










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.58 suppl.1 San José May. 2010
Mid-term coral-algal dynamics and conservation status of a Gorgona Island (Tropical Eastern Pacific) coral reef
Fernando A. Zapata1, Alberto Rodríguez-Ramírez2, Carlos Caro-Zambrano2 & Jaime Garzón-Ferreira2
1. Departamento de Biología, Universidad del Valle, Apartado Aéreo 25360, Cali, Colombia; fernando.zapata@correounivalle.edu.co
2. Instituto de Investigaciones Marinas y Costeras (INVEMAR), Apartado Aéreo 1016, Santa Marta, Colombia; betorod@invemar.org.co, cabetoz78@hotmail.com, jgarzon@invemar.org.co
Abstract
Colombian coral reefs, as other reefs worldwide, have deteriorated significantly during the last few decades due to both natural and anthropogenic disturbances. The National Monitoring System for Coral Reefs in Colombia (SIMAC) was established in 1998 to provide long-term data bases to assess the changes of Colombian coral reefs against perturbations and to identify the factors responsible for their decline or recovery. On the Pacific coast, data on coral and algal cover have been collected yearly during seven consecutive years (1998-2004) from 20 permanent transects in two sites at La Azufrada reef, Gorgona Island. Overall, coral cover was high (55.1%-65.7%) and algal cover low (28.8%-37.5%) and both exhibited significant changes among years, most notably on shallow areas. Differences between sites in both coral and algal cover were present since the study began and may be explained by differences in sedimentation stress derived from soil runoff. Differences between depths most likely stem from the effects of low tidal sub-aerial exposures. Particularly intense sub-aerial exposures occurred repeatedly during January-March, 2001 and accounted for a decrease in coral and an increase in algal cover on shallow depths observed later that year. Additionally, the shallow area on the Northern site seems to be negatively affected by the combined effect of sedimentation and low tidal exposure. However, a decrease in coral cover and an increase of algal cover since 2001 on deep areas at both sites remain unexplained. Comparisons with previous studies suggest that the reef at La Azufrada has been more resilient than other reefs in the Tropical Eastern Pacific (TEP), recovering pre-disturbance (1979) levels of coral cover within a 10 year period after the 1982-83 El Niño, which caused 85% mortality. Furthermore, the effects of the 1997-98 El Niño, indicated by the difference in overall live coral cover between 1998 and 1999, were minor (<6% reduction). Despite recurrent natural disturbances, live coral cover in 2004 was as high as that existing before 1982 at La Azufrada, and one of the highest observed on healthy coral reefs in the TEP region. Rev. Biol. Trop. 58 (Suppl. 1): 81-94. Epub 2010 May 01.
Key words: algae, corals, coral reefs, Gorgona Island, spatial variation, temporal dynamics, Tropical Eastern Pacific.
Resumen
A través del Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia (SIMAC) se han colectado anualmente (1998-2004) datos de cobertura de corales y algas en 20 transectos de 10m localizados en dos profundidades de dos sitios del arrecife de La Azufrada, Isla Gorgona (costa Pacífica). En general, la cobertura coralina fue alta (55.1% - 65.7%) y la de algas baja (28.8% - 37.5%), con una significativa variación entre años, más notablemente en áreas someras. Las diferencias entre sitios tanto en cobertura de coral como de algas estuvieron presentes desde que comenzó el estudio y pueden ser explicadas por diferencias en estrés por sedimentación (escorrentía terrestre). Las diferencias entre las profundidades muy probablemente se deben a los efectos de exposiciones aéreas durante mareas bajas. Las exposiciones aéreas particularmente intensas y repetidas entre enero-marzo de 2001 explican la disminución de la cobertura de coral, y aumento de la de algas en áreas someras observadas más tarde ese año. Adicionalmente, el área somera del sitio norte parece ser negativamente afectada por el efecto combinado de la sedimentación y la exposición mareal. Sin embargo, una tendencia a la disminución de la cobertura coralina y aumento de la de algas desde 2001 en las áreas profundas de ambos sitios permanece sin explicación. Comparaciones con estudios previos indican que el arrecife de La Azufrada ha sido más elástico que otros arrecifes del Pacífico Oriental Tropical (POT). A pesar de las perturbaciones naturales recurrentes, la cobertura coralina en 2004 fue tan alta como la existente antes de 1982 en La Azufrada, y una de las más altas observadas en arrecifes coralinos saludables de la región del POT.
Palabras clave: algas, corales, arrecifes coralinos, dinámica temporal, Isla Gorgona, Pacífico Oriental Tropical, variación espacial.
Our perception of coral reefs has changed from predictable and stable ecosystems developed under benign environmental conditions to highly dynamic ones in which dramatic changes often occur as part of natural cycles of disturbance and recovery over various spatial and temporal scales (Connell 1978, 1997, Aronson & Precht 1997, Brown 1997a, Connell et al. 1997, 2004, Pandolfi 1999). Nonetheless, signs of deterioration of coral reefs worldwide have increased at alarming rates in the last few decades (Bryant et al. 1998) and there is increasing evidence that human activity may be upsetting the dynamics of natural cycles to a point beyond which coral reefs may not recover (Nyström et al. 2000, Hughes et al. 2003, Pandolfi et al. 2003). Signs of deterioration include a progressive reduction in live coral cover and a shift to dominance by benthic algae (Hallock et al. 1993, Ginsburg 1994, Hughes 1994, Gardner et al. 2003), massive mortalities of functionally important organisms (Gladfelter 1982, Lessios et al. 1984, Carpenter 1990, Garzón-Ferreira & Zea 1992, Liddell & Ohlhorst 1992, Hughes 1994), depletion of fisheries resources (Rogers 1985, Hughes 1994) and an increase in the frequency, intensity and geographic distribution of coral bleaching events (Williams & Bunkley-Williams 1990, Glynn 1993, Brown 1997b, Glynn et al. 2001, McWilliams et al. 2005). In general, the causes of these changes are associated with increased human settlement adjacent to coral reefs (Hallock et al. 1993, Wilkinson 1992, Birkeland 1997), El Niño events and global warming (Enfield 2001, Aronson et al. 2002, Harvell et al. 2002, Hughes et al. 2003, Buddemeier et al. 2004), continental runoff and sewage discharge that increase turbidity, sedimentation and nutrient loads in coastal zones (Loya 1976, Wittenberg & Hunte 1992, Hallock et al. 1993, Szmant 2002), indiscriminate fishing of commercially valuable organisms (Munro 1983, Jennings & Polunin 1996, Jackson et al. 2001, Pandolfi et al. 2003), recreational activities that damage or disturb coral reef organisms (Díaz et al. 2000, Rouphael & Inglis 2002), and the proliferation of coral diseases (Santavy & Peters 1997, Garzón-Ferreira et al. 2001, Sutherland et al. 2004).
Clearly, a combination of both natural and human-induced factors has led coral reefs worldwide to deteriorate significantly during the last decades, and ecologists now face the challenge of assessing the extent to which changes occurring on coral reefs are the result of natural vs. anthropogenic processes and their interactions. However, few coral reef sites have been studied for sufficiently long periods (e.g., Connell et al. 1997, 2004) to document their background variability, measure significant departures and partition causal processes of change into natural and anthropogenic components. Colombian coral reefs have not been exempt from the processes of degradation occurring on coral reefs in general and have shown signs of deterioration in most areas (Díaz et al. 2000). In the Colombian Caribbean live coral cover decreased by an average of ~38% during the last 30 years of the 20th century reaching an average of 20-30% in some areas (Garzón-Ferreira & Kielman 1994, Garzón-Ferreira 1997, Díaz et al. 2000, Garzón-Ferreira et al. 2002a). In contrast, the decline of Colombian Pacific coral reefs has been much less severe. Current levels of live coral cover are relatively high (≥50-74% average; Rodríguez-Ramírez et al. 2006) in spite of the massive coral mortality that occurred throughout the Tropical Eastern Pacific (TEP) during the 1982-83 El Niño event (Glynn 1990, Vargas-Ángel et al. 2001, Zapata et al. 2001, Zapata & Vargas-Ángel 2003). However, detailed quantitative assessments of the impacts of natural and anthropogenic perturbations on most Colombian coral reefs have been hampered by the lack of adequate baseline data on the natural variability of their major physical and biological components. Although a number of studies have described many of Colombian coral reefs and documented their most significant changes (Garzón-Ferreira & Kielman 1994, Díaz et al. 1995, 1996, 2000, Zea et al. 1998, Zapata et al. 2001, Zapata & Vargas-Ángel 2003); only in 1998 a monitoring program was established to evaluate the status and dynamics of such reefs. The National Monitoring System for the Coral Reefs of Colombia (SIMAC for its Spanish acronym) has obtained detailed data for several coral reefs including both Caribbean and Pacific areas. Here we report the results of analyses of some of these data (1998-2004) for La Azufrada reef, located at Gorgona Island and one of the largest and best developed coral reefs in the Colombian Pacific (Zapata & Vargas-Ángel 2003). We focused on temporal and spatial variation in abundance of corals and algae because they are major benthic components of coral reef communities and serve as primary indicators of coral reef "health" (Brown 1997a).
Materials and methods
Study area: Coral reefs in the Colombian Pacific occur only at a few localities, covering 0.5km2 (Díaz et al. 2000, Zapata & Vargas-Ángel 2003). The largest and most developed coral reefs occur at Gorgona Island (2°58'N, 78°10'W), which is a continental island located 35km off the Colombian continental coast (Fig. 1), on the Southern end of Panama Bight (Glynn et al. 1982). The climate at Gorgona is primarily influenced by the position of the Intertropical Convergence Zone, which determines its biseasonal climate and unimodal annual pattern of precipitation. From 1987 to 2004 mean annual precipitation was 6849mm and monthly precipitation was below average between December and April (lowest in March) and above average most of the rest of the year (highest in June). Sea surface temperatures around Gorgona vary between 26 and 29ºC, but occasionally decrease to <19ºC during upwelling events that occur at the beginning of the year (Prahl et al. 1979, Díaz et al. 2001, Zapata 2001), or increase up to 30-32ºC during anomalous conditions associated with El Niño events (Vargas-Ángel et al. 2001). As is typical of coastal waters in the region, seawater salinity at Gorgona is relatively low and variable (29-33) compared to that of the open ocean, but nonetheless it is higher and more stable than that of the mainland coast, which is usually <20ºC (Prahl 1986). The tidal regime at Gorgona is semidiurnal with a maximum vertical range of ~5.7 m. Climatic and oceanographic conditions are described in greater detail by Díaz et al. (2001) and Giraldo et al. (2008) for Gorgona, and by Devis-Morales et al. (2002) for the Colombian Pacific basin. Besides strong El Niño warming events that periodically (on average once every 13 years) cause severe coral bleaching and mortality (Glynn 1990, Vargas-Ángel et al. 2001), the most significant physical perturbation on Gorgona Island coral reefs are repeated aerial exposures during extreme low tides. These occur irregularly, on average every three months (although intervals between consecutive events vary between one and 15 months), and occasionally cause significant coral bleaching and mortality, particularly on the shallowest portions of reefs (Zapata et al. 2001, Zapata & Vargas-Ángel 2003).
Monitoring stations: La Azufrada coral reef, located on the leeward side of the island, is the largest (11.2ha) continuous reef of Gorgona. It is a 1km long fringing reef with an extensive (~80-130m) reef flat built mainly by pocilloporid corals, and a sloping reef front and outer reef base where massive species of the genus Pavona Lamarck, 1801 and Gardineroseris Scheer and Pillai, 1974 occur, often in patches (Glynn et al. 1982, Zapata 2001, Zapata & Vargas-Ángel 2003). Following the established protocol for SIMAC (Garzón-Ferreira et al. 2002b, Garzón-Ferreira & Rodríguez-Ramírez 2010) two study sites were selected on this reef in November 1998 based on their good state of conservation, and similar degree of reef development. The first site (La Azufrada North or AN; Fig. 1) is located in the Northern section of the reef near a circular, crater-like depresion, which is a notable, persistent feature of this reef (Glynn et al. 1982, Zapata 2001). The second site (La Azufrada South or AS) is located in the Southern section of the reef about 400m from AN (Fig. 1). More detailed information on the two sites may be found in Garzón-Ferreira et al. (2002b).
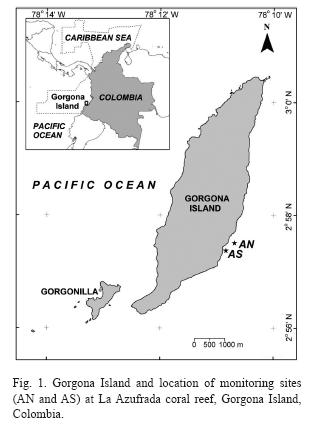
Assessment of benthic cover: At each site, five 10m long transects were haphazardly laid out and permanently marked at each of two depths. The shallow plot was located on the reef flat at ~0.5m below the mean lower low water (MLLW) tidal datum, while the deep plot was located at the interface between the reef front and the outer reef base at ~4m below MLLW. Data on the benthic cover were collected once a year between 1998 and 2004, during a single week of September through November, depending on the year, to minimize the seasonal variability of certain components such as the algae (Bula-Meyer 1990, Díaz-Pulido & Garzón-Ferreira 2002), even though at Gorgona seasonality in algal abundance is not marked (Bula-Meyer 1995). The continuous line intersept transect method (Loya 1978) was used to estimate the percentage of the substrate covered by different benthic components following a protocol modified from CARICOMP (1997, 2001; see also Garzón-Ferreira et al. 2002b, Garzón-Ferreira & Rodríguez-Ramírez 2010).
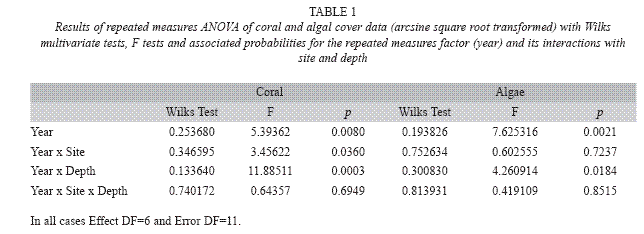
The statistical analysis was applied only to coral and algal components because they comprised >80% of the substrate in 90% of transects (N=140). Data on the percentage of cover were transformed prior to analysis as X'= arcsin √p, where p is the proportion of a given substrate category in a transect (Zar 1996). A repeated measures ANOVA was used to analyze these data because the same sampling units (transects) were repeatedly sampled every year. This design thus included one within-subject, repeated measures factor (year) with seven levels representing the number of consecutive years from 1998 to 2004, and two additional factors, each with two levels: site (AN and AS) and depth (shallow and deep plots). Because we were interested in comparing the patterns of temporal variation in coral and algal cover at different sites and depths, we focused the analysis on the year factor and its interactions. These were analyzed using the multivariate approach to repeated measures ANOVA (Wilks test) because this method does not rest on the assumptions of sphericity and compound symmetry (StatSoft 2003).
Results
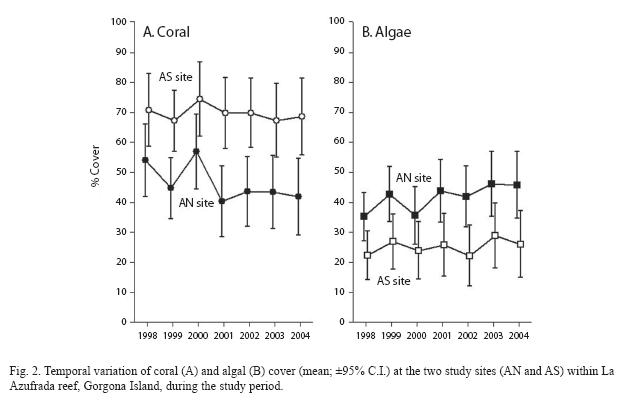
Temporal and spatial variation in coral and algal cover: Coral cover was persistently higher and algal cover lower on the Southern site than on the Northern site throughout the study period (Fig. 2). Mean coral cover varied between 67.2±17.1% and 74.5±20.1% at the Southern site in comparison with values ranging between 40.3±29.1% and 56.9 ±23.6% at the Northern site. Although overall temporal patterns of variation in coral cover tended to be similar at both sites, differences between sites were not constant through time (significant year x site interaction; Table 1). Such differences were smallest in 1998 and largest in 2001 (Fig. 2A). Patterns of variation in algal cover were the opposite of those exhibited by coral. Mean algal cover varied between 22.2±19.5% and 28.9±19.5% at the Southern site while it ranged between 35.2±15.8% and 46.1±26.3% at the Northern site. However, the temporal pattern of variation in algal cover did not differ between sites (Table 1, Fig. 2B).
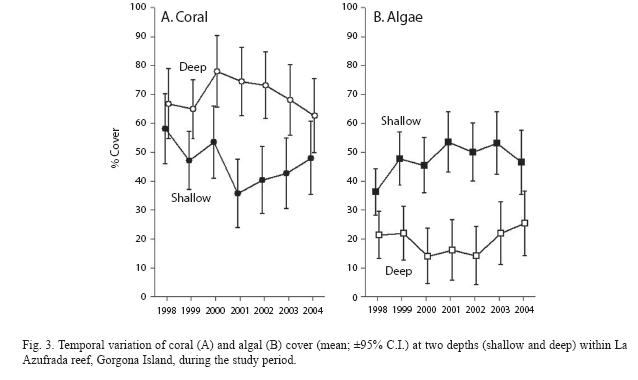
Most notable were the patterns of variation in coral and algal cover in relation to depth. In particular, depth levels exhibited distinct and opposing temporal patterns and therefore a significant depth by year interaction in both cases (Table 1, Fig. 3): while in deep plots coral cover tended to increase toward the middle of the study period and then decline, it tended to decline and then increase in shallow plots (Fig. 3A). This pattern was reversed for algal cover (Fig. 3B). In both cases, differences between depths were largest in 2001 reaching lowest values of coral (35.7±20.9%) and highest values of algal cover (53.5±17.5%) on shallow areas. Finally, depth differences in coral cover showed similar temporal patterns at both sites, while the same was true for algal cover. This was indicated for both variables by the lack of a significant year x site x depth interaction (Table 1).
Discussion
A great deal of effort has been devoted to study the stability and resilience of coral reef communities, that is, the persistence of community properties and the ability of communities to withstand perturbation, respectively (Connell 1978, 1997, Connell et al. 1997, 2004, Nystrom et al. 2000, Halford et al. 2004, Gardner et al. 2005). However, assessing stability and resilience on coral reefs has been difficult for at least two reasons: 1) there is a lack of adequate long-term data to provide a basis for assessments, particularly in less-developed countries where most coral reefs are located, and 2) different criteria have been used to determine the degree of stability and resilience of coral reef communities. While other characteristics (e.g., species richness and diversity, recruitment strength, colony size structure, growth form diversity, structural complexity and degree of framework development) can be very useful to assess stability, resilience and generally the health of coral reefs (Brown 1997a), here we focused on coral and algal cover because they are two of the most important components of the benthic community on coral reefs, they often compete strongly for primary space (e.g., Tanner 1995, McCook et al. 2001), and their relative cover can be used as an indicator of reef health. Although our study covers a period of only seven years, it is one of the first efforts to begin the description of the long-term dynamics of reef corals and algae in the TEP (but see also Eakin 2001, Guzmán & Cortés 2001, Jiménez & Cortés 2003), and provide a basis for future, longer-term studies. Here we discuss the temporal variation in coral and algal cover in relation to spatial (site and depth) factors in the light of specific natural and anthropogenic disturbances, which we hypothesize, are important drivers of the observed dynamics. Finally, we use the results of this study to assess the current conservation status of La Azufrada coral reef by comparing them with previous descriptions of this reef.
Temporal dynamics in coral and algal cover: Several major features of the temporal patterns of variation on the reef at La Azufrada were apparent in this study: 1) coral and algal cover showed significant inter-annual variation during a relatively short period of seven years (maximum absolute differences between any two years in mean coral and algal cover of <11% and <9%, respectively). 2) There was a net decrease in overall mean coral cover from 62% to 55%, and a concomitant net increase in overall algal cover from 29% to 36% from the beginning to the end of the study. 3) Temporal variation in coral and algal cover varied according to specific site and depth differences indicating the occurrence of persistent spatial effects on the temporal dynamics.
Two major natural perturbations that can affect the dynamics of coral and algal abundance intermittently affect Gorgona Island's coral reefs: low frequency, strong El Niño warming events, and high frequency, sub-aerial exposures during extreme low tides, both of which can cause significant bleaching and mortality (Zapata et al. 2001, Zapata & Vargas-Ángel 2003). Although no significant El Niño events occurred during the study period, the study began at the end of the 1997-98 El Niño, when Gorgona Islands coral reefs were showing clear signs of recovery from a bleaching event (Vargas-Ángel et al. 2001). Levels of bleaching reported for La Azufrada were at the most 32% of live coral cover on the reef slope at the peak of this event even though the magnitude of SST anomalies (up to 3.3ºC above the long-term mean) and the duration of the event were similar to those of the 1982-83 El Niño, which caused up to 85% coral mortality (Prahl 1983, Glynn 1990, Vargas-Ángel et al. 2001). However, during the 1997-98 event coral mortality at Gorgona was low (<1%), although greater in agariciids than in the dominant pocilloporids (Vargas-Ángel et al. 2001). Nonetheless, it is plausible that the observed overall mean decline of 6% in coral cover from 1998 to 1999 may have been the result of direct or indirect longer-term consequences of the 1997-98 El Niño disturbance. Such a decline is comparable to the levels of coral mortality reported as a consequence of this disturbance on other coral reefs in the TEP except those in Galápagos: 5% at Caño Island, Costa Rica (Guzmán & Cortés 2001), 7% on mainland Ecuador and 13% in the Gulf of Chiriquí, Panama (Glynn et al. 2001).
Sub-aerial exposure of coral reefs during extreme low tides is a phenomenon that occurs frequently at Gorgona as it does elsewhere in the TEP (Glynn 1976, 2003, Eakin & Glynn 1996, Zapata et al. 2001, Cortés & Jiménez 2003, Maté 2003, Zapata & Vargas-Ángel 2003). This phenomenon likely explains much of the dynamics in coral and algal cover and the differences between depths observed in this study (Fig. 3). According to an estimate made at La Azufrada in 1993, tidal emersion of coral occurred when water levels fell below ~–0.4 m relative to the MLLW datum. However, more recent observations suggest that coral tidal emersion may occur during less extreme low tides (~–0.3 m), probably as a result of upward coral growth during the last decade. Not all aerial exposures cause coral mortality, but those that are prolonged (>1h), occur at mid-day or otherwise coincide with unfavorable weather conditions (e.g., heavy rain, Maté 2003) and occur repeatedly during consecutive days are more likely to do so (Glynn 1976, Zapata & Vargas-Ángel 2003). Tidal emersion events and their consequences for coral reefs at Gorgona are similar to those described for Panamá (Glynn 1976, Zapata & Vargas-Ángel 2003). Soon after tissue death, the affected portions of colonies are colonized by filamentous algae whose continued growth, can in a few weeks, cover and kill entire colonies, therefore creating algal patches of varying sizes, depending on reef topography. Thus, generally the effect of aerial tidal emersion is a decrease in coral cover and a concomitant increase in algal cover on shallow areas.
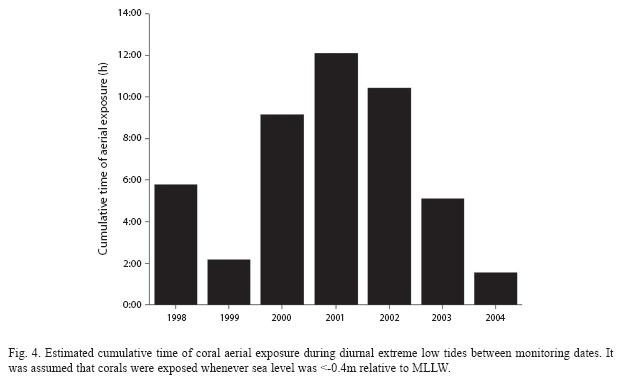
We suggest that the decline in coral and increase in algal cover from 1998-2001 on shallow areas (Fig. 3) can be explained by tidal emersion events. Using tide predicting software (WXTide32) we calculated the cumulative time that sea level was <-0.4 m relative to MLLW during daytime between successive monitoring dates. This was particularly long in 2000, 2001 and 2002 (Fig. 4). Tide tables also indicated that particularly extreme low tidal emersion events occurred repeatedly (once a month) during March 2001. Each event consisted of three consecutive days of mid-day (10:30 a.m. to noon) extreme low tides ranging from -0.4 to -0.58 m relative to MLLW (IDEAM 2000) and lasting between 0.5 and 2.0h each day. These events likely caused significant coral mortality as each event probably exacerbated the effects of the previous one and would thus explain the marked decline in coral cover observed in shallow areas from 2000 to 2001. Although the changes in coral cover observed in both 1999-2000 and 2001-2002 in shallow areas (Fig. 3) would seem to contradict this argument, these were not significant (Tukey HSD tests). Furthermore, aerial exposures in 2002 were unlikely to further reduce coral cover because corals exposed that year probably had already died during the tidal emersion events of 2001. Finally, it is interesting that coral cover on shallow areas seems to be returning to levels observed in 1998 after changes that occurred primarily in 2001 (Fig. 3). This recovery is consistent with a decrease in the cumulative time of aerial exposure since 2003 (Fig. 4). Thus, temporal changes in coral and algal cover are consistent with the hypothesis that tidal emersion events are a major driver of the dynamics of these two biotic components on shallow areas of the reef at La Azufrada. In contrast, the declining trend in coral and increasing trend in algal cover on deep plots after 2001 (Fig. 3) are difficult to explain by the effects of tidal emersion, and plausible causes of these trends remain unknown.
The temporally consistent differences between sites (in spite of a weak but significant year x site interaction in coral but not algal cover; (Table 1) indicate that such differences must precede this study (Fig. 2). Differences between sites must be the result of a factor that acts chronically to hamper an increase in coral cover in the northern site, particularly at shallow depths. Sedimentation stress as a result of terrestrial runoff may be one such factor, although not necessarily the only one. Coral mortality due to sedimentation has been reported at La Azufrada reef since the first studies (Prahl et al. 1979, Glynn et al. 1982), particularly in the Northern half of the reef in front of a site formerly know as El Aeropuerto, where the AN site was located. The forest adjacent to this site was extensively cleared for the construction of a landing strip for a penal facility built in Gorgona in the early 1960s. Coral burial by sediment has been frequently observed on the inner edge of the reef (Prahl et al. 1979), even many years after natural reforestation began (Zapata et al. 2001) and still today. Although sedimentation rates at La Azufrada have not been studied in detail, preliminary measurements (Lozano et al. unpublished data) clearly indicate that sedimentation is greater on the back reef (95g/m2.d) than on the outer slope (34g/m2.d). Furthermore, the prevalence of Northward-moving coastal currents (Díaz et al. 2001) that can transport sediments during periods of heavy rainfall from a major freshwater stream mouth located in the middle of La Azufrada may explain the consistent differences in coral and algal cover between the Northern and Southern study sites on both shallow and deep plots. Thus sedimentation stress may explain differences in coral and algal cover between sites but does not appear to significantly affect their temporal dynamics.
In conclusion, La Azufrada coral reef has shown significant temporal variation in two of its major biological components (corals and algae) over a relatively short period of time (1998-2004). Some of this variation appears to have been caused by the 1997-98 El Niño, which was likely responsible for a small but significant decline in coral cover during the first year of the study. Temporal and depth-related variation in the abundance of coral and algae is also partially explained by low tidal emersions, while differences between sites appear to be related to sedimentation stress. Although shallow areas, which are apparently most affected by low tidal emersions, are showing a gradual recovery of coral cover, deep areas are exhibiting a recent decline in coral cover due to an unidentified factor. While the SIMAC monitoring program has permitted a description of the temporal dynamics of La Azufrada coral reef and, to some extent, the identification of plausible causes of change, more studies are needed to identify what are the mechanisms mediating coral mortality, coral recovery and resilience on this reef.
Conservation status of La Azufrada coral reef: In comparison with coral reefs elsewhere, overall levels of coral cover were high (55-66%) and algal cover low (30-37%) during the course of this study, though not unusual for this coral reef. Since the implementation of the SIMAC monitoring program in 1998, Gorgona Islands coral reefs have consistently shown the highest coral cover of all the monitored reefs in Colombia (Rodríguez-Ramírez et al. 2006). However, high coral cover is not unusual among healthy TEP coral reefs, particularly those built by pocilloporids, which tend to form dense carpets. Other coral formations in the Colombian Pacific also exhibit relatively high live coral and low algal cover (e.g., 57.5% and 34.1%, respectively on an Ensenada de Utría reef and 49.6% and 36.4%, respectively, on a Malpelo Island reef; Rodríguez-Ramírez et al. 2005, 2006). Additionally, these reefs usually have few other occupants of primary space (e.g., sponges, milleporids or soft corals).
Overall mean levels of coral and algal cover at La Azufrada in 2004 were not different from levels observed before the beginning of SIMAC. Based on only seven transects made (Glynn et al. 1982), overall live coral cover recorded in 1979 was between 36% (all transects considered) and 51% (only transects with coral at depths <11m, n=5). No estimates of algal cover were reported in that study. Additional data collected on varying numbers of haphazardly laid 10m transects (Zapata et al. unpublished data) indicate that overall live coral and algal cover were, respectively, 56% and 36% in 1992 (n=12), 70% and 26% in 1995 (n=18), and 57% and 42% in 1996 (n=57). Because coral cover varies significantly among reef zones, being highest on the reef crest and front, and lowest on the back reef and slope (with algal cover showing the opposite pattern), the particularly high values recorded in 1995 may reflect some bias in the location of transects in areas of high coral cover (crest and front). Nonetheless, it is clear that by 1992 coral cover at La Azufrada had already recovered to predisturbance levels (1979) after the 1982-83 El Niño. Thus, the available long-term data suggest that, in comparison with other TEP coral reefs, La Azufrada reef has exhibited a notable degree of resilience in the face of recurrent natural perturbations.
The slightly decreasing trend in coral and increasing trend in algal cover observed during the study period are suggestive enough to warrant continued monitoring of La Azufrada reef for a longer period. Since this study did not allow the examination of processes occurring over longer time scales that may be affecting this reef, and given the recurrent nature of the major natural perturbations that affect its coral community, it is important to establish with greater certainty whether coral cover is temporally stable, declining slowly or exhibiting cyclic dynamics. Although the recent history of La Azufrada reef since coral reef studies began at Gorgona Island three decades ago (Prahl et al. 1979, Glynn et al. 1982) suggest a remarkable ability of this reef to recover from past perturbations, the frequency and magnitude of both natural and anthropogenic threats are likely to continue increasing as they are worldwide (Wilkinson 2002, Gardner et al. 2003, Hughes et al. 2003). Their small size, patchy occurrence and modest development under marginal conditions (Guzmán & Cortés 1993, Cortés 1997, Glynn &Ault 2000) make TEP coral reefs unique but extremely vulnerable to both natural and anthropogenic disturbances as the near disappearance of coral reefs in the Galápagos Islands after the 1982-83 El Niño warming so well illustrates (Glynn 2003).
Acknowledgments
The development and implementation of SIMAC (National Monitoring System for the Coral Reefs of Colombia) has been possible at Gorgona thanks to the support of the Institute for Marine and Coastal Research (INVEMAR), the Colombian Institute of Science and Technology (COLCIENCIAS) through grant Colciencias-BID 2105-09-327-97, the Regional Coordination Unit for the Caribbean of the United Nations Environment Program (UNEP- CAR/RCU), the National Environmental Fund (FONAM) through Environmental Program Agreement BID-7740C/CO, the Special Administrative Unit of the System of National Natural Parks (UAESPNN) of the Ministry of the Environment, Housing and Territorial Development, and the Coral Reef Ecology Research Group of Universidad del Valle. We are grateful for the decisive support of UAESPNN personnel at Gorgona, particularly Claudia Acevedo, Camilo Gómez, Gustavo Mayor and Luis Chasqui. The latter also made available to us an unpublished analysis of Gorgonas metereological data. We also thank Sven Zea for advice, and P. Herrón, D. Gil, J. Rojas, J. Pinzón, M.C. Reyes-Nivia, R. Navas, S. Bejarano, S. Lozano, C. Gómez, M.F. Maya, N. Bernal, M. López, M. Rodríguez, L. Chasqui and C. Muñoz for help with field work. We are grateful to Adolfo Salinas, Pablo Montoya and Harold Botero for significant discounts on diving and transportation costs. This manuscript benefited from critical comments made by S. Zea and two anonymous reviewers. Contribution No. CTBR-1044 from INVEMAR.
References
Aronson, R.B. & W.F. Precht. 1997. Stasis, biological disturbance, and community structure of a Holocene coral reef. Paleobiology 23: 326-346. [ Links ]
Aronson, R.B., W.F. Precht, M.A. Toscano & K.H. Koltes. 2002. The 1998 bleaching event and its aftermath on a coral reef in Belize. Mar. Biol. 141:435-447. [ Links ]
Birkeland, C. 1997. Life and death of coral reefs. Chapman & Hall, New York, New York, USA. [ Links ]
Brown, B.E. 1997a. Disturbances to reefs in recent times, p. 354-379. In C. Birkeland (ed.). Life and death of coral reefs. Chapman & Hall, New York, New York, USA. [ Links ]
Brown, B.E. 1997b. Coral bleaching: causes and consequences. Proc. 8th Int. Coral Reef Symp. 1: 65-74. [ Links ]
Bryant, D., L. Burke, J. McManus & M. Spalding. 1998. Reefs at risk: a map-based indicator of threats to the worlds coral reefs. World Resources Institute, Washington D.C., USA. [ Links ]
Buddemeier, R.W., J.A. Kleypas & R.B. Aronson. 2004. Coral reefs and climate change. Potential contribution of climate change to stresses coral reef ecosystems. Pew Center on Global Climate Change, Arlington, Virginia, USA. [ Links ]
Bula-Meyer, G. 1990. Altas temperaturas estacionales del agua como condición disturbadora de las macroalgas del Parque Nacional Natural Tayrona, Caribe colombiano: una hipótesis. An. Inst. Invest. Mar. Punta Betín 19-20: 9-21. [ Links ]
Bula-Meyer, G. 1995. Macroalgas de la Isla de Gorgona (Pacífico Colombiano) con nuevos registros y una explicación de la baja diversidad y biomasa. Instituto de Ciencias Naturales, Museo de historia Natural, Universidad Nacional de Colombia. Biblioteca José Jerónimo Triana 11: 23-45. [ Links ]
CARICOMP.1997. CARICOMP monitoring of coral reefs. Proc. 8th Int. Coral Reef Symp. 1: 651-656. [ Links ]
CARICOMP. 2001. CARICOMP Methods manual, levels 1 and 2: Manual of methods for mapping and monitoring of physical and biological parameters in the coastal zone of the Caribbean. CARICOMP Data Management Center, Univ. of the West Indies, Jamaica. [ Links ]
Carpenter, R.C. 1990. Mass mortality of Diadema antillarum. I. Long-term effects on sea urchin population dynamics and coral reef algal communities. Mar. Biol. 104: 67-77. [ Links ]
Connell, J.H. 1978. Diversity in tropical rain forests and coral reefs. Science. 199: 1302-1310. [ Links ]
Connell, J.H. 1997. Disturbance and recovery of coral assemblages. Coral Reefs 16: S101-S113. [ Links ]
Connell, J.H., T.P. Hughes & C.C. Wallace. 1997. A 30-year study of coral abundance, recruitment, and disturbance at several scales in space and time. Ecol. Monogr. 67: 461-488. [ Links ]
Connell, J.H., T.P. Hughes, C.C. Wallace, J.E. Tanner, K.E. Harms & A.M. Kerr. 2004. A long-term study of competition and diversity of corals. Ecol. Monogr. 74: 179-210. [ Links ]
Cortés, J. 1997. Biology and geology of eastern Pacific coral reefs. Coral Reefs 16: S39-S46. [ Links ]
Cortés, J. & C. Jiménez. 2003. Corals and coral reefs of the Pacific of Costa Rica: history, research and status, p. 361-385. In J. Cortés (ed.). Latin America coral reefs. Elsevier Science B.V., Amsterdam, The Netherlands. [ Links ]
Devis-Morales, A., I. García-Hansen, I. Malikov & N.L. Villegas-Bolaños. 2002. Compilación oceanográfica de la cuenca Pacífica colombiana. Serie Publicaciones Especiales Vol. 1, Centro Control Contaminación del Pacífico (CCCP), Tumaco, Colombia. [ Links ]
Díaz, J.M., J. Garzón-Ferreira & S. Zea. 1995. Los arrecifes coralinos de la Isla de San Andrés, Colombia: Estado actual y perspectivas para su conservación. Acad. Colomb. Cien. Exac. Fis. Nat., Colec. Jorge Alvarez Lleras 7, Bogotá, Colombia. [ Links ]
Díaz, J.M., Díaz-Pulido, G.; Garzón-Ferreira, J., Geister, J., Sánchez, J. & Zea, S. 1996. Atlas de los arrecifes coralinos del Caribe colombiano. Complejos arrecifales oceánicos. INVEMAR. Serie de Publicaciones Especiales No. 2, Santa Marta, Colombia. [ Links ]
Díaz, J.M., L.M. Barrios, M.H. Cendales, J. Garzón-Ferreira, J. Geister, M. López-Victoria, G.H. Ospina, F. Parra-Velandia, J. Pinzon, B. Vargas-Ángel, F.A. Zapata & S. Zea. 2000. Áreas coralinas de Colombia. INVEMAR, Serie de Publicaciones Especiales No. 5, Santa Marta, Colombia. [ Links ]
Díaz, J.M., J.H. Pinzón, A.M. Perdomo, L.M. Barrios & M. López-Victoria. 2001. Generalidades, p. 17-26. In L.M. Barrios & M. López-Victoria (eds.). Gorgona Marina: Contribución al conocimiento de una isla única. INVEMAR, Serie de Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Díaz-Pulido, G. & J. Garzón-Ferreira. 2002. Seasonality in algal assemblages on upwelling-influenced coral reefs in the Colombian Caribbean. Bot. Mar. 45: 284-292. [ Links ]
Eakin, C.M. 2001. A tale of two ENSO events: carbonate budgets and the influence of two warming disturbances and intervening variability, Uva Island, Panama. Bull. Mar. Sci. 69: 171-186. [ Links ]
Eakin, C.M. & P.W. Glynn. 1996. Low tidal exposures and reef mortalities in the Eastern Pacific. Coral Reefs 15: 120. [ Links ]
Enfield, D.B. 2001. Evolution and historical perspective of the 1997-1998 El Niño-Southern Oscillation event. Bull. Mar. Sci. 69: 7-25. [ Links ]
Gardner, T.A., I.M. Côté, J.A. Gill, A. Grant & A.R. Watkinson. 2003. Long-term region-wide declines in Caribbean corals. Science 301: 958-960. [ Links ]
Gardner, T.A., I.M. Côté, J.A. Gill, A. Grant & A.R. Watkinson. 2005. Hurricanes and Caribbean coral reefs: impacts, recovery patterns, and role in long-term decline. Ecology. 86: 174-184. [ Links ]
Garzón-Ferreira, J. 1997. Arrecifes Coralinos: ¿Un tesoro camino a la extinción?. Colombia: Ciencia y Tecnología 15: 11-19. [ Links ]
Garzón-Ferreira, J. & A. Rodríguez-Ramírez. 2010. SIMAC: Development and implementation of a coral reef monitoring network in Colombia. Rev. Biol. Trop. 58 (Suppl. 1): 67-80. [ Links ]
Garzón-Ferreira, J. & M. Kielman. 1994. Extensive mortality of corals in the Colombian Caribbean during the last two decades, p. 247-253. In R.N. Ginsburg (ed.). Proceedings of the Colloquium on Global Aspects of Coral Reefs. Health, Hazards and History. RSMAS, University of Miami, Miami, USA. [ Links ]
Garzón-Ferreira, J. & S. Zea. 1992. A mass mortality of Gorgonia ventalina (Cnidaria: Gorgoniidae) in the Santa Marta area, Caribbean coast of Colombia. Bull. Mar. Sci. 50: 522-526. [ Links ]
Garzón-Ferreira, J., D.L. Gil-Agudelo, L.M. Barrios & S. Zea. 2001. Stony coral diseases observed in southwestern Caribbean reefs. Hydrobiologia 460: 65-69. [ Links ]
Garzón-Ferreira, J., J. Cortes, A. Croquer, H. Guzmán, Z. Leao & A. Rodríguez-Ramírez. 2002a. Status of coral reefs in southern tropical America in 2000-2002: Brazil, Colombia, Costa Rica, Panama and Venezuela, p. 343-360. In C. Wilkinson (ed.). Status of coral reefs of the World: 2002. Australian Institute of Marine Science (AIMS), Townsville, Australia. [ Links ]
Garzón-Ferreira, J., M.C. Reyes-Nivia & A. Rodríguez-Ramírez. 2002b. Manual de métodos del SIMAC. INVEMAR, Santa Marta, Colombia. [ Links ]
Ginsburg, R.N. (ed.). 1994. Proceedings of the colloquium on global aspects of coral reefs. Health, hazards and history. RSMAS, University of Miami, Miami, USA. [ Links ]
Giraldo, A., E. Rodríguez-Rubio & F.A. Zapata. 2008. Condiciones oceanográficas en Isla Gorgona, Pacífico oriental tropical de Colombia. Lat. Am. J. Aquat. Res. 36: 121-128. [ Links ]
Gladfelter, W.B. 1982. White band disease in Acropora palmata: Implication for the structure and growth of shallow reefs. Bull. Mar. Sci. 32: 639-643. [ Links ]
Glynn, P.W. 1976. Some physical and biological determinants of coral community structure in the Eastern Pacific. Ecol. Monogr. 46: 431-456. [ Links ]
Glynn, P.W. 1990. Coral mortality and disturbances to coral reefs in the tropical Eastern Pacific, p. 55-126. In P.W. Glynn (ed.). Global ecological consequences of the 1982-83 El Niño-Southern Oscillation. Elsevier, Amsterdam, The Netherlands. [ Links ]
Glynn, P.W. 1993. Coral reef bleaching: ecological perspectives. Coral Reefs. 12: 1-17. [ Links ]
Glynn, P.W. 2003. Coral communities and coral reefs of Ecuador, p. 449-472. In J. Cortés (ed.). Latin America coral reefs. Elsevier Science B.V., Amsterdam, The Netherlands. [ Links ]
Glynn, P.W. & J.S. Ault. 2000. A biogeographic analysis and review of the far Eastern Pacific coral reef region. Coral Reefs 19: 1-23. [ Links ]
Glynn, P.W., H. von Prahl & F. Guhl. 1982. Coral reef of Gorgona Island, Colombia, with special reference to corallivores and the influence on community structural reef development. An. Inst. Inv. Mar. Punta Betín 12: 185-214. [ Links ]
Glynn, P.W., J.L. Maté, A.C. Baker & M. Calderón. 2001. Coral Bleaching and mortality in Panama and Ecuador during the 1997-1998 El Niño-Southern oscillation event: Spatial and temporal patterns and comparisons with the 1982-1983 event. Bull. Mar. Sci. 69: 79-109. [ Links ]
Guzmán, H.M. & J. Cortés. 1993. Arrecifes coralinos del Pacífico Oriental Tropical: revisión y perspectivas. Rev. Biol. Trop. 41: 535-557. [ Links ]
Guzmán, H.M. & J. Cortés. 2001. Changes in reef community structure after fifteen years of natural disturbances in the Eastern Pacific (Costa Rica). Bull. Mar. Sci. 69: 133-149. [ Links ]
Halford, A., A. J. Cheal, D. Ryan & D. McB. Williams. 2004. Resilience to large-scale disturbance in coral and fish assemblages on the Great Barrier Reef. Ecology 85: 1892-1905. [ Links ]
Hallock, P., F.E. Müller-Karger & J.C. Halas. 1993. Coral reef decline. Nat. Geogr. Res. Explor. 9: 358-378. [ Links ]
Harvell, C.D., C.E. Mitchell, J.R. Ward, S. Altizer, A.P. Dobson, R.S. Ostfeld & M.D. Samuel. 2002. Climate warming and disease risks for terrestrial and marine biota. Science 296: 2158-2162. [ Links ]
Hughes, T.P. 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265: 1547-1551. [ Links ]
Hughes, T.P., A.H. Baird, D.R. Bellwood, M. Card, S.R. Connolly, C. Folke, R. Grosberg, O. Hoegh-Guldberg, J.B.C. Jackson, J. Kleypas, J.M. Lough, P. Marshall, M. Nyström, S.R. Palumbi, J.M. Pandolfi, B. Rosen & J. Roughgarden. 2003. Climate change, human impacts, and the resilience of coral reefs. Science 301: 929-933. [ Links ]
IDEAM. 2000. Pronóstico de pleamares y bajamares en la costa pacífica colombiana año 2001. Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM), Bogotá, Colombia. [ Links ]
Jackson, J.B.C., M.X. Kirby, W.H. Berger, K.A. Bjorndal, L.W. Botsford, B.J. Bourque, R.H. Bradbury, R. Cooke, J. Erlandson, J.A. Estes, T.P. Hughes, S. Kidwell, C.B. Lange, H.S. Lenihan, J.M. Pandolfi, C.H. Peterson, R.S. Steneck, M.J. Tegner & R.R. Warner. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science 293: 629-638. [ Links ]
Jennings, S. & N. Polunin. 1996. Impacts of fishing on tropical ecosystems. Ambio 25: 44-49. [ Links ]
Jiménez, C.E & J. Cortés. 2003. Coral cover change associated to El Niño, Eastern Pacific, Costa Rica, 1992-2001. P.S.Z.N. Mar. Ecol. 24: 179-192. [ Links ]
Lessios, H.A., D.R. Robertson & J.D. Cubit. 1984. Spread of Diadema mass mortality through the Caribbean. Science 226: 335-337. [ Links ]
Liddell, W.D. & S.L. Ohlhorst. 1992. Ten years of disturbance and change on a Jamaican fringing reef. Proc. 7th Int. Coral Reef Symp. 1: 144-150. [ Links ]
Loya, Y. 1976. Effects of water turbidity and sedimentation on the community structure of Puerto Rican corals. Bull. Mar. Sci. 26: 450-466. [ Links ]
Loya, Y. 1978. Plotless and transect methods, p. 197-217. In D.R. Stoddart & R.F. Johannes (eds.). Coral reefs: research methods. Monographs on oceanographic research 39. UNESCO, Norwich, England. [ Links ]
Maté, J.L. 2003. Corals and coral reefs of the Pacific coast of Panamá, p. 387-417. In J. Cortés (ed.). Latin America coral reefs. Elsevier Science B.V., Amsterdam, The Netherlands. [ Links ]
McCook, L.J., J. Jompa & G. Díaz-Pulido. 2001. Competition between corals and algae on coral reefs: a review of evidence and mechanisms. Coral Reefs 19: 401-417. [ Links ]
McWilliams, J.P., I.M. Côté, J.A. Gill, W.J. Sutherland & A.R. Watkinson. 2005. Accelerating impacts of temperature-induced coral bleaching in the Caribbean. Ecology 86: 2055-2060. [ Links ]
Munro, J.L. 1983. Caribbean coral reef fishery resources. ICLARM Studies and Reviews 7. Manila, Phippines. [ Links ]
Nystrom, M., C. Folke & F. Moberg. 2000. Coral reef disturbance and resilience in a human-dominated environment. Trends Ecol. Evol. 15: 413-417. [ Links ]
Pandolfi, J.M. 1999. Response of Pleistocene coral reefs to environmental change over long temporal scales. Am. Zool. 39: 113-130. [ Links ]
Pandolfi, J.M., R.H. Bradbury, E. Sala, T.P. Hughes, K.A. Bjorndal, R.G. Cooke, D. McArdle, L. McClenachan, M.J.H. Newman, G. Paredes, R.R. Warner & J.B.C. Jackson. 2003. Global trajectories of the long-term decline of coral reef ecosystems. Science 301: 955-957. [ Links ]
Prahl, H. von. 1983. Blanqueo masivo y muerte de corales en la Isla de Gorgona, Pacífico Colombiano. Cespedesia 12: 125-129. [ Links ]
Prahl, H. von. 1986. Corales y arrecifes coralinos, p. 61-87. In H. von Prahl & M. Alberico (eds.). Isla Gorgona. Biblioteca Banco Popular, Bogotá, Colombia. [ Links ]
Prahl, H. von, Guhl, F. & Grögl, M. 1979. Gorgona. Futura, Bogotá, Colombia. [ Links ]
Rodríguez-Ramírez, A., J. Garzón-Ferreira, S. Bejarano-Chavarro, R. Navas-Camacho, C. Reyes-Nivia, G. Duque, C. Orozco, F. Zapata & O. Herrera. 2005. Estado de los arrecifes coralinos en Colombia en el año 2004, p. 77-114. In INVEMAR. Informe del estado de los ambientes marinos y costeros en Colombia 2004. INVEMAR, Serie de Publicaciones Periódicas No. 8, Santa Marta, Colombia. [ Links ]
Rodríguez-Ramírez A., M.C. Reyes-Nivia, R. Navas-Camacho, S.Bejarano, J. Garzón-Ferreira & F. Zapata. 2006. Status of the coral reefs of Colombia in 2003. Proc. 10th Int. Coral Reef Symp. 1: 976-981. [ Links ]
Rogers, C.S. 1985. Degradation of Caribbean and Western Atlantic coral reefs and decline of associated fisheries. Proc. 5th Int. Coral Reef Symp. 6: 491-496. [ Links ]
Rouphael, A.B. & G.J. Inglis. 2002. Increased spatial and temporal variability in coral damage caused by recreational scuba diving. Ecol. Appl. 12: 427-440. [ Links ]
Santavy, D.L. & E.C. Peters. 1997. Microbial pests: coral disease in the Western Atlantic. Proc. 8th Int. Coral Reef Symp. 1: 607-612. [ Links ]
StatSoft, Inc. 2003. STATISTICA (data analysis software system), version 6. StatSoft, Tulsa, Oklahoma, USA. [ Links ]
Sutherland, K.P., J.W. Porter & C. Torres. 2004. Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Mar. Ecol. Prog. Ser. 266: 273-302. [ Links ]
Szmant A.M., 2002. Nutrient enrichment on coral reefs: Is it a major cause of coral reef decline? Estuaries 25: 743-766. [ Links ]
Tanner, J.E. 1995. Competition between scleractinian corals and macroalgae: An experimental investigation of coral growth, survival and reproduction. J. Exp. Mar. Biol. Ecol. 190: 151-168. [ Links ]
Vargas-Ángel, B., F.A. Zapata, H. Hernández & J. M. Jiménez. 2001. Coral and coral reef responses to the 1997-98 El Niño event on the Pacific coast of Colombia. Bull. Mar. Sci. 69: 111-132. [ Links ]
Wilkinson, C.R. 1992. Coral reefs of the world are facing widespread devastation: can we prevent this through sustainable management practices? Proc. 7th Int. Coral Reef Symp. 1: 11-21. [ Links ]
Wilkinson, C.R. 2002. Status of coral reefs of the world: 2002. Australian Institute of Marine Science, Townsville, Australia. [ Links ]
Williams, E.H. & L. Bunkley-Williams. 1990. The world wide coral reef bleaching cycle and related sources of coral mortality. Atoll. Res. Bull. 335: 1-71. [ Links ]
Wittenberg, M. & W. Hunte. 1992. Effects of eutrophication and sedimentation on juvenile corals: I. Abundance, mortality and community structure. Mar. Biol. 112: 131-138. [ Links ]
Zapata, F.A. 2001. Formaciones coralinas de isla Gorgona, p. 27-40. In L.M. Barrios & M. López-Victoria (eds.). Gorgona Marina: Contribución al conocimiento de una isla única. INVEMAR, Serie de Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Zapata, F.A. & B. Vargas-Ángel. 2003. Corals and coral reefs of the Pacific coast of Colombia, p. 419-447. In J. Cortés (ed.). Latin America coral reefs. Elsevier Science B.V., Amsterdam, The Netherlands. [ Links ]
Zapata, F.A., B. Vargas-Ángel & J. Garzón-Ferreira. 2001. Salud y conservación de las comunidades coralinas, p. 41-50. In L.M. Barrios & M. López-Victoria (eds.). Gorgona Marina: Contribución al conocimiento de una isla única. INVEMAR, Serie de Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Zar, J.H. 1996. Biostatistical analysis. Prentice-Hall, Upper Saddle River, New Jersey, USA. [ Links ]
Zea, S., J. Geister, J. Garzón-Ferreira & J.M. Díaz. 1998. Biotic changes in the reef complex of San Andrés Island (Southwestern Caribbean Sea, Colombia) occurring over nearly three decades. Atoll Res. Bull. 456: 1-30. [ Links ]
Corresponde a: Fernando A. Zapata. Departamento de Biología, Universidad del Valle, Apartado Aéreo 25360, Cali, Colombia; fernando.zapata@correounivalle.edu.coAlberto Rodríguez-Ramírez. Instituto de Investigaciones Marinas y Costeras (INVEMAR), Apartado Aéreo 1016, Santa Marta, Colombia; betorod@invemar.org.coCarlos Caro-Zambrano Instituto de Investigaciones Marinas y Costeras (INVEMAR), Apartado Aéreo 1016, Santa Marta, Colombia; cabetoz78@hotmail.com
Jaime Garzón-Ferreira. Instituto de Investigaciones Marinas y Costeras (INVEMAR), Apartado Aéreo 1016, Santa Marta, Colombia; jgarzon@invemar.org.co
Received 31-VIII-2009. Corrected 12-X-2009. Accepted 12-XII-2009.

