










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.58 suppl.3 San José Oct. 2010
Monitoring coral reefs, seagrasses and mangroves in Costa Rica (CARICOMP)
Jorge Cortés1, 2, Ana C. Fonseca1, 3, Jaime Nivia-Ruiz1, Vanessa Nielsen-Muñoz1, Jimena Samper-Villarreal1, Eva Salas1, Solciré Martínez1 & Priscilla Zamora-Trejos1
1. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280; jorge.cortes@ucr.ac.cr
2. Escuela de Biología, Universidad de Costa Rica, 11501-2060 San José, Costa Rica
3. World Wildlife Fund (WWF), San José, Costa Rica, Interlink #718, P.O. Box 02-5635, Miami, Florida 33102, USA.
The coral reefs, seagrasses and mangroves from the Costa Rican Caribbean coast have been monitored since 1999 using the CARICOMP protocol. Live coral cover at Meager Shoal reef bank (7 to 10m depth) at the Parque Nacional Cahuita (National Park), increased from 13.3% in 1999, to 28.2% in 2003, but decreased during the next 5 years to around 17.5%. Algal cover increased significantly since 2003 from 36.6% to 61.3% in 2008. The density of Diadema antillarum oscillated between 2 and 7ind/m2, while Echinometra viridis decreased significantly from 20 to 0.6ind/m2. Compared to other CARICOMP sites, live coral cover, fish diversity and density, and sea urchin density were low, and algal cover was intermediate. The seagrass site, also in the Parque Nacional Cahuita, is dominated by Thalassia testudinum and showed an intermediate productivity (2.7±1.15 g/m2/d) and biomass (822.8±391.84 g/m2) compared to other CARICOMP sites. Coral reefs and seagrasses at the Parque Nacional Cahuita continue to be impacted by high sediment loads from terrestrial origin.The mangrove forest at Gandoca, within the Refugio Nacional de Vida Silvestre Gandoca-Manzanillo (National Wildlife Refuge), surrounds a lagoon and it is dominated by the red mangrove, Rhizophora mangle. Productivity and flower production peak was in July. Biomass (14kg/m2) and density (9.0±0.58 trees/100m2) in Gandoca were relatively low compared to other CARICOMP sites, while productivity in July in Costa Rica (4g/m2/d) was intermediate, similar to most CARICOMP sites. This mangrove is expanding and has low human impact thus far. Management actions should be taken to protect and preserve these important coastal ecosystems. Rev. Biol. Trop. 58 (Suppl. 3): 1-22. Epub 2010 October 01.
Key words: Cahuita, Gandoca, Caribbean, Costa Rica, coral reef, productivity, Thalassia testudinum, Rhizophora mangle, Diadema antillarum, mangrove, seagrass, CARICOMP.
Resumen
Los arrecifes coralinos, pastos marinos y manglares de la costa Caribe de Costa Rica han sido monitoreados desde 1999 siguiendo el protocolo de CARICOMP. La cobertura de coral vivo en el arrecife de Meager Shoal (7 a 10m de profundidad) en el Parque Nacional Cahuita, aumentó de 13.3% en 1999, a 28.2% en 2003, pero después bajó, por los siguientes 5 años, a aproximadamente 17.5%; la cobertura de algas aumentó significativamente de 36.6% en 2003 a 61.3% en 2008. La densidad de Diadema antillarum osciló entre 2 y 7 ind/m2 mientras que Echinometra viridis decreció significativamente de 20 a 0.6 ind/m2. Comparado con otros sitios CARICOMP, la cobertura de coral vivo, diversidad y densidad de peces, y densidades de erizos de mar fueron bajas y la cobertura algal intermedia. El sitio de pastos marinos, también en el Parque Nacional Cahuita, está dominado por Thalassia testudinum y tiene una productividad (2.7±1.15g/m2/d) y biomasa (822.8±391.84g/m2) intermedia comparado a otros sitios CARICOMP. Los arrecifes coralinos y pastos marinos en el Parque Nacional Cahuita continúan siendo impactados por sedimentos terrígenos. El bosque de manglar en Laguna Gandoca, dentro del Refugio Nacional de Vida Silvestre Gandoca-Manzanillo, está bordeado por una laguna y predomina el mangle rojo, Rhizophora mangle. El pico de productividad y producción de flores fue en julio. La biomasa (14 kg/m2) y densidad (9.0±0.58 árboles/100 m2) en Gandoca fueron relativamente bajas comparadas con otros sitios CARICOMP, mientras que la productividad en julio en Costa Rica (4g/m2/d) fue intermedia, similar a la mayoría de los sitios CARICOMP. Este manglar se está expandiendo y tiene muy poco impacto humano hasta ahora. Se deben tomar acciones de manejo para proteger y preservar estos importantes ecosistemas costeros.
Palabras claves: Cahuita, Gandoca, Caribe, Costa Rica, cobertura de coral, productividad, Thalassia testudinum, Rhizophora mangle, Diadema antillarum, floración de Thalassia, CARICOMP.
Materials and methods
Site descriptions
The climate of the Caribbean coast of Costa Rica consists of two rainy seasons, November to March and June to August. Annual rainfall rate in this southern section of the Caribbean coast is lower (2 500mm) than in the northern section (6 000mm). The micro-climate within REGAMA is characterized by nightly rains and ample periods of sunlight during the day (Herrera 1984, Alfaro 2002). Tides are mixed, mainly diurnal, and range between 30 to 50cm (Lizano 2006). Wave direction depends on the position of the Intertropical Convergence Zone and is mainlyfrom the northeast between January and June, and from the east from July to December (Lizano 2007). Currents flow from northwest to southeast, with small eddies in the opposite direction (Cortés 1994), which transport terrestrial sediments and contaminants derived from deforested and urbanized lands in upstream watersheds and coasts (Cortés 1981, 1994, Cortés & Risk 1985, Fonseca & Cortés 2002). These sediments have been the main cause of coral reef and seagrass degradation during the last 50 years, combined with other increasing natural and anthropogenic disturbances (Cortés et al. 1992, 1994, Fonseca & Cortés 2002, Cortés & Jiménez 2003, Fonseca 2003, Roder 2005, Roder et al. 2009).
Coral reef and seagrasses of Cahuita National Park (PNC)
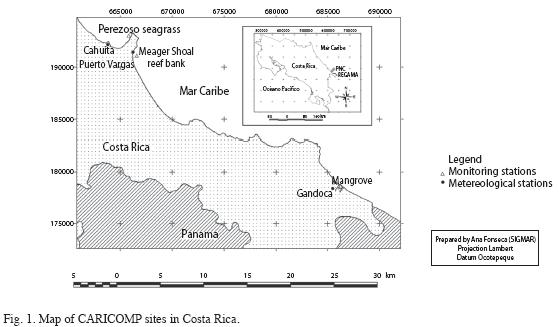
PNC is located on the southern Caribbean coast of Costa Rica (Fig. 1). It was created in 1970 and consists of 1 100ha of lowland rainforest and 22 400ha of arine territory, of which 600ha correspond to coral reef substrate. This is the largest and most studied fringing coral reef and associated seagrasses of Costa Rica (Cortés & Risk 1985, Cortés & Jiménez 2003, Fonseca 2003, Cortés 2009a). The coral reef at PNC consists of three reef crests; the main one (5 km long) along the front of Punta Cahuita and the other two on either sides of this point. A narrow spur-and-groove system reaches depths of 10-15 m on the fore-reef of the main crest. Several small patch reefs and seagrass beds occur in the lagoon and there are several offshore carbonate banks (Risk et al. 1980, Cortés & Risk 1985). This reef also has one of the highest diversities in the country, including: three species of stony hydroids (Cortés 1992), 31 species of zooxanthellate scleractinian corals (Cortés & Guzmán 1985b, Cortés 2009b), six species of azooxanthellate corals (Cortés 2009b) and 19 species of octocorals (Guzmán & Cortés 1985, Breedy 2009). There are also four species of seagrass (Wellington 1974, Cortés & Salas 2009).The coral reef at PNC is considered an economic asset to the local region estimated at over US $1.4 million a year (Blair et al. 1996). The population of Cahuita increased from 3 000 to 4 000 inhabitants in the last 13 years, and it receives more than 50 000 tourists per year (MINAE pers. comm.). The main economic activities in the town are tourism and fishing (Cortés in prep.). The coral reef monitoring site is the reef bank known as Meager Shoal (9º43´50" N 82º48´32" W), located 1km offshore from the beach of Puerto Vargas and south from the main reef crest of Cahuita (Fig. 1). It has a circular area of about 10 000m2, and the base of the bank is 10m deep while the top is at 7m. Corals were restricted to the top and the dominant species were Agaricia agaricites, Porites astreoides, Siderastrea siderea and Montastraea faveolata. Other coral species present were Mycetophyllia spp., Madracis decactis, Dichocoenia stokesii and Siderastrea radians. This reef bank was surrounded by terrestrial mud, which was easily re-suspended by the prevailing currents and strong waves. There are 10 permanent transects located at this site (Fonseca et al. 2006).
The seagrass monitoring site is known by local fishermen as "Perezoso" (9°44´13.3" N-82°48´24" W), located 500m north of the Perezoso (Sloth) creek, within the lagoon of the fringing reef of Cahuita (<1m deep). The is dominated by turtle grass, Thalassia testudinum, intermixed with manatee grass, Syringodium filiforme (Risk et al. 1980, Cortés & Guzmán 1985a, Paynter et al. 2001, Fonseca et al. 2007a, Nielsen-Muñoz & Cortés 2008). Flowering of T. testudinum occurred between March and June (Fonseca et al. 2007a, Nielsen- Muñoz & Cortés 2008). The permanent station for atmospheric temperature and rainfall measurements at PNC was located in the park rangers house at Puerto Vargas (9º43´56" N-82º48´58" W).
Mangrove of the Gandoca-Manzanillo National Wildlife Refuge (REGAMA)
Methods
We used the Caribbean Coastal Marine Productivity (CARICOMP Level I and II) methodology (http://isis.uwimona.edu.jm/centres/cms/caricomp/carinew.htm) to compare our monitoring results with those from other Caribbean countries. This protocol was established in the region since 1990. In Costa Rica, attempts to set up a permanent station at the coral reef of Cahuita were carried out in 1997, but without local community involvement the rods marking the sites were frequently lost (Cortés 1998). In 1999 new locations in the three ecosystems were marked and although data is only complete for some years, they have been evaluated yearly since. Concurrently, the local communities were educated about the importance of marine and coastal ecosystems and trained to collaborate with the scientists togather information, specially the weekly physical measurements.
Daily measurements
Continuously recording temperature sensors gathered air and water temperature information every 15 minutes; precipitation was measured in the meteorological stations by the park rangers and volunteers.
Weekly measurements
Weekly, volunteers from the local communities measured surface water emperature and salinity in Cahuita.
Coral reef
Meager Shoal has 10 permanent 10m long transects, and substrate relative cover is assessed yearly with the continuous intercept chain method, as is sea urchin density using the 1m wide belt method. Additionally, four transects (20x2m) were set to evaluate coral colony density and diseases. On September 13 2000, ten samples were taken to assess the algal biomass. From 1999 to 2005, two water samples (2 liters each), were taken during the rainy and dry seasons to determine the suspended particulate matter (SPM). Fish composition and density were evaluated from 2005 to 2007 following the AGRRA (Atlantic and Gulf Rapid Reef Assessment) protocol, i.e. 30x2 m belt transects (n=10), and in 2004, 2005 and 2007 using the REEF (Reef Environmental Education Foundation) rover diver method. Water temperature was recorded with an underwater temperature sensor, salinity with a refractometer and transparency with a Secchi disk in 1999, 2000 and 2001.Seagrasses
The first year, samples (n=6) were taken monthly, from March 1999 to April 2000, in order to determine seasonal productivity. Data from May and December, 1999 and February and March 2000 could not be collected because of water turbulence and low visibility. Based on the monthly sampling between 1999 and 2000, June showed the highest productivity and February the lowest. Productivity was supposed to be evaluated afterward in these two months yearly; however, in recent years, rough seas precluded data taking in July. Only one month was sampled in 2001, 2003, 2006 and 2007, two in 2004, three in 2000 and 2005 and four in 2008. At present, there is an underwater temperature sensor to monitor the water temperature around the seagrasses at 15 min intervals. At the same time, the local community was educated about the importance of seagrasses and scientific research and on gathering information, specially marking the shoots for growth measurements.
Mangrove
Three 10x10m plots were established in 1999 to evaluate biomass, productivity, seedlings and forest structure. They were located in an accessible area, close to the boundary between the mangrove and the lagoon´s water body. There was a permanent underwater sensor to monitor the water temperature within the mangrove roots (at a depth of 0.1m). Unfortunately, the mangrove plots were lost due to flooding in January 2005, so new ones have been established. The first year, samples were taken monthly, from June 1999 to March 2000, to determine the productivity peaks. During the rainy season, July showed the highest productivity. Therefore, productivity was evaluated again in July 2000, 2001, 2004, 2007 and 2008. At the same time, the local community was educated about the importance of mangroves and trained to help scientific studies, specially collecting litter fall from the productivity boxes.
Statistical analysis
Results
Atmospheric temperature and rainfall
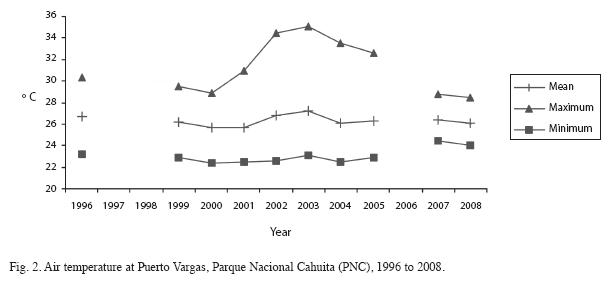
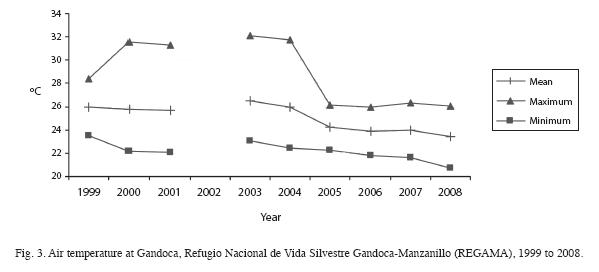
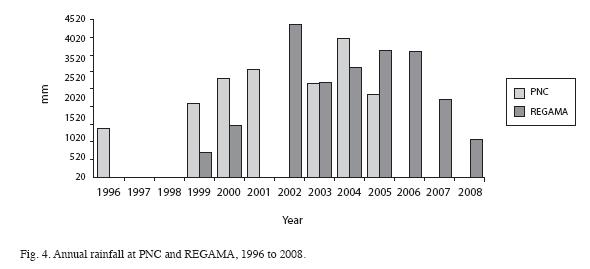
In the last 10 years, air temperature oscillated between 22.5 and 35.1°C, averaging around 26.5°C at PNC (Fig. 2) and between 20.7 and 32.1°C, with an average around 26°C at REGAMA (Fig. 3). Yearly rainfall ranged from approximately 1 400 to 4 000 mm at PNC and from approximately 1 400mm at REGAMA (Fig. 4). The warmest months were May to June and September to October at both locations.
Water temperature, salinity and suspended particulate matter
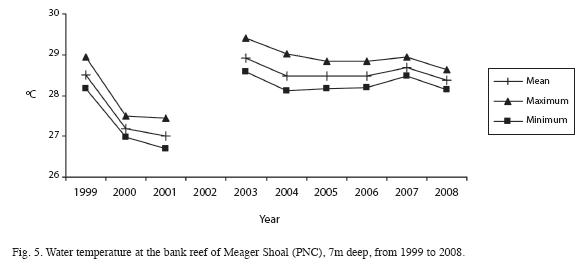
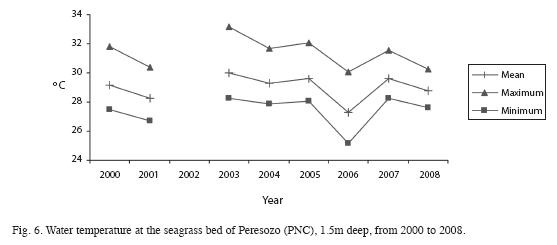
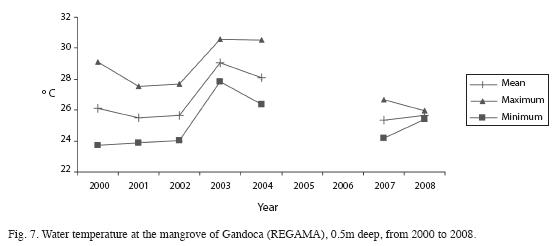
In PNC, water temperature at 7m depth ranged from 26.6 to 29.6°C (Fig. 5) and from 25.3 to 33.1°C at 1.5m depth (Fig. 6). The maximum water temperature was recorded in 2003 and the minimum in 2001. Salinity at the PNC coral reef ranged from 34.2±1.2 to 37.8±0.8 PSU (n=19) and the mean Secchi depth was 3.5±2.2 m (n=19). Suspended particulate matter ranged from 4.1 to 34.5mg/l, with an average of 17.5±2.4 mg/l.In REGAMA, water temperature within the mangrove roots ranged from 23.7 to 30.5°C, with spikes as low as 21.9°C and as high as 36.1°C. The highest water temperatures were in October 2004 and the lowest temperatures in February 2001 (Fig. 7). Salinity of interstitial waters ranged from 6 PSU (April) to 14 PSU (September), with an average of 8.2±4.1 between 1999 and 2009.
Overall, temperature seems to be decreasing, from 2003 to 2008 at the reef site (Fig. 5),but was higher than previous recorded years. The seagrass site (Fig. 6) also presents a decrease in temperature from a high in 2003 to levels similar to those recorded in previous years.
Coral reef
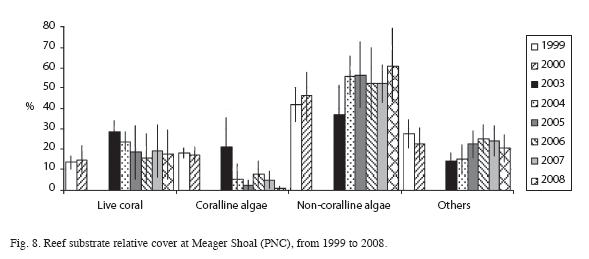
Live coral cover ranged from a low of 13.3% in 1999 to a high of 28.3% in 2003, dropping to a little less than 20% (with difference less than 3%) from 2005 to 2008. However, non-coralline algae increased 20% (F=6.9, df=1, p<0.05), and coralline algae decreased 11% (F=17.9, df=1, p<0.05). Total algae cover was higher than the live coral cover, and has an increasing trend, especially between 2000 and 2004 (F=7.2, df=1, p<0.05) (Fig. 8). Algal biomass was 1.92±1.29g/100m2 in 2000. Turf algae (<1cm high) was dominant and macroalgae biomass was low. The mean density of coral colonies has been decreasing, from as high as 4.6 colonies/ m2 in 1999 to 3.0 colonies/m2 in 2004 (using the AGRRA method). In 2008, using the ARICOMP method, there were 2.3±0.4 colonies/m2 in areas with dense cover and 0.5±0.3 colonies/ m2 in lower density areas. The dominant coral species were Agaricia agaricites, Porites astreoides, and Siderastrea siderea. The proportion of affected colonies by diseases, injuries and bleaching decreased from 24% in 2000 to 11% in 2004; however, but the difference is not statistically significant (F=0.7, df=1, p>0.05) (Fig. 9). In both years White Plague Disease (WPD) and Dark Spot Disease I (DSD-1) were the main diseases. Diseases affected mainly colonies of S. siderea, and bleaching was seen mainly in S. siderea and P. astreoides. Black Band Disease (BBD), Yellow Band Disease (YBD), Dark Spot Disease (DSD-II), White Band Disease (WBD) and Aspergillosis (ASP) were not observed. No diseased or bleached corals were encountered in 2008. Density of Diadema antillarum (between 2 and 7ind/m2) and Eucidaris tribuloides were low in Meager Shoal, though there was an increase in the first species. Density of Echinometra viridis was relatively high, yet decreased significantly from 20 to 0.6ind/m2 between 2000 to 2008 (F=10.8, df=1, p<0.05) (Fig. 10). Fish density ranged from 8.8 to 17.3ind/100m2, with the parrot fishes the most abundant group. The largest fish were encountered in 2006 (20cm angelfishes) and most fishes had a mean size of less than 15cm with the AGRRA protocol (Table 1). Using the REEF protocol, pomacentrids and labrids were the most abundant fishes in Meager Shoal (Table 2). A total of 51 fish species were ncountered after four years of fish counts (Table 2).
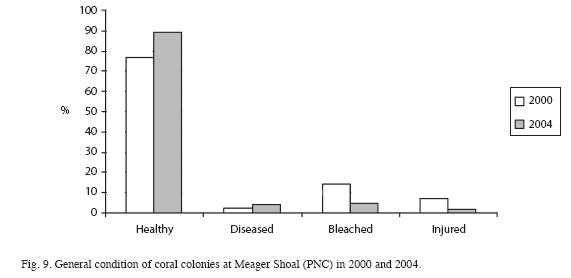
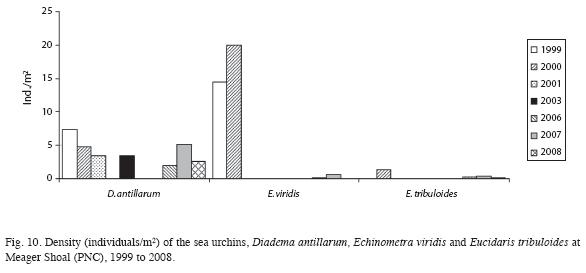
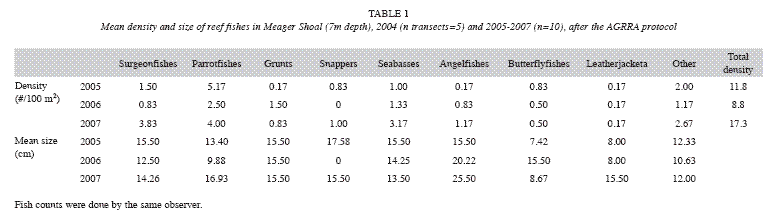
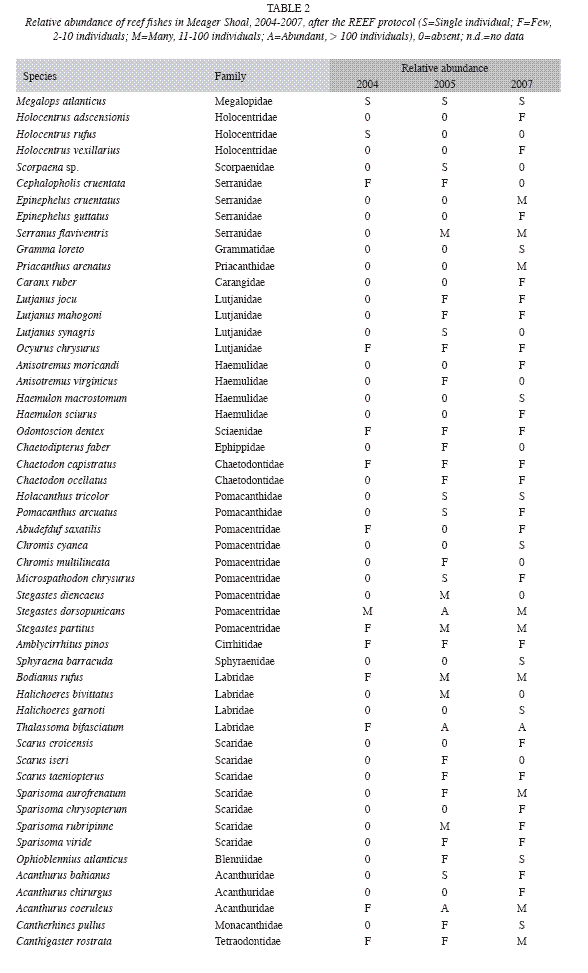
Seagrasses
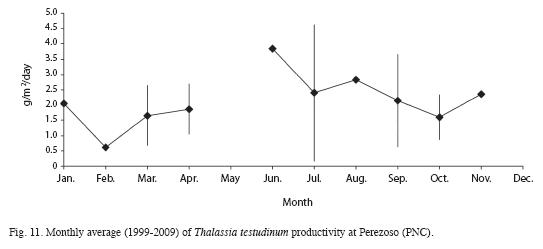
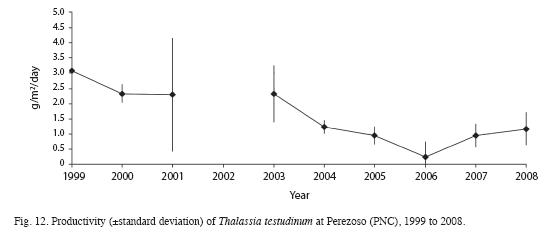
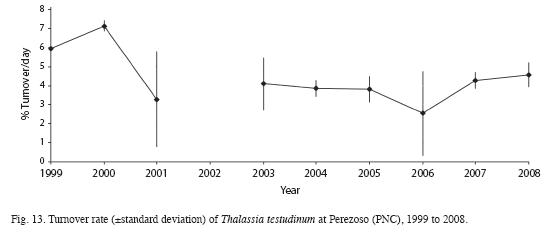
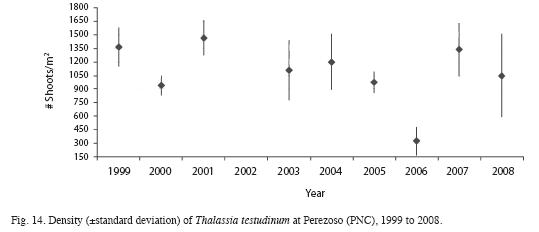
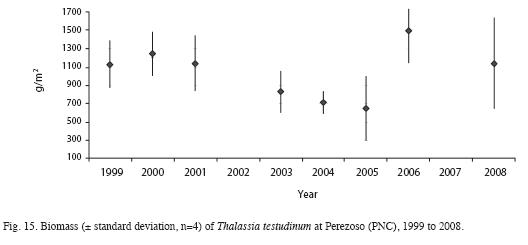
Mean length, width and area of Thalassia testudinum leaves and Leaf Area Index have decreased from the 2000 survey compared to 2008 (Table 3). Sexual reproduction was observed between March and June. Productivity was highest in June and lowest in February (Fig. 11). Between 1999 and 2008, productivity decreased from 3.1 to 0.24g/m2/day (Fig. 12). Turnover rate was also higher in 1999-2000 than the rest of the period, and has remained relatively stable since 2001 around 4% (Fig. 13). Shoot density has oscillated between 900 and 1 500shoots/m2, with an outlier of 300shoots/m2 in 2006 (Fig. 14). Density was considerably higher in April 1999, compared to September 1999 and March 2000 (F=3.65, df=8.95, p<0.05). There are highly significant differences between years in terms of productivity (ANOVA, F=13.13; df=4.47, p<0.000; Fig. 12). The Tukey a posteriori test indicates that differences are highly significant (p<0.001) during 1999-2003 and the remaining years. Turnover rate was also significantly different between years (ANOVA, F=8.327, p=0.000; Fig. 13), specifically during 1999-2000 and the remaining years (Tukey, p<0.01). Seagrass density was significantly lower in March 2000 when compared to March 1999 and March 2001 (F=4.25, df=4, p<0.05) (Fig. 14). Mean seagrasses biomass (T. testudinum) at Perezoso in March, from 1999 to 2008 has oscillated between a minimum of 650g/m2 in 2005 and a maximum of 1 500 g/m2 in 2006 (Fig. 15). Biomass decreased with time (F=3.84, df=4, p<0.05) and was significantly lower in 2005 than in 1999; however, it has gone up since then, to some of the highest levels so far. Seagrass biomass showed a high inverse correlation with mean (r=-0.87), minimum (-0.66) and especially with maximum temperature (-0.97). Productivity also showed a high inverse correlation with maximum temperature (-0.74).
Mangrove
The variables measured to determine forest structure showed no significant differences between years (F=0.32, df=4, 160, p>0.05). Mean circumference at breast height of R. mangle was 46.0±31.3cm, prop root length 2.2±1.2m, trunk length 2.5±2.8m, tree height 11.8±2.2m, trunk diameter 14.8±9.8cm, basal area 22.9±9.1m2/ ha, and trunk volume 0.3±0.2m3. With a mean density of 9.3±1.3trees/10 m2, mean biomass was 4.6±0.7kg/m2, after Golley et al. (1962), and 13.9±5.6kg/m2 using Cintron and Schaeffer- Novelli (1984) equation. Sapling mortality from 1999 to 2000 was 46%, and 66.4% during 2007-2008; mean sapling height was 45.2±18.8cm. Total productivity was significantly higher in July (H=76.43, df=9, p<0.05) and averaged 156.6±37.7g/m2/month. Flower production was also significantly higher in July (H=128.95, df=9, p<0.05), while fruit production was low and highly variable without significant difference between months (H=15.09, df=9, p>0.05). Leaves corresponded to 67% of total litter fall, flowers 13%, bracts 7%, fruits 7%, branches 6% and 0.21% is miscellaneous material.
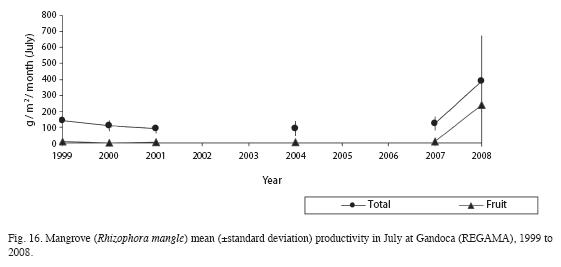
Yearly productivity was significantly higher in July 1999 than the same month in 2001 and 2004 (F=12.34, df=3, p<0.05, Fig. 16). However, when this period is compared to 2007-2008, productivity is currently significantly higher (F=13.26, df=3, p<0.001). The water table in the mangrove forest rises to 33 cm above the floor and the salinity is as low as 7PSU.
Discussion
Coral reef
Water temperatures were optimal for coral growth at the study site, which is close to the corals upper tolerance level. But, in 2003 and 2005 there were warming events that caused coral bleaching (Fonseca et al. 2006, McField et al. 2008, Fonseca & Nielsen-Muñoz in prep.). Although the proportion of diseased colonies decreased by 14%, live coral and coralline algae cover are low, while non-coralline and turf algae dominated the substrate and have increased. Since the decline in live coral cover, from 40% in the late 1970´s to 11% in the early 1990´s (Cortés 1994), in the last four years it has remained relatively stable at around 18%. Suspended particulate matter continues to be the main source of stress and have increased between 1979 (9mg/l) and 1992 (~20mg/l) (Cortés 1994), reaching 24mg/l in 2005 (Roder 2005, Roder et al. 2009). It seems that live coral cover cannot increase due to the presence of these sediments, possibly affecting coral recruitment as well as the diversity of reef-building coral species that can grow under those conditions. The sea urchin D. antillarum, a key herbivore species, was common in Cahuita (43 ind/10m2; Cortés 1981) before the 1983 mass mortality in the Caribbean region (Lessios et al. 1984; Murillo & Cortés 1984). Since then, the density of D. antillarum decreased (0.01ind/10m2, Cortés 1994) but seems to be recovering as densities are now higher than 10 years ago, with 0.1-1ind/10m2 in shallower sites of Cahuita (Fonseca 2003, Alvarado et al. 2004) and 2-7ind/10m2 from 2000 to 2008 in Meager Shoal. Additionally, several individuals were seen spawning in 2000 (Fonseca 2003). In the last eight years the densities of E. viridis decreased significantly (Fonseca et al. 2006 and Fig. 10). It has been observed at other coastal points that populations of E. viridis vary widely during the year apparently due to changes in algal density (J.J. Alvarado, pers. com. 2010).
Seagrasses
Seasonal differences in T. testudinum density, biomass and primary production, with maximum values in the summer and minimum values in winter, have been reported by Zieman (1974a, b, 1975), Thorhaug and Roesler (1977) and Van Tussenbroek (1995) for sites in the United States and Mexico. Leaf production reaches peak levels from May to July, in the Gulf of Mexico (Van Tussenbroek 1995), which coincides with the peak of productivity and turnover rate found in June in this study. Kaldy & Dunton (2000), found that seasonal fluctuations are the dominant controls on growth and production of T. testudinum in a subtropical coastal lagoon (Lower Laguna Madre, Texas). The more stable light and temperature conditions in tropical regions are reflected in a generally more uniform biomass throughout the year (Duarte 1989, Hillman et al. 1989). However, considerable seasonal biomass fluctuations have been observed in the tropics. Moderate changes in daily light period and temperature in some cases partially explain such observations (Mellors et al. 1993, Lanyon & Marsh 1995).
Total above and below ground biomass was highest (>4 000g/m2) in Belize, lowest in Tobago and Curacao (200-500g/m2), and intermediate to high (750-1 500g/m2) at all other CARICOMP sites (Colombia, Cuba, Mexico, Puerto Rico and Venezuela), including Costa Rica (978.6±281.6 g/m2, n=42). Intermediate biomass of Thalassia testudinum in Costa Rica could be attributed to greater abundance of epiphytes on their leaves as reported for Mochima Bay, Venezuela (Díaz-Díaz & Liñero-Arana 2007). Productivity in the Caribbean varies from <0.5g/m2/d (Bahamas) to >5g/m2/d (Venezuela) and turnover rates average 3.9%/d. In Costa Rica, productivity (1.6±0.7g/m2/d, n=74) is also intermediate, compared to other sites, and similar to that found in Colombia. Turnover rates are high (4.4±1.2%, n=74) compared to what was found in March and August at other sites. Similar values were found between March and June in Barbados, México, Bermudas and Puerto Rico (Linton & Fisher 2004). Turnover rates are probably higher in seagrass beds growing along the shorelines of continents or big islands subjected to a higher discharge of terrestrial nutrients, which makes them more vigorous. Shoot densities average 725shoots/m2 in the Caribbean region but in Costa Rica are higher (1079±250.0shoots/m2) perhaps due to the sampling sites being in a protected area. Average leaf length and width in the entire region were 14.4cm and 10.6mm, respectively, which are higher than in Costa Rica (9.2cm and 7.9mm respectively). The average leaf area index for the Caribbean sites is 3.4, while it is 2.4 in Costa Rica (CARICOMP 1997b, Creed et al. 2003, Linton & Fisher 2004).
Since the seagrass beds in Cahuita are found adjacent to the fringing coral reef, they are in close interaction and serve as nursery and feeding ground for reef organisms. Recreational boating, swimmers and nutrient loading from deforested lands in the coast, the upstream rivers and local pollution (Roder 2005, Roder et al. 2009) are all potential sources of impact to seagrass cover and productivity and therefore to the fishing activity of the coastal community (Creed et al. 2003). These interaction need to be evaluated in the near future. Furthermore, establishing the habitat requirements of each seagrass species and evaluating their particular response to water quality and sediment types is also needed.
Mangrove
The peak of productivity and flower production of R. mangle in Costa Rica were in July. Mangrove productivity decreased since 2001, and water temperature seems to have decreased; however, more sampling is needed to determine if they are correlated. The productivity trend for CARICOMP sites was clearly related to temperature, and also appears to be affected by rainfall, yet a more extensive data set will be necessary to evaluate this relationship.
Conclusions
-Coral reefs at Cahuita National Park continue to be impacted by terrigenous sediments.
-Compared to other CARICOMP sites, live coral cover, fish diversity and density, and sea urchin density were low, while algal cover was intermediate. Seagrass productivity, biomass, average leaf length and width were intermediate, and seagrass turnover rates, shoot density nd leaf area index were rather high. Mangrove biomass and density were relatively low, while productivity was intermediate.
-Management actions, mainly inland, should be considered in order to protect and preserve these important coastal ecosystems. The recovery of forest cover, especially riverine forests, in the watersheds must be a priority to reduce sediments loads that reach the coral reefs.
Acknowledgments
Monitoring the coastal ecosystems of our Caribbean coast have been possible thanks to the support of CARICOMP, Vicerrectoría deInvestigación and CIMAR (Universidad de Costa Rica), Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICIT), and the United Nations Environmental Program- Jamaica (UNEP). Thanks also to the local communities of Cahuita and Gandoca, all the personnel at Cahuita National Park and the REGAMA and the assistants and administrators from CIMAR for their help. We appreciate the review of the paper by Christopher S. Vaughan, and the detailed comments and suggestion of two anonymous reviewers.
Literature cited
Alfaro, E.J. 2002. Some characteristics of the annual precipitation cycle in Central America and their relationships with its surrounding tropical oceans. Tóp. Meteorol. Oceanogr. 9: 88-103. [ Links ]
Blair, N., C. Geraghty, G. Gund & B. Jones. 1996. An economic evaluation of Cahuita National Park: establishing the economic value of an environmental asset. Kellogg Graduate School of Management, Boston. Tech Rept. 24 p. [ Links ]
Breedy, O. 2009. Octocorals. Text: Pp. 161-167, Species List: Compact Disc Pp. 108-111. In: I.S. Wehrtmann and J. Cortés (Eds.). Marine Biodiversity of Costa Rica, Central America. Monogr. Biol. 86. Springer + Business Media B.V., Berlin, Germany. [ Links ]
CARICOMP. 1997a. Caribbean coastal marine productivity (CARICOMP): a research and monitoring network of marine laboratories, parks, and reserves. Proc. 8th Int. Coral Reef Symp., Panamá 1: 641-646. [ Links ]
CARICOMP 1997b. Variation in ecological parameters of Thalassia testudinum across the CARICOMP Network. Proc. 8th Int. Coral Reef Symp., Panamá 1: 663-668. [ Links ]
CARICOMP. 1997c. Structure and productivity of mangrove forests in the Greater Caribbean Region. CARICOMP. Proc. of the 8th ICRS, Panamá 1: 669-672. [ Links ]
CARICOMP. 2002. Status and temporal trends at CARICOMP coral reef sites. Proc. 9th Int. Coral Reef Symp., Bali, Indonesia 1: 325-330. [ Links ]
Cintrón, G. & Y. Schaeffer-Novelli. 1984. Methods for studying mangrove structure. Pp. 91-113. In S.C. Snedaker and J.G. Snedaker (eds.). The Mangrove Ecosystem: Research Methods. UNESCO, Bungay, United Kingdom. [ Links ]
Clough, B.F. 1992. Primary productivity and growth of mangrove forests. Pp. 225-249. In A.I. Robertson and D.M. Alongi (Eds.). Tropical Mangrove Ecosystems. Amer. Geophys. Union, Washington, D.C., USA. [ Links ]
Coll, M., A.C. Fonseca & J. Cortés. 2001. El manglar y otras asociaciones vegetales de la laguna Gandoca, Limón, Costa Rica. Rev. Biol. Trop. 49: 321-329. [ Links ]
Coll, M., J. Cortés & D. Sauma. 2004. Características físico-químicas y determinación de plaguicidas en el agua de la laguna de Gandoca, Limón, Costa Rica. Rev. Biol. Trop. 52: 33-42. [ Links ]
Cortés, J. 1981. The Coral Reef at Cahuita, Costa Rica: A Reef Under Stress. M.Sc. thesis, McMaster University, Hamilton, Ontario, Canada. [ Links ]
Cortés, J. 1991. Ambientes y organismos marinos del Refugio Nacional de Vida Silvestre Gandoca Manzanillo, Limón, Costa Rica. Geoistmo 5: 62-68. [ Links ]
Cortés, J. 1992. Organismos de los arrecifes coralinos de Costa Rica: V. Descripción y distribución geográfica de hidrocorales (Cnidaria; Hydrozoa: Milleporina and Stylasterina) de la costa Caribe. Brenesia 38: 45-50. [ Links ]
Cortés, J. 1994. A reef under siltation stress: a decade of degradation. Pp. 240-246. In R.N. Ginsburg (Compiler). Proc. Colloquium and Forum on Global Aspects of Coral Reefs: Health, Hazards and History. RSMAS, University of Miami, Miami, Florida. [ Links ]
Cortés, J. 1998. Cahuita and Laguna Gandoca, CostaRica. Pp. 107-113. In B. Kjerfve (ed.). CARICOMP: Caribbean Coral Reef, Seagrass and Mangrove Sites.UNESCO, Paris. [ Links ]
Cortés, J. 2009a. A history of marine biodiversity scientific research in Costa Rica, p. 47-80. In I.S. Wehrtmann & J. Cortés (eds.). Marine Biodiversity of Costa Rica,Central America. Monogr. Biol. 86. Springer + Business Media B.V., Berlin, Germany. [ Links ]
Cortés, J. 2009b. Stony corals, p. 169-173. Species List: Compact Disc p. 112-118. In I.S. Wehrtmann and J. Cortés (eds.). Marine Biodiversity of Costa Rica,Central America. Monogr. Biol. 86. Springer + Business Media B.V., Berlin, Germany. [ Links ]
Cortés, J. & H.M. Guzmán. 1985a. Arrecifes coralinos de la costa Atlántica de Costa Rica. Brenesia 23: 275-292. [ Links ]
Cortés, J. & H.M. Guzmán. 1985b. Organismos de los arrecifes coralinos de Costa Rica: III. Descripción y distribución geográfica de corales escleractinios (Cnidaria; Anthozoa: Scleractinia) de la costa Caribe. Brenesia 24: 63-124. [ Links ]
Cortés, J. & C.E. Jiménez. 2003. Past, present and future of the coral reefs of the Caribbean coast of Costa Rica, p. 223-239. In J. Cortés (ed.). Latin American Coral Reefs. Elsevier Science B.V., Amsterdam, Holland. [ Links ]
Cortés, J. & M.J. Risk. 1985. A reef under siltation stress: Cahuita, Costa Rica. Bull. Mar. Sci. 36: 339-356. [ Links ]
Cortés, J. & E. Salas. 2009. Seagrasses, p. 119-122. Species List: Compact Disc p. 71-72. In I.S. Wehrtmann & J. Cortés (eds.). Marine Biodiversity of Costa Rica, Central America. Monogr. Biol. 86. Springer + Business Media B.V., Berlin, Germany. [ Links ]
Cortés, J., R. Soto, C. Jiménez & A. Astorga. 1992. Death of intertidal and coral reef organisms as a result of a 7.5 earthquake. Proc. 7th Int. Coral Reef Symp., Guam 1: 235-240. [ Links ]
Cortés, J., R. Soto & C. Jiménez. 1994. Efectos ecológicos del terremoto de Limón. Rev. Geol. Amér. Central, Vol. Esp. Terremoto de Limón: 187-192. [ Links ]
Creed, J.C., R.C. Phillips & B.I. Van Tussenbroek. 2003. The seagrasses of the Caribbean, p. 234-242. In E.P. Green & F.T. Short. World Atlas of Seagrasses: Present Status and Future Conservation. University of California, Berkeley, USA. [ Links ]
Denyer, P. 1998. Historic-prehistoric earthquakes, seismic hazards and Tertiary and Quaternary geology of the Gandoca-Manzanillo National Wildlife Refuge, Limón, Costa Rica. Rev. Biol. Trop. 46: 237-250. [ Links ]
Díaz-Díaz, O. & I. Liñero-Arana. 2007. Biomass and density of Thalassia testudinum beds in Mochima Bay, Venezuela. Acta Bot. Venez. 30: 217-226. [ Links ]
Duarte, C.M. 1989. Temporal biomass variability and production/biomass relationships of seagrass communities. Mar. Ecol. Prog. Ser: 269-76. [ Links ]
Fonseca, A.C. 2003. A rapid assessment at Cahuita National Park, Costa Rica, 1999 (Part I: stony corals and algae). In J.C. Lang (Ed.). Status of Coral Reefs in the Western Atlantic: Results of Initial Surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA) Program. Atoll Res. Bull. 496: 249-257. [ Links ]
Fonseca, A.C. & J. Cortés. 2002. Land use in the La Estrella River basin and soil erosion effects on the Cahuita reef system, Costa Rica: 68-82 and 114-118. In: B. Kjerfve, W.J. Wiebe, H.H. Kremer, W. Salomons and J.I. Marshall Crossland (Eds.). Caribbean Basins: LOICZ Global Change Assessment and Synthesis of River Catchment/ Island-Coastal Sea Interaction and Human Dimension. LOICZ Reports and Studies No. 27. LOICZ, Texel, The Netherlands. [ Links ]
Fonseca, A.C. & C. Gamboa 2003. A rapid assessment at Cahuita National Park, Costa Rica, 1999 (Part 2: Reef fishes). In J.C. Lang (Ed.). Status of Coral Reefs in the Western Atlantic: Results of Initial Surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA) Program. Atoll Res. Bull. 496: 258-266. [ Links ]
Fonseca, A.C., E. Salas & J. Cortés. 2006. Monitoreo Del arrecife coralino Meager Shoal, Parque Nacional Cahuita (sitio CARICOMP). Rev. Biol. Trop. 54: 755-763. [ Links ]
Fonseca, A.C., V. Nielsen & J. Cortés. 2007a. Monitoreo de pastos marinos en Perezoso, sitio CARICOMP en Cahuita, Costa Rica. Rev. Biol. Trop. 55: 55-66. [ Links ]
Fonseca, A.C., J. Cortés & P. Zamora. 2007b. Monitoreo del manglar de Gandoca, Costa Rica (Sitio CARICOMP). Rev. Biol. Trop. 55: 23-31. [ Links ]
Ginsburg, R.N. & J. Lang. 2003. Foreword. In J.C. Lang (Ed.). Status of Coral Reefs in the Western Atlantic: Results of Initial Surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA) Program. Atoll Res. Bull. 496: vii-xiii. [ Links ]
Golley, F., H.T. Odum & R.F. Wilson. 1962. The structure and metabolism of a Puerto Rican red mangrove forest in May. Ecology 43: 9-19. [ Links ]
Guzmán, H.M. & J. Cortés. 1985. Organismos de los arrecifes coralinos de Costa Rica: IV. Descripción y distribución geográfica de octocoralarios de la costa Caribe. Brenesia 24: 125-174. [ Links ]
Herrera, W. 1984. Clima de Costa Rica. Editorial Universidad Estatal a Distancia (EUNED), San José, Costa Rica, 118 p. [ Links ]
Hillman, K., D.I. Walker, A.W.D Larkum & A.J. McComb.1989. Productivity and nutrient limitation, p. 635-685. In A.W.D. Larkum, A.J. McComb & S.A. Shepherd (eds.). Biology of Seagrasses. Elsevier, Amsterdam, Holland. [ Links ]
Hoegh-Guldberg, O., P.J. Mumby, A.J. Hooten, R.S. Steneck, P. Greenfield, E. Gomez, C.D. Harvell, P.F. Sale, A.J. Edwards, K. Caldeira, N. Knowlton, C.M. Eakin, R. Iglesias-Prieto, N. Muthinga, R.H. Bradbury, A. Dubi & M.E. Hatziolos. 2007. Coral reefs under rapid climate change and ocean acidification. Science 318: 1737-1742. [ Links ]
Hughes, T.P., A.H. Baird, D.R. Bellwood, M. Card, S.R. Connolly, C. Folke, R. Grosberg, O. Hoegh-Guldberg, J.B.C. Jackson, J. Kleypas, J.M. Lough, P. Marshall, M. Nyström, S.R. Palumbi, J.M. Pandolfi, B. Rosen & J. Roughgarden. 2003. Climate change, human impacts, and the resilience of coral reefs. Science 301: 929-933. [ Links ]
Jiménez, J.A. 1988. The dynamics of Rhizophora racemosa Meyer, forests on the Pacific coast of Costa Rica. Brenesia 30: 1-12. [ Links ]
Jiménez, J.A. 1994. Los Manglares del Pacífico Centroamericano. EFUNA, Heredia, Costa Rica. 352 p. [ Links ]
Jiménez, J.A. & R. Soto. 1985. Patrones regionales en la estructura y composición florística de los manglares de la costa Pacífica de Costa Rica. Rev. Biol. Trop. 33: 25-37. [ Links ]
Kaldy, J.E. & K.H. Dunton. 2000. Above-and belowground production, biomass and reproductive ecology of Thalassia testudinum (turtle grass) in a subtropical coastal lagoon. Mar. Ecol. Prog. Ser. 193: 271-283. [ Links ]
Lanyon, J.M. & H. Marsh. 1995. Temporal changes in the abundance of some tropical intertidal sea grasses in North Queensland. Aquat. Bot. 217-37. [ Links ]
Lee, K.S. & K.H. Dunton. 1997. Effects of in situ light reduction on the maintenance, growth and partitioning of carbon resources in Thalassia testudinum Banks ex König. J. Exp. Mar. Biol. Ecol. 210: 53-73. [ Links ]
Lee, K.S., S.R. Park & Y.K. Kim. 2007. Effects of irradiance, temperature, and nutrients on growth dynamics of seagrasses: A review. J. Exp. Mar. Biol. Ecol. 350: 144-175. [ Links ]
Lessios, H.A., D.R. Robertson & J.D. Cubit. 1984. Spread of Diadema mass mortality through the Caribbean. Science 226: 335-337. [ Links ]
Linton, D. & T. Fisher. 2004. Caribbean Coastal Marine Productivity Program, 1993-2004. CARICOMP, Kingston, Jamaica. [ Links ]
Lizano, O.G. 2006. Algunas características de las mareas en las costa Pacífica y Caribe de Centroamérica. Cienc. Tecnol. 24: 51-64. [ Links ]
Lizano, O.G. 2007. Climatología del viento y oleaje frente a las costas de Costa Rica. Cienc. Tecnol. 25: 43-56. [ Links ]
McField, M., N. Bood, A. Fonseca, A. Arrivillaga, A. Franquesa- Rinos & R.M. Loreto-Viruel. 2008. Status of the Mesoamerican reef after the 2005 coral bleaching event: 45-60. In: C. Wilkinson & D. Souter (eds.). Status of Caribbean Coral Reefs after Bleaching and Hurricanes in 2005. GCRMN & RRRC, Townsville, Australia. [ Links ]
Mellors, J.E., H. Marsh & R.G. Coles. 1993. Intra-annual changes in seagrass standing crop, Green Island, Northern Queensland. Austr. J. Mar. Freshwater Res. 44: 33-41. [ Links ]
Murillo, M.M. & J. Cortés. 1984. Alta mortalidad en la población del erizo de mar Diadema antillarum Philippi (Echinodermata: Echinoidea), en el Parque Nacional Cahuita, Limón, Costa Rica. Rev. Biol. Trop. 32: 167-169. [ Links ]
Nielsen-Muñoz, V. 2007. Abundancia, biomasa y floración de Thalassia testudinum (Hydrocharitaceae) en el Caribe de Costa Rica. Thesis, Universidad de Costa Rica, San Pedro, San José, Costa Rica. [ Links ]
Nielsen-Muñoz, V. & J. Cortés. 2008. Abundancia, biomasa y floración de Thalassia testudinum (Hydrocharitaceae en el Caribe de Costa Rica. Rev. Biol. Trop. 56: 175-189. [ Links ]
Paddack, M.J., J.D. Reynolds, C. Aguilar, R.S. Appeldoorn, J. Beets, E.W. Burkett, P.M. Chittaro, K. Clarke, R. Esteves, A.C. Fonseca, G.E. Forrester, A.M. Friedlander, J. García-Sais, G. González-Sansoón, L.K.B. Jordan, D.B. McClellan, M.W. Miller, P.P. Molloy, P.J. Mumby, I. Nagelkerken, M. Nemeth, R. Navas- Camacho, J. Pitt, N.V.C. Polunin, M.C. Reyes-Nivia, D.R. Robertson, A. Rodríguez-Ramírez, E. Salas, S.R. Smith, R.E. Spieler, M.A. Steele, I.D. Williams, C.L. Wormald, A.R. Watkinson & I.M. Côté. 2009. Recent region-wide declines in Caribbean reef fish abundance. Curr. Biol. 19: 1-6. [ Links ]
Pandolfi, J.M., R.H. Bradbury, E. Sala, T.P. Hughes, K.A. Bjorndal, R.G. Cooke, D. McArdle, L. McClenachan, M.J.H. Newman, G. Paredes, R.R. Warner & J.B.C. Jackson. 2003. Global trajectories of the long-term decline of coral reef ecosystems. Science 301: 955-958. [ Links ]
Paynter, C., J. Cortés & M. Engels. 2001. Biomass, productivity and density of the sea grass Thalassia testudinum at three sites in Cahuita National Park, Costa Rica. Rev. Biol. Trop. 49: 265-272. [ Links ]
Phillips, P.C. & M.J. Pérez-Cruet. 1984. A comparative survey of reef fishes in Caribbean and Pacific of Costa Rica. Rev. Biol. Trop. 32: 95-102. [ Links ]
Pool, D.J., S.C. Snedaker & A.E. Lugo. 1977. Structure of mangrove forests in Florida, Puerto Rico, Mexico, and Costa Rica. Biotropica 9: 195-212. [ Links ]
Risk, M.J., M.M. Murillo & J. Cortés. 1980. Observaciones biológicas preliminares sobre el arrecife coralino en el Parque Nacional Cahuita, Costa Rica. Rev. Biol. Trop. 28: 361-382. [ Links ]
Roder, C. 2005. Land-based pollution on the Caribbean coast of Costa Rica: nutritional and skeletal characteristics of the reef-building coral Siderastrea sidereal as a response. M.Sc. Thesis, ISATEC, University of Bremen. Bremen, Germany. [ Links ]
Roder, C., J. Cortés, C. Jiménez & R. Lara. 2009. Riverine input of particulate material and inorganic nutrients to a coastal reef ecosystem at the Caribbean coast of Costa Rica. Mar. Poll. Bull. 58: 1937-1943. [ Links ]
Short, F.T. & S. Wyllie-Echeverria. 1996. Natural and human-induced disturbances of sea grass. Environ. Conserv. 23: 17-27. [ Links ]
Short, F.T., R.G. Coles & C. Pergent-Martini. 2001. Global seagrass distribution, p. 5-30. In F.T. Short & R.G. Coles (eds.). Global Seagrass Research Methods. Elsevier Sci. Publ., Amsterdam, Holland. [ Links ]
Thorhaug, A. & M.A. Roesler. 1977. Seagrass community dynamics in a subtropical estuarine lagoon. Aquaculture 12: 253-277. [ Links ]
Underwood, A. 1997. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance. University of Cambridge, Cambridge, England. [ Links ]
Valiela, I., J.L. Bowen & J.K. York. 2001. Mangrove forests: one of the world´s threatened major tropical environments. BioScience 51: 807-815. [ Links ]
Van Tussenbroek, B.I. 1995. Thalassia testudinum leaf dynamics in a Mexican Caribbean coral reef lagoon. Mar. Biol. 122: 33-40. [ Links ]
von Ende, C.N. 1993. Repeated-measures analysis: growth and other time-dependent measures, p. 134-157. In S. Scheiner & J. Gurevich (eds.). Design and Analysis of Ecological Experiments. Oxford University, Oxford, England. [ Links ]
Waycott, M., C.M. Duarte, T.J.B. Carruthers, R.J. Orth, W.C. Dennison, S. Olyarnik, A. Calladine, J.W. Fourqurean, K.L. Heck, Jr., A.R. Hughes, G.A. Kendrick, W.J. Kenworthy, F.T. Short & S.L Williams. 2009. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Nat. Acad. Sci. (www.pnas.org_cgi_doi_10.1073_pnas.0905620106). [ Links ]
Wellington, G.M. 1974. The benthic flora of Punta Cahuita: annotated list of species with additions to the Costa Rican Atlantic flora. Brenesia 3: 19-30. [ Links ]
Zieman, J.C. 1974a. Methods for the study of the growth and production of turtle grass, Thalassia testudinum König. Aquaculture 4: 139-143. [ Links ]
Zieman, J.C. 1975. Seasonal variation of turtle grass, Thalassia testudinum Konig, with reference to temperature and salinity effects. Aquat. Bot. 1: 107-123. [ Links ]
Zieman, J.C. & E.J.F. Wood. 1975. Effects of thermal pollution of tropical-type estuaries, with emphasis on Biscayne Bay, Florida, p. 75-98. In E.J.F. Wood & R. Johannes (eds.). Tropical Marine Pollution. Elsevier, Amsterdam, Holland. [ Links ]
Ana C. Fonseca. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280 / World Wildlife Fund (WWF), San José, Costa Rica, Interlink #718, P.O. Box 02-5635, Miami, Florida 33102, USA.
Jaime Nivia-Ruiz. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280
Vanessa Nielsen-Muñoz. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280
Jimena Samper-Villarreal Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280
Eva Salas. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280Solciré Martínez. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280
Priscilla Zamora-Trejos. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica. Fax (506) 2511-3280
Received 03-VIII -2009. Corrected 17-II -2010. Accepted 12-VII -2010.

