provided most of the required logistics and indirect costs. Thanks to Francisco Arias (INVEMAR), John Ogden, Debby Haynes, Sandy Vargo and Dean Milliken (Florida Institute of Oceanography), Dulcie Linton (Data Management Center-Univ. West Indies, Jamaica) and Alessandra Vanzella (UNEP). Numerous colleagues and students from INVEMAR and other institutions have participated directly in the monitoring activities at Chengue; without their assistance, this study would not have been possible. We finally would like to thank four anonymous reviewers for their constructive criticisms which reatly improved the manuscript. Contribution No. CTRB-1055 from INVEMAR.
Aronson, R.B., I.G. MacIntyre, C.M. Wapnick & M.W. O´Neill. 2004. Phase shifts, alternative states, and the unprecedented convergence of two reef systems. Ecology 85: 1876-1891. [ Links ]
Bellwood, D.R., T.P. Hughes, C. Folke & M. Nystrom. 2004. Confronting the coral reef crisis. Nature 429: 827-833. [ Links ]
Bruno, J. & E.R. Selig. 2007. Regional decline of coral cover in the Indo-Pacific: Timing, extent, and subregional comparisons. PLoS ONE 2: e711. [ Links ]
Bula-Meyer, G. 1977. Algas marinas bénticas indicadoras de un agua afectada por aguas de surgencia frente a la costa Caribe de Colombia. An. Inst. Invest. Mar. Punta Betín 9: 45-71. [ Links ]
CARICOMP. 1997a. Caribbean Coastal Marine Productivity (CARICOMP): A research and monitoring network of marine laboratories, parks and reserves. Proc. 8th Int. Coral Reef Symp. 1: 641-646. [ Links ]
CARICOMP. 1997b. Studies on Caribbean coral bleaching, 1995-96. Proc. 8th Int. Coral Reef Symp. 1: 673-678. [ Links ]
CARICOMP. 1997c. Variation in ecological parameters of Thalassia testudinum across the CARICOMP network. Proc. 8th Int. Coral Reef Symp. 1: 663-668. [ Links ]
CARICOMP. 1997d. Structure and productivity of mangrove forests in the greater Caribbean region. Proc. 8th Int. Coral Reef Symp. 1: 669-672. [ Links ]
CARICOMP. 2001. CARICOMP Methods Manual, Levels 1 and 2: Manual of methods for mapping and monitoring of physical and biological parameters in the coastal zone of the Caribbean. CARICOMP Data Management Center, University of West Indies, Jamaica. [ Links ]
CARICOMP. 2002. Status and temporal trends at CARICOMP coral reef sites. Proc. 9th Int. Coral Reef Symp. 1: 325-330. [ Links ]
Ceccarelli, D.M., G.P. Jones & L.J. McCook. 2001. Territorial damselfishes as determinants of the structure of benthic communities on coral reefs. Oceanogr. Mar. Biol. Annu. Rev. 39: 355-389. [ Links ]
Côté, I.M., J.A. Gill, T.A. Gardner & A.R. Watkinson. 2005. Measuring coral reef decline through metaanalyses. Phil. Trans. R. Soc. B. 360: 385-395. [ Links ]
Díaz, J.M. & D.I. Gómez. 2003. Cambios históricos en la distribución y abundancia de praderas de pastos marinos en la bahía de Cartagena y áreas aledañas (Colombia). Bol. Invest. Mar. Cost. 32: 57-74. [ Links ]
Dorenbosch, M., M.C. van Riel, I. Nagelkerken & G. van der Velde. 2004. The relationship of reef fish densities to the proximity of mangrove and seagrass nurseries. Estuar. Coast. Shelf Sci. 60: 37-48. [ Links ]
Duarte, C.M. & C.L. Chiscano. 1999. Seagrass biomass and production: a reassessment. Aquat. Bot. 65:159-174. [ Links ]
Duarte, C.M., J.W. Fourqurean, D. Krause-Jensen & B. Olesen. 2006. Dynamics of seagrass stability and change, p. 271-294. In A.W.D. Larkum, R.J. Orth & C.M. Duarte (eds.). Seagrasses: Biology, ecology and conservation. Springer, Amsterdam, Netherlands. [ Links ]
Edmunds, P.J. & R. Elahi. 2007. The demographics of a 15-year decline in cover of the Caribbean reef coral Montastraea annularis. Ecol. Monogr. 77: 3-18. [ Links ]
Fonseca, A.C., J. Cortés & P. Zamora. 2007. Monitoreo del manglar de Gandoca, Costa Rica (sitio CARICOMP). Rev. Biol. Trop. 55: 23-31. [ Links ]
Gardner, T.A., I.M. Côté, J.A. Gill, A. Grant & A.R. Watkinson. 2003. Long-term region-wide declines in Caribbean corals. Science 301: 958-960. [ Links ]
Garzón-Ferreira, J. 1998. Bahía de Chengue, Parque Natural Tayrona, Colombia, p. 115-126. In B. Kjerfve (ed.). CARICOMP-Caribbean coral reef, seagrass and mangrove sites. Coastal region and small island papers 3, UNESCO, Paris, France. [ Links ]
Garzón-Ferreira, J. & M. Cano. 1991. Tipos, distribución, extensión y estado de conservación de los ecosistemas marinos del Parque Nacional Natural Tayrona. Final Report, INVEMAR, Santa Marta, Colombia. [ Links ]
Garzón-Ferreira J. & A. Rodríguez-Ramírez. 2010. SIMAC: Development and implementation of a coral reef monitoring network in Colombia. Rev. Biol. Trop. 58: 67-80. [ Links ]
Gil-Agudelo, D.L., J. Garzón-Ferreira, A. Rodríguez- Ramírez, M.C. Reyes-Nivia, R. Navas-Camacho, D.E. Venera-Pontón, G. Díaz-Pulido & J.A. Sánchez. 2006. Blanqueamiento Coralino en Colombia Durante el Año 2005, p. 51-58. In INVEMAR (ed.). Informe del Estado de los Ambientes Marinos y Costeros en Colombia: Año 2005. Serie de publicaciones periódicas INVEMAR No. 8, Santa Marta, Colombia. [ Links ]
Golley, F., H.T. Odum & R.F. Wilson. 1962. The structure and metabolism of a Puerto Rican red mangrove forest in May. Ecology 43: 9-19. [ Links ]
Granek, E., J. Compton & D. Phillips. 2009. Mangroveexported nutrient incorporation by sessile coral reef invertebrates. Ecosystems 12: 462-472. [ Links ]
Harborne, A.R., P.J. Mumby, F. Micheli, C.T. Perry, C.P. Dahlgren, K.E. Holmes & D.R. Brumbaugh. 2006. The functional value of caribbean coral reef, seagrass and mangrove habitats to ecosystem processes. Adv. Mar. Biol. 50: 57-189. [ Links ]
Halpern, B.S., K.A. Selkoe, F. Micheli & C.V. Kappel. 2007. Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conserv. Biol. 21: 1301-1315. [ Links ]
Holdridge, L., W.C. Grenke, W.H. Hatheway, T. Liang & J.A. Tosi. 1971. Forest Environment in Tropical Life Zones. Pergamon, New York, USA. Juman, R. 2005. Biomass, litterfall and decomposition rates for the fringed Rhizophora mangle forest lining the Bon Accord Lagoon, Tobago. Rev. Biol. Trop. 53: 207-217. [ Links ]
Kramer, P.A. 2003. Synthesis of coral reef health indicators for the western Atlantic: Results of the AGRRA Program (1997-2000). Atoll Res. Bull. 496: 1-57. [ Links ]
Linton, D. & T. Fisher (eds.). 2004. CARICOMP. Caribbean Coastal Marine Productivity Program. 1993-2003. CARICOMP, Kingston, Jamaica. [ Links ]
Lotze, H.K., H.S. Lenihan, B.J. Bourque, R.H. Bradbury, R.G. Cooke, M.C. Kay, S.M. Kidwell, M.X. Kirby, C.H. Peterson & J.B.C. Jackson. 2006. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312: 1806-1809. [ Links ]
Mora, C., S. Andrefouet, M.J. Costello, C. Kranenburg, A. Rollo, J. Veron, K.J. Gaston & R.A. Myers. 2006. Coral reefs and the global network of marine protected areas. Science 312: 1750-1751. [ Links ]
Mumby, P.J., A.J. Edwards, J.E. Arias-Gonzalez, K.C. Lindeman, P.G. Blackwell, A. Gall, M.I. Gorczynska, A.R. Harborne, C.L. Pescod, H. Renken, C.C.C. Wabnitz & G. Llewellyn. 2004. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature 427: 533-536. [ Links ]
Nagelkerken, I., G. van der Velde, M.W. Gorissen, G.J. Meijer, T. van´t Hof & C. den Hartog. 2000. Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuar. Coast. Shelf S. 51: 31-44. [ Links ]
Ogden, J.C. 1988. The influence of adjacent systems on the structure and function of coral reefs. Proc. 6th Int. Coral Reef Symp. 1: 123-129. [ Links ]
Ogden, J. & E.H. Gladfelter. 1983. Coral reefs, seagrass beds and mangroves: their interaction in the coastal zones of the Caribbean. UNESCO Rep. Mar. Sci. 23: 1-133. [ Links ]
Pandolfi, J.M., & J.B.C. Jackson. 2006. Ecological persistence interrupted in Caribbean coral reefs. Ecol. Lett. 9: 818-826. [ Links ]
Pandolfi, J.M., R.H. Bradbury, E. Sala, T.P. Hughes, K.A. Bjorndal, R.G. Cooke, D. McArdle, L. McClenachan, M.J.H. Newman, G. Paredes, R.R. Warner & J.B.C. Jackson. 2003. Global trajectories of the long-term decline of coral reef ecosystems. Science 301: 955-957. [ Links ]
Powell, G.V.N., W.J. Kenworthy & J.W. Fourqurean. 1989. Experimental evidence for nutrient limitation of seagrass growth in a tropical estuary with restricted circulation. Bull. Mar. Sci. 44: 324-340. [ Links ]
Prahl, H. von. 1990. Manglares. Villegas, Bogotá, Colombia. [ Links ]
Ramírez, G. 1983. Características físico-químicas de la bahía de Santa Marta (agosto 1980-Julio 1981). An. Inst. Inv. Mar. Punta de Betín 13: 111-121. [ Links ]
Rodríguez-Ramírez, A. & J. Garzón-Ferreira. 2003. Monitoreo de arrecifes coralinos, pastos marinos y manglares en la Bahía de Chengue (Caribe colombiano): 1993-1999. INVEMAR. Serie de Publicaciones Especiales No. 8, Santa Marta, Colombia. [ Links ]
Rodríguez-Ramírez, A. & M.C. Reyes-Nivia. 2008. Evaluación rápida de los arrecifes del huracán Beta en la Isla Providencia (Caribe colombiano). Bol. Invest. Mar. Cost. 37: 217-224. [ Links ]
Rodríguez-Ramírez, A., J. Nivia-Ruiz & J. Garzón-Ferreira. 2004. Características estructurales y funcionales del manglar de Avicennia germinans en la bahía de Chengue (Caribe Colombiano). Bol. Invest. Mar. Cost. 33: 223-244. [ Links ]
Rodríguez-Ramírez, A., C. Bastidas, S. Rodríguez, Z. Leão, R. Kikuchi, M. Oliveira, D. Gil, J. Garzón- Ferreira, M.C. Reyes-Nivia, R. Navas-Camacho, N. Santodomingo, G. Diaz-Pulido, D. Venera-Ponton, L. Florez-Leiva, A. Rangel-Campo, C. Orozco, J.C Márquez, S. Zea, M. López-Victoria, J.A. Sánchez & M.C. Hurtado. 2008. The effects of coral bleaching in the Southern Tropical America: Brazil, Colombia, and Venezuela, p 105-114. In C. Wilkinson & D. Souter (eds.). Status of Caribbean Coral Reefs After the Bleaching and Hurricanes of 2005. GCRMN and RRRC, Townsville, Australia. [ Links ]
Rodríguez-Ramírez, A., M.C. Reyes-Nivia, S. Zea, P. Herron, R. Navas-Camacho, J. Garzón-Ferreira, S. Bejarano & C. Orozco. 2010. Recent dynamics and condition of coral reefs in the Colombian Caribbean. Rev. Biol. Trop. 58: 107-131. [ Links ]
Salazar-Vallejo, S.I. 2002. Huracanes y biodiversidad costera tropical. Rev. Biol. Trop. 50: 415-428. [ Links ]
Santodomingo, N., A. Rodríguez-Ramírez & J. Garzón-Ferreira. 2002. Territorios del pez Stegastes planifrons en formaciones coralinas del Parque Nacional Natural Tayrona, Caribe Colombiano: un panorama general. Bol. Invest. Mar. Cost. 31: 65-84. [ Links ]
Short, F. & R. Coles (eds.). 2001. Global Seagrass Research Methods. Elsevier Science B.V., Amsterdam, Netherlands. [ Links ]
Solano, O.D. 1987. Estructura y diversidad de la comunidad de corales hermatípicos en la Bahía de Chengue. Tesis de Maestría, Universidad Nacional, Bogotá, Colombia. [ Links ]
Valiela, I., J.L. Bowen & J.K. York. 2001. Mangrove forests: One of the world´s threatened major tropical environments. BioScience 51: 807-815. [ Links ]
Waycott, M., C.M. Duarte, T.J.B. Carruthers, R.J. Orth, W.C. Dennison, S. Olyarnik, A. Calladine, J.W. Fourqurean, K.L. Heck, A.R. Hughes, G.A. Kendrick, W.J. Kenworthy, F.T. Short & S.L. Williams. 2009. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 106: 12377-12381. [ Links ]
Villalobos, R, J. Justavino, D. Henríquez, O. Ruiz, A. González, H. Mendoza, H. Barrios, C. Korytkowski & A. Aiello. 1996. Plagas y enfermedades de los manglares. Instituto Nacional de Recursos Naturales Renovables. Organización Internacional de las Maderas Tropicales. [ Links ]
Weil, E., G.W. Smith & D.L. Gil-Agudelo. 2006. Status and progress in coral reef disease research. Dis. Aquat. Org. 69: 1-7. [ Links ]
Werding, B. & H. Erhardt. 1976. Los corales (Anthozoa y Hydrozoa) de la Bahía de Chengue en el Parque Nacional Tayrona (Colombia). Mitt. Inst. Colombo-Alemán Invest. Cient. Punta de Betín 8: 45-57. [ Links ]
Wilkinson, C. & D. Souter (eds.). 2008. Status of Caribbean Coral Reefs After the Bleaching and Hurricanes of 2005. GCRMN and RRRC, Townsville, Australia. [ Links ]
Correspondencia: Alberto Rodríguez-Ramírez. The University of Queensland, St. Lucia, Brisbane, QLD 4072, Australia; alberto.rodriguez@uq.edu.au,catalina.reyes@uq.edu.au
Jaime Garzón-Ferreira. Brewster Academy, 80 Academy Drive, Wolfeboro, NH 03894, USA; anisotremus@gmail.com
Angélica Batista-Morales. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; angelica_batista@invemar.org.co
Diego L. Gil. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; diego.gil@invemar.org.co,
Diana Isabel Gómez-López. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; digomez@invemar.org.co
Kelly Gómez-Campo. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; kelly_gomez@invemar.org.co
Tomás López-Londoño. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; tomas_lopez@invemar.org.co
Raúl Navas-Camacho. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; rnavas@invemar.org.co
María Catalina Reyes-Nivia1 & Johanna Vega-Sequeda. Instituto de Investigaciones Marinas y Costeras INVEMAR. A.A 1016 Santa Marta, Colombia; johanna_vega@invemar.org.co

