










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 suppl.1 San José Nov. 2009
Parasitism and predation of the planthopper Tagosodes orizicolus (Homoptera: Delphacidae) by a dryinid parasitoid in Costa Rica
Floria Mora-Kepfer1,2,4 & Ana Mercedes Espinoza2,3
1. Escuela de Biología, Universidad de Costa Rica, 11501-2060, San José, Costa Rica; floriamk@bio.miami.edu
2. Centro de Investigaciones en Biología Celular y Molecular, Universidad de Costa Rica, 11501-2060, San José, Costa Rica.
3. Escuela de Agronomía, Facultad de Ciencias Agroalimentarias, Universidad de Costa Rica, 11501-2060, San José, Costa Rica.
4. Department of Biology, University of Miami, Coral Gables, Florida 33124-0421; floriamk@bio.miami.edu
Abstract: We analyzed the effect of predation and parasitism by the native dryinid Haplogonatopus hernandezae as a natural enemy of the rice pest Tagosodes orizicolus under controlled conditions in an insectary in Costa Rica. We found no change in the proportion of T. orizicolus nymphs that were fed upon, parasitized, and that showed no apparent damage throughout the adult stage in H. hernandezae, which favors a stable control of the pest. The percentage of T. orizicolus nymphs that were parasitized (37 %) and preyed upon (36.5 %) was similar, eliminating 73,5 % of the total nymphs causing damage to the rice plants (N = 1 099). Of the total 324 parasitized nymphs, female nymphs were more parasitized than male nymphs (95 % and 5 % respectively) and dryinids developed more successfully in female host nymphs. These tendencies did not change with dryinid age. Survival of dryinids was higher in female T. orizicolus, both for larvae that emerged from the host and pupated, and for pupae that moulted to adults. In cages where the nymphs had no contact with dryinids we found a higher amount of males than females (57.44 ± 28.5 vs 45.22 ± 25.85). In contrast, when the nymphs were in contact with dryinids, the sex ratio was two females to one male reaching adulthood since the dryinids fed more on male nymphs (N = 692). Our results indicate that female dryinids prefer to oviposit in female T. orizicolus nymphs and prey on males. Reproduction by parthenogenesis, acceleration of the development in an insectary with controlled conditions and eficiency of H. hernandezae as a parasitoid and predator indicate that this dryinid is an effective biological control agent of T. orizicolus. Rev. Biol. Trop. 57 (Suppl. 1): 203-211. Epub 2009 November 30.
Key words: rice, biological control, Homoptera, Haplogonatopus hernandezae, parasitoid, Tagosodes orizicolus, Costa Rica.
The planthopper Tagosodes orizicolus (Muir) is a monophagous pest of rice, Oryza sativa (Lin), and the vector of the hoja blanca virus (RHBV) (Everett 1969, Meneses et al. 1998). T. orizicolus can cause severe crop losses when the rice variety planted is susceptible to the insect and to RHBV (Calvert & Reyes 1999, Peñaranda et al. 1999) and is found in all the rice growing regions in tropical America and the Caribbean (Morales & Niessen 1985).
Populations of T. orizicolus have previously been attacked by using synthetic insecticides (Rey & García 1980, Peñaranda et al. 1999). However, the use of synthetic insecticides has contributed to resistant insect selection (Corbett 1974), the elimination of native natural enemies (Mattenson 2000) and vast outbreaks of planthoppers due to overuse of insecticides in the 1970s (Gallagher et al. 1994, Heinrichs 1994, Mattenson et al. 1994).
The severe crop loss for rice farmers and the large-scale use of synthetic insecticides which cause highly toxic effects on living organisms make it indispensable to look for alternative strategies to control populations of T. orizicolus. A feasible strategy is the use of natural enemies of this pest as biological control agents.
To date, the studies on natural enemies of T. orizicolus have included parasitoids belonging to Gonatopodinae (Hymenoptera: Dryinidae). Several species of dryinids have demonstrated satisfactory results as biological control agents of T. orizicolus and other delphacids (Chandra 1980a, Chua & Dyck 1982, Hernández & Bellotti 1984, Olmi 1984, Virla & Olmi 1998) as they parasitize a specific group of hosts (Virla & Olmi 1998). Haplogonatopus vitiensis was successfully introduced to Hawaii from the Fiji Islands to control the populations of Perkinsiella saccharicida Kirk., a severe pest of sugar cane (Virla & Olmi 1998). Biological control of T. orizicolus using the dryinid Gonatopus sp. has replaced the use of synthetic insecticides in Cuba since 1987 (Meneses unpubl). In Colombia, doses of 100, 200 and 300 individuals of Haplogonatopus hernandezae per Ha of the crop were released to control the populations of T. orizicolus, reaching a parasitism of 83% of the pest population (Peñaranda et al.1999). H. hernandezae has been found in Costa Rica, Colombia and Argentina (Hernández & Belloti 1984, Olmi1995, Virla & Olmi 1998). This species reproduces parthenogenetically (Olmi 1984, Hernández & Belloti 1984), generating only females (Suomalainen 1962, Johanowicz & Hoy 1998, Jervis & Kidd 1996).
Females of H. hernandezae feed from secretions that contain sugar, and also prey on T. orizicolus nymphs. They choose to paralyze certain nymphs and oviposit on other nymphs where their larvae develop (Olmi 1984, Hernández & Belloti 1984). The female captures the nymph with her anterior legs whose tarsi have been modified as quelae and feeds from the tissues and hemolymph (Hernández & Belloti 1984). Previous studies indicate that predation on nymphs in addition to oviposition in other nymphs can lead to higher mortality and more effective biological control (Olmi 1984, Waloff 1974, Kidd & Jervis 1989, Jervis et al. 1996).
Although in a preliminary study Hernández & Belloti (1984) suggest the biological control potential of H. hernandezae as an enemy of T. orizicolus, little is documented about its impact as a parasitoid and predator and any preference if any on specific T. orizicolus nymphs. Since T. orizicolus females cause direct damage in the rice plants by laying eggs and transmitting the hoja blanca virus, it is indispensable to achieve a biological control strategy that targets females. To date there is no study that evaluates the effect of the sex of the T. orizicolus nymphal instar on the success in the development of the H. hernandezae larva.
In this study we analyze the effect of the dryinid H. hernandezae as a natural enemy of T. orizicolus, both as a predator and as a parasitoid. We also explore the effect of this dryinid on the proportion of T. orizicolus nymphs that are used for oviposition or as food, and the developmental success of H. hernandezae larvae in nymphs of each sex.
Materials and methods
Establishment of T. orizicolus and H. hernandezae colonies: Colonies of T. orizicolus in cages were established from nymphs and adults collected in rice fields in Guanacaste, Parrita and Turrúcares in Costa Rica, during 2000 and 2001. Nymphs and adults with visible signs of parasitism, such as a dark cyst on their thorax or abdomen, were isolated and reared in a small dryinid rearing cage (Chandra 1980b) to obtain H. hernandezae larvae that emerged from them to pupate on the rice leaves (Fig. 1.). The T. orizicolus colonies (Mora et al. 2007) and the dryinids were reared continuously in cages in an insectary under controlled conditions, humidity of 70-80% and temperature (25-30 °C). The T. orizicolus individuals in cages fed from Blue Bonnet 50 rice plants placed on a flat tray filled with 10 cm of water to exclude ants and other insects. The dryinid cages consisted of a cylindrical container placed on a rice plant (Chandra 1980b). Two or three rice leaves were placed inside the cage and held by foam taps in each end to avoid the escape of the dryinid. Individual dryinids were isolated in each container.
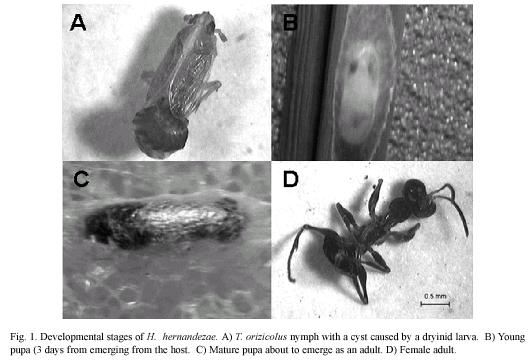
Parasitism and predation observations: We observed each of the containers with a single parasitized T. orizicolus nymph on a daily basis. We recorded the sex of the dryinid that emerged and the sex of the nymph that reached the adult stage, the duration of the complete life cycle and of each developmental stage. We also recorded the number of progeny for each dryinid, their survival rate, maximum developmental stage reached, and the quantity of parasitoid cysts in each nymph of T. orizicolus.
We introduced ten nymphs of instar 3-4 every day to each container with a single female dryinid. On the following day, the nymphs were kept in that container and the female dryinid was transferred to a new container with ten new nymphs, and so on until the dryinid died.
Because dryinids sometimes did not immediately kill nymphs on which they fed, we quantified the number of predated nymphs in each container and the remaining nymphs were observed for the next two days to register the total individuals that survived predation attacks. To ensure that this attack had caused the death of the host, we observed each nymph under a stereoscope, since the physical feeding is a clear indicator of the destructive host feeding.
We observed the remaining nymphs for eight days after they were exposed to the wasp to detect signs of parasitism (Fig. 1A). Parasitized hosts that reached the adult stage were kept in the same container with other planthoppers of the opposite sex that were not parasitized to determine if these adults were able to reproduce. They were then removed from the container and isolated only two to three days before the larvae emerged from the host.
We registered the number of days it took each nymph to reach the adult stage, and its sex as an adult, since it is not possible to distinguish the sex of the nymphs (Mora. et al . 2001). The age of each parasitoid at the moment it oviposited on the host, the developmental stage the parasitoid reached and whether it caused the death of the host were also recorded. To determine a control sex ratio of T. orizicolus using only nymphs that were not in contact with parasitoids, we had to wait for these nymphs to become adults and quantify the number of females and males in 9 rearing cages.
Statistical Analyses: To explore differences in the proportions of parasitism, predation and nymphs with no apparent damage according to the age of the dryinid female, we performed Repeated Measure ANOVAS. These proportions were analyzed after being expressed by using an arcosenic transformation (Zar 1996).
To determine if the sex proportion of T. orizicolus tends to a 1:1 relation, we performed a Wilcoxons test for matched pairs. We also explored differences in the sex ratio in potential nymphs in contact with a dryinid with a Chi-square test.
Results
We found individuals of T. orizicolus parasitized by H. hernandezae in all three rice fields in the different geographical locations that we visited. All the dryinids collected from the rice fields (N=64) and the progeny that were reared in controlled conditions (N=228) were female, and virgin females in captivity produced fertile female offspring by parthenogenesis.
A total of 228 descendants were obtained from dryinid females over seven generations. Each female produced an average of 12 ±2.2 offspring of which an average of 9 ±1.56 individuals reached the adult stage (N=47).
Of the T. orizicolus nymphs raised in the insectary, 22 were superparasitized, with two cysts in each individual. In 73% of those that were superparasitized, both larvae emerged from the host to pupate on the rice leaves: only one larva survived in 18% of the cases, no larvae emerged from the host in the remaining 9%.
The duration of most developmental stages of H. hernandezae reared in controlled temperature conditions decreased in comparison to those reared in natural conditions. However, the average duration of the adult stage in female dryinids did not vary with respect to the temperature at which were reared in (Table 1).
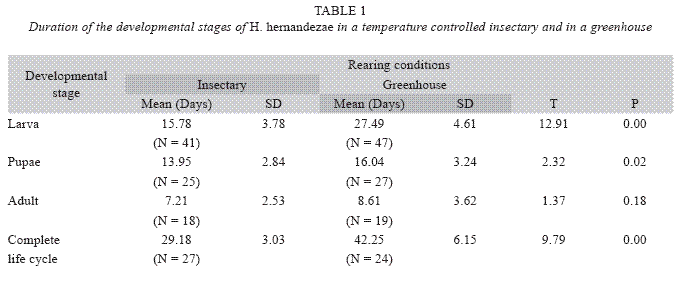
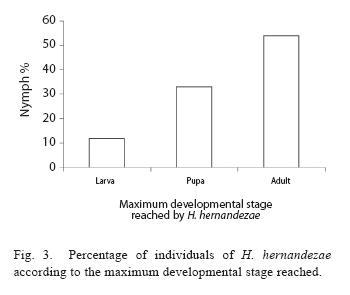
A total of 1099 nymphs were in contact with 40 female adult dryinids, of which 73,5% of the nymphs were attacked successfully by the dryinid (Fig. 2), and were either parasitized (36.5%) or fed upon (37%). Of the total dryinid larvae, 87% emerged from the nymph host and pupated on the rice leaves, causing the death of the nymph. The remaining 13% died as larvae inside the nymph, and 33% as pupae. Only 54% of the dryinid larvae that produced cysts were able to emerge from the nymph and reach the adult stage (N=407; Fig. 3).
None of the parasitized nymphs that reached the adult stage in contact with other non-parasitized nymphs of the opposite sex was able to reproduce. In contrast, in four cases we obtained progeny from individuals that were exposed to a female dryinid but were not parasitized.
The proportions of parasitized, consumed and unconsumed nymphs did not vary with respect to host age (F=0.008, d.f.=6, P>0.05, F=0.0188, d.f.=6, P>0.05; F=0.0188, d.f.=6, P>0.05 respectively, N=1099; Fig. 4). The proportion of male nymphs was significantly higher than females, 57.44±28.5 vs 45.22±25.85, in rearing cages in which the nymphs had no contact with wasps (T=4.5, d.f.=8, P=0.03, N=1063). In contrast, the sex ratio varied in the case of planthoppers that were in contact with a parasitoid during the nymphal stages, the proportion of female adults was 2 for each male that reached adulthood (χ2 = 94.88 , d.f.=1, N=692, P<0.05).
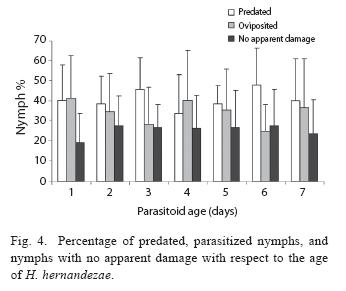
Female nymphs were also more parasitized than male nymphs (χ2=43.61, d.f.=1, P =0.001, Fig. 5) and wasps developed more successfully in female host nymphs. Larvae in female hosts were also more successful in emerging from the host to pupate. (χ2 =10.57, d.f.=1, P=0.001; Fig. 5.), as were pupae in females hosts that moulted into adults ( χ2 =8.74, d.f.=1, P =0.003).
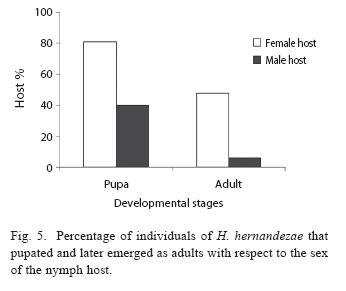
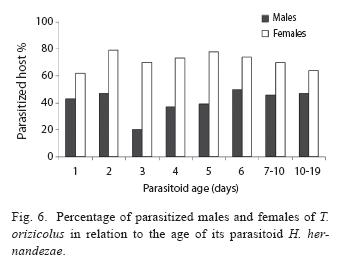
Parasitoids that emerged from female hosts (78,8%), were significantly more successful in completing their life cycle than those that emerged from male hosts (χ2=26.06, d.f.=1, P=0.00, N=392). The tendency to preferentially
attack female nymphs did not vary with parasitoid age (χ2=11.43, d.f.=8, P=0.178; Fig. 6).
Discussion
The establishment of this dryinid in controlled conditions of temperature and humidity was not only successful but we were able to accelerate the developmental stages to reach the adult stage by using higher temperature conditions. However, these conditions did not alter the duration of the dryinid adult life span or their effectiveness as a control agent and we obtained a stable number of progeny per female during several generations. The number of nymphs attacked for parasitism and predation were similar and the effectiveness of the dryinid as a biological control agent did not change with age.
Even though only about 50% of the dryinid progeny reached the adult stage, 73% of the larvae emerged from the cyst to pupate causing the death of the nymph. These results show the same tendency as previous observations of Haplogonatopus larvae leaving their host (Hernández & Belloti 1984, Olmi 1984). None of the nymphs with visible signs of parasitism survived to adulthood even if the parasitoid did not survive enough to pupate outside the nymphal body. This result suggests that the death of the developing larvae caused the death of the nymph.
None of the parasitized nymphs that reached the adult stage while parasitized and had contact with individuals of the opposite sex, was able to reproduce. This could be the result of parasitic castration, which has been observed in other parasitoids (Olmi 1984, Dutt & Giri 1978). A previous study reported the same result with young nymphs of Pocatello furrier that were parasitized by Haplogonatopus apicalis, which were also unable to reproduce (Kitamura 1988). Thus, another advantage of Haplogonatopus hernandezae as a biological control agent is that the dryinid larvae will stop its host nymph from reproducing and cause its death.
The proportion of parasitized, predated and undamaged nymphs did not vary with respect to the age of the adult female of H. hernandezae. This contrasts with another study (Sahragard et al., 1991) which indicated that the number of individuals fed upon increased as the dryinid Dicondylus indianus became older and the number of hosts parasitized decreased.
The proportion of male and female Tagosodes orizicolus being parasitized also remained similar with dryinid age. However, we observed a bias towards more female nymphs than male nymphs being parasitized throughout the adult life of the dryinid female. This bias towards females indicates a higher predation of male nymphs and the use of female nymphs as hosts for the dryinid progeny. Visual differentiation of sex in T. orizicolus nymphs is not possible until the fifth nymphal instar, but there is a sexual dimorphism in size (Mora et al. 2001). Adult females are larger and therefore female nymphs may also be larger than male nymphs. For a female dryinid, to oviposit in a smaller host could have a negative effect on the developmental success of the progeny. Also, to feed from a small host could represent less energetic cost with respect to the capture, paralysis and host handling (Mora-Kepfer & Espinoza, unpublished). Female H. hernan-dezae feed from the thorax or abdomen causing death of the nymph by destructive host feeding. However, they do not consume the entire host (Mora-Kepfer, pers. obs.).
Our results also reveal that dryinid larvae that developed inside female nymphs were more successful in reaching the adult stage than those developing in male nymphs. Adult female T. orizicolus live an average of 31.1 days, while adult males live only an average of 14.6 days (Gómez-Sousa & Kamara 1980). Due to the shorter lifespan of males, dryinid larvae may not have enough time to develop and leave the nymph host to pupate.Since a dryinid larva can take up to 27 days to develop inside the host and emerge to pupate, ovipositing in third or fourth instar female nymphs would assure that a larvae will have enough time to develop and also cause the death of the nymph.
Future studies in rice fields are needed to confirm these findings in natural conditions. The relatively low number of H. hernandezae individuals found in rice fields may be due to the constant application of insecticides to control T. orizicolus populations. However, adult females and parasitized nymphs had a wide distribution, as they were found in every rice field that we visited. One essential aspect to further investigate in rice fields is the possible effect of hyperparasitoids attacking native populations of H. hernandezae.
In summary, our results support the feasibility of H. hernandezae as a biological control agent of T. orizicolus. Reproduction by parthenogenesis of this dryinid is also advantageous for rearing in captivity, as it eliminates possible problems regarding courtship, successful mating and sex ratios that many parasitoids suffer in massive rearing conditions (Ode & Heinz 2002) Another advantage of using H. hernandezae is the elimination of all the problems associated with the introduction of exotic species, their success in establishing themselves and possible interference with other insect species, since this dryinid is a native species in Costa Rica, (Jervis & Kidd 1996).
Acknowledgments
We thank Cynthia Barboza, Rebeca Mora, Miguel Muñoz and Erwin Gamboa for their help in the collection of field colonies and mantaining the planthopper and dryinid colonies kept in the insectary. We thank Carlos García-Robledo for statistical assistance. We also express our gratitude to Paul Hanson, William Eberhard, Massimo Olmi and Keith Waddington for reviewing and improving this manuscript.
Resumen
En un insectario en condiciones controladas analizamos los efectos de depredación y parasitismo del dryinido Haplogonatopus hernandezae, enemigo natural del vector del virus de la hoja blanca Tagosodes orizicolus. Nuestros resultados indican que la proporción de depredación y parasitismo es estable a lo largo de la vida del dryinido lo cual favorece un control continuo eliminando 73% de la plaga (N = 1 099). De un total de 324 ninfas de T. orizicolus parasitadas, las hembras fueron más parasitadas que los machos y las larvas del dryinido se lograron desarrollar exitosamente a partir de ninfas hembra en contraste con las ninfas macho. Encontramos una mayor proporción de machos en comparación con las hembras en las jaulas de T. orizicolus en que las ninfas no tuvieron ningún contacto con hembras del dryinido (57.44 ± 28.5 vs. 45.22 ± 25.85). Al contrario, cuando las ninfas estuvieron en contacto con las hembras del dryinido, la proporción de sexos fue de dos hembras por cada macho. Este resultado indica que las hembras del dryinido prefieren ovipositar en las hembras de T. orizicolus y alimentarse de las ninfas macho. La reproducción por partenogénesis, aceleración de las etapas de desarrollo en condiciones controladas, y la eficiencia como parasitoide y depredador de este dryinido indican su efectividad como agente de control biológico de T. orizicolus.
Palabras clave: arroz, control biológico, depredador, Haplogonatopus hernandezae, parasitoide, Tagosodes orizicolus.
Received 03-X-2007. Corrected 09-X-2008. Accepted 18-XI-2008.
References
Calvert, L. & L.A. Reyes. 1999. Manejo del complejo "Sogata-Virus de la Hoja Blanca" en el cultivo del arroz. CIAT-FEDEARROZ. [ Links ]
Chandra, G. 1980a. Dryinid parasitoids of rice planthoppers and planthoppers in the Philippines. I. Taxonomy and bionomics. Acta Oecol. Appl. 1: 161-172. [ Links ]
Chandra, G. 1980b. Dryinid parasitoids of rice planthoppers and planthoppers in the Philippines. II. Rearing techniques. Entomophaga 25: 187-192. [ Links ]
Chua, T.H. & V.A. Dyck. 1982. Assessment of Pseudogonatopus flavifemur E. & H. (Dryinidae: Hymenoptera) as a biocontrol agent of the rice brown planthopper. Proc. Int. Conf. in Tropics 253-265. [ Links ]
Corbett, J.R. 1974. The biochemical mode of action of pesticides. Academic, New York. In R.F. Denno and C. J: Perfect. (Ed). Planthoppers, their ecology and management. Chapman and Hall, New York, USA. [ Links ]
Dutt, N. & P. Giri. 1978. A dryinid parasite of brown planthopper of paddy, Nilaparvata lugens Stal (Homoptera: Delphacidae). Sci. & Cult. 44: 3, 127-128. [ Links ]
Everett, T. 1969. Hoja blanca, p. 361-377 In K. Maramorosch (ed.). Viruses, vectors andvegetation. John Wiley and Sons, USA. [ Links ]
Gallagher, K.D., P.E. Kenmore & K. Sogawa. 1994. Judicial use of insecticides deter planthopper outbreaks and extend the life of resistant varieties in Southeast Asian Rice, p. 599-614 In R.F. Denno & J.T. Perfect (eds.). Planthoppers: Their ecology and Management. Chapman and Hall, New York, USA. [ Links ]
Gómez-Sousa, J. & F. Kamara. 1980. Determinación de algunos parámetros en la biología de Sogatodes orizicola (Muir). Centro Agrícola 7: 13-20. [ Links ]
Heinrichs, E.A. 1994. Impact of insecticides on the resistance and resurgence of rice planthoppers, p. 571-598. In R.F. Denno & C.J. Perfect. (eds.). Planthoppers: their ecology and management. Chapman and Hall, New York, USA. [ Links ]
Hernández M.P. & A. Belloti. 1984. Ciclos de vida y hábitos de Haplogonatopus hernadezae Olmi (Hymenoptera: Dryinidae) controlador natural del delfácidos del arroz Sogatodes orizicola (Muir). Rev. Colombiana Ent. 10: 3-8. [ Links ]
Jervis, M. & N.A.C. Kidd. 1996. Insect Natural Enemies. Practical approaches to their study and evaluation. Chapman and Hall, London, England. [ Links ]
Kitamura, K. 1988. Comparative studies on the biology of dryinid wasps in Japan. 5) Development and reproductive capacity of hosts attacked by Haplogonatopus apicalis (Hymenoptera, Dryinidae) and the development of progenies of parasites in their hosts. Kontyû. 56: 659-666 [ Links ]
Johanowicz, D.L & M.A. Hoy. 1998. The manipulation of arthropod reproduction by Wolbachia endosimbionts. Fla. Entomol. 81: 310-317. [ Links ]
Mattenson, P.C. 2000. Insect pest management in tropical Asian irrigated rice. Annu. Rev. Entomol. 45: 549-574. [ Links ]
Mattenson, P.C., K.D. Gallagher & P.E. Kenmore. 1994. Extension of integrated pest management for planthoppers in Asian irrigated rice: Empowering the user, p. 656-685. In R.F. Denno and C.J. Perfect. (eds). Planthoppers, their ecology and management. Chapman and Hall, Col., New York, USA. [ Links ]
Meneses, R., A. Gutiérrez, G. Antigua, J. Gómez & F. Correa. 1998. Guía para el Manejo Integrado de Plagas del Arroz. IIA-CIAT-FLAR. [ Links ]
Mora, R., A. Retana & A.M. Espinoza. 2001. External morphology of Tagosodes orizicolus (Homoptera: Delphacidae) revealed by scanning electron microscopy. Ann. Entomol. Soc. Am. 94: 438-448. [ Links ]
Mora R., J.E. Ibarra & A.M. Espinoza. 2007. A reliable bioassay procedure to evaluate per os toxicity of Bacillus thurigiensis strains against the rice delphacid Tagosodes orizicolus Muir (Homoptera: Delphacidae). Rev. Biol. Trop. 55: 373-383. [ Links ]
Morales, F.J. & A. Niessen. 1985. Rice hoja blanca virus. Description of plant viruses. No. 299. Commonwealth Mycological Institute/ Association of Applied Biologists, Kew Surrey, England. [ Links ]
Ode, P.J. & K.M. Heinz. 2002. Host-size-dependent sex ratio theory and improving mass-reared parasitoid sex ratios. Biol. Control. 24: 31-41. [ Links ]
Olmi, M. 1984. A Revision of the Dryinidae (Hymenoptera). Memoirs of the American Entomological Institute. 37: 1-1913. [ Links ]
Olmi , M. 1995. Dryinidae, p. 493-503. In P.E. Hanson & I.D. Gauld (eds.). The Hymenoptera of Costa Rica. Oxford University, Oxford, England. [ Links ]
Peñaranda, V.H., L.A. Reyes, R. Perafán, I. Devia, & P. Hernández. 1999. Manejo integrado de sogata (Tagosodes orizicolus) Muir en el cultivo de arroz en los Llanos orientales. Federación de Arroceros de Colombia, Bogotá, Colombia. [ Links ]
Rey, X. & A. García. 1980. Estudio de algunos aspectos del ciclo de vida del Sogatodes orizicola. Cienc. Tec. Agric. Arroz 3: 31-65. [ Links ]
Sahragard, A., M.A. Jervis & N.A.C. Kidd. 1991. Influence of host availability on rates of oviposition and host feeding, and on longevitiy in Dicondylus indianus Olmi (Hym., Dryinidae), a parasitoid of the rice brown Planthopper Nilaparvata lugens Stål ) Hem, Delphacidae). J. Apl. Ent. 112: 153-162. [ Links ]
Suomalaine, E. 1962. Significance of parthenogenesis in the evolution of insects. Annu. Rev. of Entomology. 7: 449-366. [ Links ]
Virla, E. & M. Olmi. 1998. The Dryinidae of Argentina. Acta Entomol. Chilena 22: 19-35. [ Links ]
Waloff, N.1974. Biology and behavior of some species of Dryinidae (Hymenoptera). J. Entomol. (A) 49: 97-109. [ Links ]
Zar, J. H. 1996. Biostatistical Analysis. Prentice Hall, New Jersey, USA. [ Links ]

