










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 suppl.1 San José Nov. 2009
Growth dynamics and state of the seagrass Thalassia testudinum in the Gandoca-Manzanillo National Wildlife Refuge, Caribbean, Costa Rica
Lucia S. Krupp1*, Jorge Cortés2, 3 & Matthias Wolff1, 4
1. Leibniz-Center for Tropical Marine Ecology (ZMT), Fahrenheitstr. 6, 28359 Bremen, Germany; lucia.krupp@zmt-bremen.de; matthias.wolff@zmt-bremen.de
2. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Universidad de Costa Rica, San Pedro, 115012060 San José, Costa Rica; jorge.cortes@ucr.ac.cr
3. Escuela de Biología, Universidad de Costa Rica, San Pedro, 11501-2060, San José, Costa Rica.
4. Charles Darwin Foundation, Puerto Ayora, Santa Cruz, Galapagos, Ecuador. Casilla 17-1-3891, Quito, Ecuador (593-5) 2526146 or 2526147 (extension 123), cellular (593) 097357343, skype matthias.wolff, matthias.wolff@fcdarwin.org.ec
* Corresponding author. Tel.: ++49 (0)421 23800 79; fax: ++49 (0)421 23800 40
Abstract: Seagrass beds are highly productive and valuable habitats, which fulfill a key role in coastal ecosystems. Spatial distribution, biomass, density, productivity and leaf dynamics of the dominant seagrass species Thalassia testudinum were studied at five locations in the Gandoca-Manzanillo National Wildlife Refuge, Caribbean coast of Costa Rica, from November 2005 until March 2006. Seagrass beds within the study area cover approximately 16 ha. Spatial and temporal differences in ecological seagrass parameters were examined along gradients of riverine inputs and related to a range of environmental parameters (depth, grain size distribution, nutrient concentrations, salinity and temperature). Average canopy cover and above-ground biomass of T. testudinum inversely correlated with depth, but did not vary significantly between study sites when compared along depth ranges. Considerable spatio-temporal variations in shoot densities, areal productivity and leaf sizes seem to be related to riverine inputs and wave energy. It appeared that T. testudinum at exposed sites respond to increased environmental disturbance related to the offset of the rainy season with clonal recruitment, whereby shoot densities increase and average leaf sizes are reduced. Lower shoot densities and greater leaf sizes, in contrast, are characteristic for locations with rather consistent environmental conditions, where seagrasses are sheltered. T. testudinum in the refuge has higher shoot densities but shorter leaves and lower productivity compared to sites in 15 other Caribbean countries. The seagrass beds appear to be in a relatively healthy state, however, observations of temporal increased stocks of filamentous epiphytes and macroalgae indicate temporal environmental stress in the system. Rev. Biol. Trop. 57 (Suppl. 1): 187-201. Epub 2009 November 30.
Key words: seagrass, Thalassia testudinum, Caribbean, CARICOMP, spatio-temporal dynamics, biomass, density, productivity, Costa Rica.
Seagrass beds are highly productive and valuable habitats, which fulfill a key role in coastal ecosystems (Duarte 2002). Despite an increasing understanding and recognition of their ecological and economic importance, seagrass declines are reported worldwide (e.g. Robblee et al. 1991, Preen et al. 1995, Baden et al. 2002). The main reason for that decline is an increased pressure on the coastal zones caused by human population expansion and tourism. Nutrients over-enrichment and excessive sediment runoff due to increased soil erosions are cited most often as the main threats, besides many other direct or indirect anthro pogenic effects (Short & Wyllie-Echeverria 1996, Waycott et al. 2009). Rivers are the most important pathway by which sediments and nutrients are transported into the ocean (Mil liman & Meade 1983). Thereby, they can cause siltation of coastal ecosystems resulting in increased water turbidity as well as higher silt, nutrient and organic matter contents in the sediment (Terrados et al. 1998; Kamp-Nielsen et al. 2002). Seagrasses are negatively affected by siltation indicated by decreasing species abundance, shoot density and leaf biomass with increasing siltation pressure (Bach et al. 1998, Terrados et al. 1998). Nutrients delivered by rivers can stimulate growth of macroalgae growing on the seafloor and on seagrass leaves, which can shade and eventually out-compete seagrasses as the dominant producer (valiela et al. 1997, McGlathery 2001). This generally suggests sub-optimal conditions for seagrass development in the vicinity of rivers (Terrados et al. 2008).
Numbers of studies focusing on seagrasses have increased during the past decades, but knowledge on tropical seagrasses is still underrepresented. Information on the ecology and conservation status of seagrasses in Central America is very limited (Cortés 2007). Only three studies on seagrasses at the Caribbean coast of Costa Rica have been published. Paynter et al. (2001) studied the basic ecology of T. testudinum seagrass beds at three sites in Cahuita National Park (9°45N, 82°48W) and found higher biomass and productivity to result from a combination of intermediate sediment size and moderate water temperature and salinity. The second paper was published by Fonseca et al. (2007) and they found in Cahuita National Park, over a six year period, that productivity and total biomass were intermediate, while turnover rates and shoot densities were higher compared to other sites in the Caribbean. The last paper was Published by Nielsen-Muñoz & Cortés (2008), they analyzed samples from Cahuita and Manzanillo. They found that average biomass of Thalassia testudinum within the substrate was more than double the biomass above ground, productivity was highest in July, and flowering was between April and June, with a peak in May. In this paper we present the results of an investigation conducted in the Gandoca-Manzanillo Nacional Wildlife Refuge (GMNWR), Caribbean coast of Costa Rica, approximately 30km Southeast of Cahuita. The occurrence of seagrass in the area was mentioned by Cortés & Guzmán (1985), Cortés (1992) and Fernández & Alvarado (2004), but a detailed assessment was as yet lacking.
Herein, we describe the distribution of seagrasses in the GMNWR and examined spatial and temporal variations in biomass, productivity, shoot density and leaf parameters of T. testudinum during the rainy season from November 2005 to March 2006. The respective parameters were used to determine the overall performance of seagrasses aiming at statements about the health of the seagrass beds in the area. It was hypothesized that seagrasses influenced by river inputs show a lower performance compared to non-impacted sites. The results are compared to data sets from CARICOMP (Caribbean Coastal Marine Productivity), the Caribbean-wide monitoring network (CARICOMP 1997).
Materials and methods
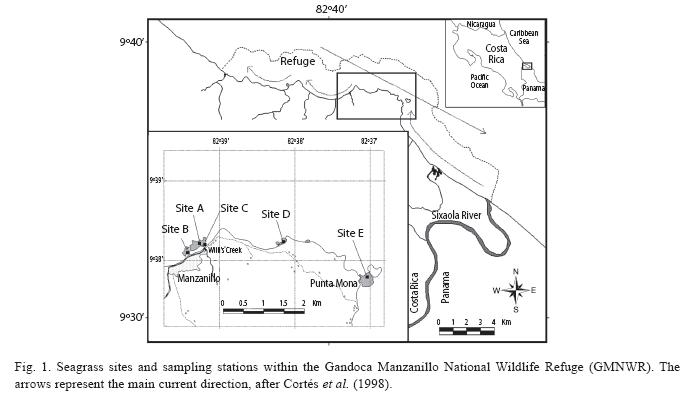
Study area: The Gandoca-Manzanillo National Wildlife Refuge (9°38N, 82°39W) is situated at the Southern Caribbean coast of Costa Rica and comprises 5 013ha of land and 4 436ha sea. The region is characterized by a tropical, hot and humid climate with intense rains (2 500-3 000mm per year) mainly during two rainy seasons (Nov-Mar and Jun-Aug). Local winds and tropical storms regularly affect the coast. Air temperatures range from 24°C to 27°C (Herrera 1984). The direction of the main marine current is from Northwest to Southeast (Cortés et al. 1998) forming eddies in the opposite direction (Fig. 1). Tides are mixed, mainly diurnal and show a range of 30 to 50cm (Cortés 1998). Due to the protection status as a wildlife refuge and relatively low degree of urban and industrial development in the vicinity, the anthropogenic pressure on the ecosystems of the GMNWR can be estimated to be modest. Seagrasses grow in reef lagoons with a water depth between 1 and 5 meters. The turtle grass, Thalassia testudinum Banks ex König, is the dominant species, occurring locally intermixed with the manatee grass, Syringodium filiforme Kützing, 1860.
Seagrass mapping and study sites: The main continuous seagrass patches between the villages of Manzanillo and Punta Mona (Fig. 1) were identified and their borders were recorded with a GPS device. Seagrass beds were mapped based on these GPS points and tracks and their spatial extent was measured using the software Arc view 3.3. A topographic map (IGN, Hoja 3644 Sixaola, 1:50000) was used as a reference to illustrate geographical and man-made features.
Five study sites exposed to apparently different environmental conditions were chosen and a particular sampling station was set up within each site (Fig. 1). Sites A, B and C were situated in the same bay close to the waterfront of Manzanillo Village, where seagrasses are often exposed to wave energy. Site C (9°3811.1N; 82°3911.8W) was directly in front of the mouth of Willis Creek, which drains substantial amounts of freshwater into the sea after heavy rains. The water of the creek has a dark brown colour and appears to contain great amounts of organic matter. After storm events the plume is deflected Northward and also affects sites A (9°3812.1N; 82°3914.3W) and B (9°3805.3N; 82°3925.0W). Site B was close to the shore imbedded in a nearshore fringing reef, whereas site A was situated further offshore and parts of the site had a greater depth. Site D (09°3813.5N; 83°3809.5W) was located in a small bay surrounded by rain forest Southeast of Manzanillo. It was chosen as the control site, as human activity is minimal, there are no rivers nearby and the site is sheltered from waves and currents by a coral reef. Site E (09°3746.3N; 82°3706.0W), near the spit of Punta Mona, was expected to be influenced by freshwater, sediment and nutrient loads from the plume of the Sixaola River, which is transported from the river mouth (approximately 9km away) by a current (Cortés et al., 1998; Fig. 1).
Structure of the seagrass beds: Fifty-four parallel transects perpendicular to the shore and spaced approximately 15 meters apart were surveyed within the designated seagrass beds (n=depending on site length). A quadrat (0.25 m²) was placed on 10 pre-determined random positions along the 50m transect tape. The cover of T. testudinum and S. filiforme was assigned in each quadrat using a rapid visual assessment technique based on the Braun-Blanquet abundance score (Braun-Blanquet 1972, Fourqurean et al. 2001) ranging from 0 (species absent) to 5 (cover >75%). The average seagrass cover (C) at each site was computed using the following statistical equation:
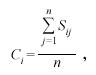
where Ci = cover of species i, j = quadrat number from 1 to n sampled at a site, Sij = the Braun-Blanquet score for species i in quadrat j.
Additionally, the percentage cover of live coral colonies, dead coral rock and macroalgae, as well as numbers of sea urchins were estimated for each quadrat and calculated according to English et al. (1994).
Biomass, shoot and leaf dynamics: For biomass determination, 10 core samples, 30 to 45cm deep and 16cm in diameter, were taken randomly from each study site using a hand corer. The approximate water depth was recorded for each sample. Plant parts were cleaned from sediments (retaining 500g wet weight for grain size distribution analysis) and frozen in plastic bags until further analysis. In the laboratory T. testudinum plants were separated into green leaves, non-green leaves and short shoots (SS), live rhizomes, live roots and dead below ground material. Leaf epiphytes were removed by rinsing green leaves in 10% hydrochloric acid. Fractions were dried on aluminum foil tares at 75°C and weighed for dry weight (DW) determination. Total-, leaf-, above- and below-ground biomasses (DW/m2) were calculated for each core sample. Above-ground biomass consists of green leaves and short shoots, whereas leaf biomass only comprises photosynthetic parts. The samples were grouped into depth ranges (0-1m, 1-2m, 2-3m, 3-4m), to visualize the bathymetric distribution of biomass.
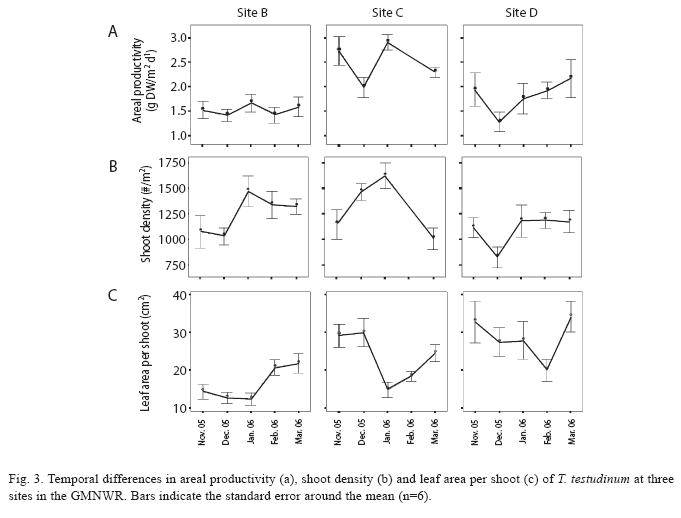
Net above-ground productivity and shoot densities of T. testudinum were determined monthly (Nov. 2005 to Mar. 2006) at the sampling stations of sites B, C and D using a leaf marking technique adapted from Zieman (1974) and Zieman et al. (1999). Shoots within six tagged wire quadrats (10x20cm) were marked by simultaneously piercing all leaves of a shoot at the leaf base with a single punch of a needle. After a growth period of 9 to 14 days, the shoots were harvested and counted, in order to estimate shoot densities. Leaves from each quadrat were sorted into three fractions: (1) New leaves (unmarked leaves that had emerged after marking), (2) Old growth (the part of the leaves between the needle mark and meristem (=green-white interface), (3) Old standing crop (the leaf part above the needle mark). Leaf fractions were decalcified in 10% hydrochloric acid, rinsed and dried at 75°C to constant weight for dry weight (DW) determination. values for areal productivity (P), turnover rate (T) and shoot density (D) were calculated for each replicate. The following equations were applied to compute P and T:
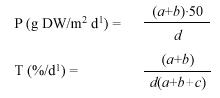
where a=DW of new leaves, b=DW of old growth, c=DW of old standing crop and d is the number of days. Sample fractions from November 2005 were additionally measured (length and width) and the daily leaf elongation was calculated. There is no data available for site C in February because all samples (quadrats) had disappeared during a storm.
Another 10 shoots were collected randomly from each sampling station every month. Leaf blades were counted, and total length and width of each leaf measured. The leaf area per shoot and the leaf area index (LAI), which is the product of leaf area per shoot and shoot density, were calculated.
Environmental parameters: Water temperature and salinity at the sampling stations were measured on a weekly basis using a mercury thermometer and a hand refractrometer respectively. Measurements of water in 50cm
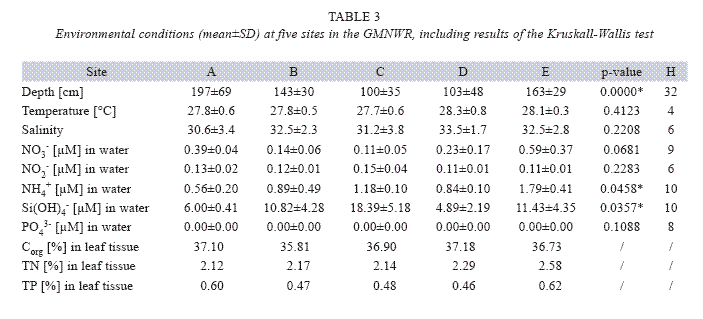
depth were conducted, whereby the mean of three subsequent measurements was recorded for each parameter. For nutrient analysis, three 250ml samples of surface water were randomly collected from each study site in March 2006 at intermediate depth, filtered, treated with HgCl2 and frozen until processing within the following month. values of nitrate (NO3-), nitrite (NO2-), ammonium (NH4+), phosphate (PO43-) and silicate (Si(OH)4-) were determined spectrophotometrically by a SAN plus automated continuous flow analyzer according to methods of Grasshoff et al. (1983). T. testudinum shoots (n=10) collected from each site in March 2006 were analyzed for contents of organic carbon (Corg), total nitrogen (TN) and total phosphorus (TP) as indicator for long-time exposure to nutrients of the surrounding seawater. Leaf tissue samples were dried and ground after rinsing and removing the epiphytes with a knife. Corg and TN values were determined with a NA 2100 elemental analyzer. TP contents were assessed with a Shimadzu Pharma Spec Uv-1700 Uv-visible spectrophotometer after muffling at 810°C and dissolving in 21.5% HNO3 (method by Scheffer & Pajenkamp 1952). Sediments collected with the sample cores for seagrass biomass analysis (n=10) were analyzed for grain size distribution. The sub-samples were dried at 100°C for 48 hours. A mechanical granulometric analysis using wet sieving was conducted with mesh sizes of a diameter between 0.075 and 25.4mm. Percentages of gravel (>4.75mm), course (1.254.75mm), intermediate (0.25-1.25mm) and fine (0.075-0.25mm) sand and silt+clay (<0.075mm) were calculated for each replicate. visual information on the appearance of seagrass health were collected at each site visit. Thereby, changes in epiphyte and macroalgae abundance, as well as other unusual observations were recorded.
Data analysis: The software SPSS13 and Statistica6 were used to perform the statistical data exploration of seagrass and environmental parameters. Site and monthly means, and standard deviations were calculated for all variables. Since the data of the majority of parameters was not normally distributed (also after various transformations), the non-parametric Kruskal-Wallis test was used to examine whether estimated parameters differed in sites or in months. If the means were significantly different (p<0.05) the non-parametric MannWhitney-U two sample test was applied, in order to compare each parameter between all sites and all months. The non-parametric Spearman test was used to detect, whether there were significant correlations (p<0.05) between seagrass and environmental parameters.
Results
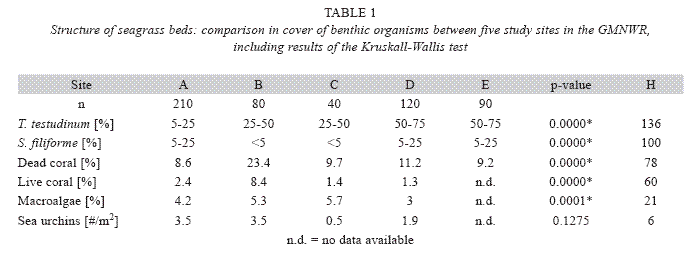
Seagrass beds in the study area cover a total area of 16ha (Fig. 1). Seagrass was encountered at 93% of the 540 evaluated random quadrats, whereby T. testudinum was found in 89% and S. filiforme in 73% of all quadrats. The average cover of T. testudinum was highest at sites D and E (50-75%) and lowest at site A (5-25%) (Table 1). There was a highly significant correlation of T. testudinum and S. filiforme cover and depth (Spearman, p<0.05). The percentage cover of bio other benthic organisms, such as live and dead corals, macroalgae and sea urchins, was significantly higher at site B (Mann-Whitney, p<0.05) significompared to most other sites (Table 1).
The average total biomass of T. testudinum at the study area was 1564±749g DW/m2 with an average above/below-ground ratio of 0.84. Total-, leaf-, above- and below-ground biomasses of T. testudinum varied significantly between sites (p<0.05), with the highest means the at site C and lowest at site B (Table 3). All
biomass fractions inversely correlated with depth (Spearman, p=0.00). Above-ground mass was fairly similar at all sites, when compared along the same depth range (p>0.05), however below-ground biomass was cantly higher at site C compared to site D at depths of 0-1m (p<0.05) and at site A compared to all other sites at a depth of 1-2m (p<0.05) (Fig. 2).
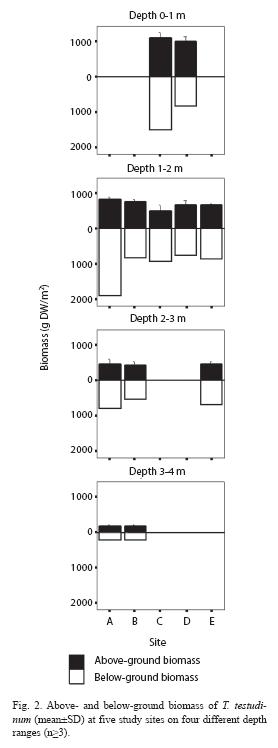
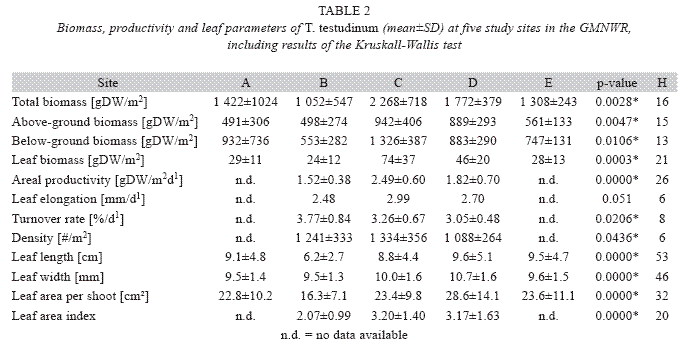
All productivity and leaf parameters varied significantly between the study sites (Kruskall Wallis, p<0.05) (Table 2). Even though deeper site B had the lowest mean areal productivity and daily leaf elongation, it had the greatest average turnover rate (3.8 %/d1) compared to sites C and D (Table 2). Site C, in turn, had the highest mean areal productivity (2.5g DW/m2d1), daily leaf elongation (3.0mm/ d1) and shoot density (1334/m2). Shoots had between 1 (Min) and 5 (Max) leaf blades (Mean: 2.6), which measured on average 8.6cm in length and 9.9mm in width. Leaves were on average significantly shorter at site B and significantly wider at site D (Mann-Whitney, p<0.05). The average leaf area per shoot and average leaf area index at sites C and D were fairly similar (Mann-Whitney, p>0.05), means at site B however were significantly smaller compared to the other two sites (Mann-Whitney, p<0.05). Productivity and leaf parameters inversely correlated with depth (Spearman, p<0.05), except turnover rate, which correlated positively, and leaf length and width, which did not correlate with depth (p>0.05).
Considerable monthly variations in productivity, shoot densities and leaf area per shoot were observed (Fig. 3): In December, the areal productivity was lowest at all study sites, whereas in January it was significantly higher at site C and also slightly higher at site B (Fig. 3a). Also shoot densities were significantly higher at site B and C in January (Mann-Whitney, p<0.05) and slightly lower again in February and March, while such a shift was not observed at site D (Fig. 3b). With a decrease towards January followed by an increase in February, leaf area per shoot showed an opposite trend compared to areal productivity and shoot density at sites B and C (Fig. 3).
Ammonium and silicate concentrations were the only environmental variables that varied significantly between the study sites (Table 3). Sites C and D were shallowest with approximately 1m average depth, sites B and E with an approximate mean of 1.5m were of intermediate depth and the largest site A was deepest with about 2m average depth (Table 3). Sea water along the study area had an average temperature of 27.9°C (±0.6°C) and salinity of 31.9 (±2.8). Both parameters showed strong variability within each month at all sites, however, monthly averages did not vary significantly between the study sites and did not correlate with seagrass parameters. Nutrient concentrations in the surface water were overall highest at the river-near sites C and E and lowest at site D. Concentrations of nitrate, ammonium and phosphate were significantly higher at site E compared to most other sites; however phosphate concentrations were probably at or below the detection level. N and P contents in T. testudinum leaf tissue were also higher at site E compared to all other sites (Table 3).
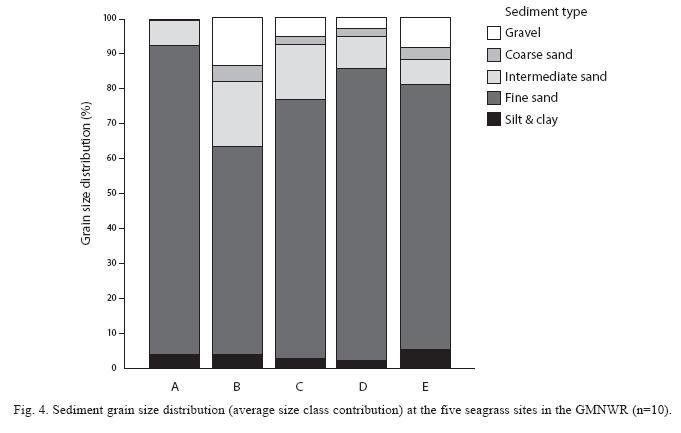
Sediments were characterized by relatively high amounts of fine sand (Fig. 4), however sites differed significantly in all grain size types (Kruskall-Wallis, p<0.05). The substrate was overall finest at site A, whereas the proportion
of coarse sediment types was greatest at site B. Relative contents of silt and clay were highest at site E and lowest at site D, however values were overall fairly low (mean of 4% ±2.5%). There were no significant correlations between sediment size composition and biomass parameters (Spearman, p<0.05).
A thick coating of filamentous leaf epiphytes, instead of the usually occurring calcareous red algae, colonized both seagrass species in early December 2005. This was most obvious at site C. One month later, the typically occurring epiphyte communities were found again. At the end of March 2006, it was observed that the seagrass sites were intermixed with substantial amounts of macroalgae. This was recorded to be particularly high at site E, where macroalgae had about twice the canopy height of the seagrass and seemed to be dominating the seagrass bed. Algae taxa observed in unusually high abundances were mainly Padina jamaicensis in shallow areas and a filamentous algal species (possibly Chaetomorpha linum) in deeper areas. The occurrence of macroalgae could not be studied in detail and leaves space for further investigation.
Discussion
Growth dynamics: The results of a comparison in canopy cover, biomass, productivity and leaf parameters of T. testudinum revealed that there are several spatial and spatio-temporal variations of T. testudinum performance within the study area. Since most seagrass parameters inversely correlate with depth, a variation in depth profiles can be evaluated as one of the most crucial factors for differences in seagrass performance between the study sites. The shallow sites C and D showed the overall best performance with the greatest average canopy cover, biomass and productivity. This is not surprising, since several studies have demonstrated that those parameters are usually higher in shallow waters as an effect of increased light irradiance (e.g., Short & Burdick 1995, Dawes 1998). While above-ground biomass and canopy cover were fairly similar between the study sites along the same depth range, significant differences were given in shoot sizes, turnover rates, productivity and shoot densities. It thus appears that seagrasses either occur in high shoot densities with smaller leafs and show great temporal variability (sites C and B), or they invest energy in greater leaf sizes and have lower shoot densities (site D).
A significant increase in shoot densities and productivity at the river near sites C and B in December and January respectively can probably be related to clonal recruitment of new seagrass shoots. Since new shoots are smaller than older shoots, average leaf sizes were lowered. Several studies discuss density-dependent regulations as a mechanism to avoid self-shading, whereby shoot densities of sea-grass populations increase with decreasing shoot sizes and vice versa (Duarte & Kalff 1987, Olesen & Sand-Jensen 1994, Hemminga & Duarte 2000). This mechanism could also explain an opposite trend observed at sites B and C in February and March: recruits grew up resulting in greater leaf sizes, while older shoots were lost.
Since variations in seagrass characteristics could neither be correlated to differences in sediment grain size, nor to any other of the environmental parameter measured, the observed spatial and temporal differences in seagrass parameters should be attributed to the varying influences of river inputs and wave strength at the study sites. The increased vegetative reproduction of short shoots at the river-exposed sites was probably a response to increased environmental stress caused by changing weather conditions. Since the study started at the onset of the rainy season, it can be expected that the first significant storm and rain events took place in November and that they became more frequent and enduring towards December and January, the peak of the rainy season. Thereby, increased wave strength and river drainage might have led to shoot declines needing the recruitment of new shoots to compensate for losses of leaf material. Hemminga & Duarte (2000) also stated that enhanced shoot recruitment might be a morphological and physiological adaptation to moderate disturbance, whereby losses involve a thinning of a seagrass bed allowing recovery through clonal growth from the surrounding rhizome apices. Such a response to disturbance might have occurred earlier at the river near site C (December) and slightly later (January) at site B with increasing amounts of rain. At site D, to the contrary, where T. testudinum appears fairly undisturbed, available energy might have been invested in leaf size maximizations rather than recruitment.
Several effects caused by river inputs and wave strength can be discussed. On the one hand, it could be argued that increased freshwater could have changed salinity and temperature conditions, but all respective measurements of both variables were within the optimum range of T. testudinum growth (Phillips 1960, Moore 1963) and mean values were rather similar at all sites. Thus, it seems more probable that reduced light levels caused by increasing loads of particulate matter in river water may have been responsible for changes in seagrass growth dynamics. In addition, physical stress caused by sedimentation was dramatically illustrated in February, when an entire part of site C disappeared due to burial by sediments. At the more exposed sites A, B and C increasing and enduring wave strength triggered by frequent storm events might have additionally caused mechanical uprooting of seagrass leaves. A higher exposure to wave strength could also explain the higher below-ground biomass at site C compared to site D. Here, a higher root-rhizome system compared to the other exposed sites might also be necessary because of the sites shallowness. This is supported by the fact that at greater depth, where wave energy is generally weaker, below-ground biomasses are almost the same at all sites.
Another hypothesis is that clonal recruitment occurred as a response to environmental stress caused by shading of filamentous epiphytes, observed on seagrass blades at site C in the beginning of December 2005. Most likely, a temporal nutrient over-enrichment after strong rains and subsequent dry and calm weather and sea conditions may have triggered an accelerated growth of these epiphytes. A shift from calcareous to filamentous epiphytes in response to elevated water-column nutrients was observed in several field studies and fertilization experiments (e.g. Tomasko & Lapointe 1991, Necklers et al. 1993, McGlathery 1995, Short & Burdick 1995). Amplified stocks of macroalgae recorded in March 2006, especially at site E, can also be related to increased nutrient levels, as reported before (e.g., Short & Burdick 1995, valiela et al. 1997, McGlathery 2001). Unsuccessful competition with macroalgae in eutrophic waters is even considered to be one of the primary reasons for seagrass declines world-wide (Short & Wyllie-Echeverría 1996, Waycott et al. 2009).
The chemical analysis of surface water confirmed higher levels of ammonium, nitrate and phosphate at site E and slightly increased values also at site C. However, water column concentrations of limiting nutrients generally appear as poor indicators for the trophic state of marine systems (Smith et al. 1981, valiela et al. 1990), due to small-scale current regimes and/or a rapid uptake and turnover (Suttle & Harrison 1988, Suttle et al. 1990, Tomasko & Lapointe 1991). Elevated values of TN and TP in T. testudinum leaf tissue, compared to data cited in the literature (e.g. Duarte 1990, Fourqurean et al. 1992, Terrados et al. 2008), however, confirm trends observed by the analysis of water nutrients. A positive relationship between nutrient contents of seagrass leaf tissue and surface water was also frequently observed (e.g. McGlathery 1995, Jones et al. 2001). Seagrasses at site E might be exposed to higher long term nutrient concentrations originating from the Sixaola River, which might be enriched with fertilizers used at the several banana plantations along the riverbanks. After strong rain, these fertilizers may be washed into the river and be transported to site E. Even modest alterations in nitrogen loadings from watersheds bare the potential for a regime shift from a seagrass to a macroalgae dominated ecosystem (valiela et al. 1997). A significantly poorer visibility observed at this site indicates light reduction, possibly due to sedimentation resulting from increasing erosion in the hinterland of the Sixaola River. Siltation stress in the area, also reported by Cortés & Risk (1985) and Cortés et al. (1992), additionally causes light stress to the seagrasses and amplifies the physiological advantages of macroalgae over seagrass growth.
Concerning observed macroalgae and epiphytes it could be argued that seagrass sites influenced by river inputs indeed show a lower performance, as it was hypothesised. However, this could not be confirmed by seagrass parameters measured, especially if sites C and D are compared. Firstly, a higher average daily leaf elongation at site C when compared with site D confirmed that values of areal productivity are not exclusively enhanced by higher shoot densities, but that growth rates are generally greater at this site. Since both sites are equal in depth, higher growth rates can most probably be attributed to a greater availability of nutrients derived by the river, as reported by Agawin et al. (1996). It appears that nutrients cause stress to seagrasses in form of competition by epiphytes and macroalgae only when a certain threshold of nutrient concentrations is exceeded. Secondly, even though there are significant differences in leaf sizes and shoot densities, the leaf area index, which combines both variables, is about the same at both sites. This important ecological variable, which is also often linked to abundance of associated macrofauna, suggests that none of the growth concepts, leaf size maximization or clonal recruitment, can be regarded as preferential.
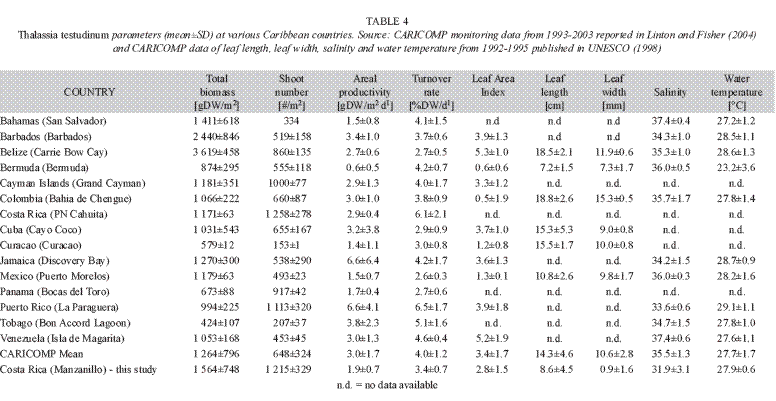
Comparison with other Caribbean sea-grass beds: An overall evaluation of ecological parameters shows that the performance of T. testudinum in the GMNWR is comparable with other countries in the Caribbean region (Table 4). In contrast to most other seagrass areas in the Caribbean, shoots in higher densities but with relatively shorter leaf lengths and lower areal productivity seem to be characteristic for T. testudinum in the GMNWR. The canopy height (leaf length) of T. testudinum at the GMNWR appears about half as high as that in Belize and Colombia, which also explains why the leaf area index and biomass of T. testudinum is greater in these countries. As CARICOMP (1997) assumes, seagrasses at continental areas and large islands allow the development of larger plants with wider leaves and denser shoots because of a higher nutrient availability. Seagrasses at the GMNWR only fulfil this theory to a limited extent. Possibly, the concept of a morphological adaptation of T. testudinum to higher disturbance by exposure to waves, currents and river inputs, resulting in greater leaf recruitment but shorter leaf sizes, as mentioned above, generally holds for the seagrasses in the GMNWR. Lower salinity values compared to all other CARICOMP sites confirm that seagrasses in the GMNWR indeed are more influenced by river systems.
Health state of the seagrass beds: Since biomass and shoot density can be regarded as important health indicators of a seagrass system, the overall state of T. testudinum in the GMNWR seems good. Most probably, seagrasses in the GMNWR benefit from their protective status as a Wildlife Refuge and the up to now moderate development of the area. Concerning the quantitative analysis of ecological parameters of T. testudinum, the overall state of the seagrass beds can be evaluated as relatively healthy. However, observations of elevated amounts of macroalgae and filamentous epiphytes give reason to concern that the seagrasses are temporarily exposed to environmental stress, which the measurement of seagrass parameters failed to detect. Nutrient over-enrichment and sedimentation seem to be the main reasons. Seagrasses at the river impacted sites C and E appear especially sensitive to a potential future development of the coastal region. Bearing these facts in mind, it could be speculated that the seagrass system at the GMNWR is at the edge of its high performance level. The seagrasses have shown a high resilience so far, but if nutrient levels further increase, the persistence of the seagrass ecosystem will be endangered.
Acknowledgments
The authors thank the Ministerio del Ambiente y Energía (MINAE), Costa Rica, for their co-operation and first of all José Saballo for his valuable assistance in the field. We thank Shawn and Gregory Larkin from the dive base AQUAMORE in Manzanillo for logistical support and Matthias Birkicht and Javier Alvarado for assistance conducting laboratory analyses. Lucia Krupp was supported by a scholarship grant of the German Academic Exchange Service (DAAD).
Resumen
Las praderas de pastos marinos son sitios altamente productivos y hábitats valiosos en los ecosistemas costeros. Se estudió los cambios espaciales y temporales en la distribución del pasto marino dominante Thalassia testudinum, en cinco localidades expuestas a un gradiente de influencia fluvial, en el Refugio Nacional de vida Silvestre Gandoca-Manzanillo, Costa Rica, entre noviembre 2005 y marzo 2006. El pasto cubre unas 16ha. Tanto cobertura como biomasa disminuyen al aumentar la profundidad, pero no hubo diferencia de cobertura y biomasa entre lugares. Las variaciones espacio-temporales en densidad de estolones, productividad y tamaño de hojas, probablemente están relacionadas con la entrada de agua del río y la energía de las olas. Al parecer T. testudinum responde a disturbios ambientales con reclutamiento clonal: la densidad de estolones aumenta y las hojas son más pequeñas. En Gandoca-Manzanillo la densidad de estolones es mayor, las hojas más cortas y la productividad inferior, en comparación con otros 15 sitios del Caribe. Según estos parámetros, los pastos marinos de Gandoca-Manzanillo están en muy buenas condiciones. Sin embargo, se observaron aumentos en algas filamentosas epífitas y de macroalgas, que indican un impacto temporal en el sistema.
Key words: pastos marinos, Thalassia testudinum, Caribe, CARICOMP, dinámica espacio-temporal, biomasa, densidad, productividad, Costa Rica.
Received 02-V-2009. Corrected 15-VII-2009. Accepted 02-IX-2009.
References
Agawin, N.S.R., C.M. Duarte & M.D. Fortes. 1996. Nutrient limitation of Philippine seagrasses (Cape Bolinao, NW Philippines): in situ experimental evidence. Mar. Ecol. Prog. Ser. 138: 233-243. [ Links ]
Bach, S.S., J. Borum, M.D. Fortes & C.M. Duarte. 1998. Species composition and plant performance of mixed seagrass beds along a siltation gradient at Cape Bolinao, The Philippines. Mar. Ecol. Prog. Ser. 174: 247-256. [ Links ]
Baden, S., M. Gullström, B. Lundén, L. Pihl & R. Rosenberg. 2002. vanishing seagrass (Zostera marina, L.) in Swedish coastal waters. Ambio 19: 113-122. [ Links ]
Braun-Blanquet, J. 1972. Plant sociology: The study of plant communities. Hafner, New York. [ Links ]
CARICOMP. 1997. variation in ecological parameters of Thalassia testudinum across the CARICOMP network. Proc. 8th Int. Coral Reef Symp., Panama 1: 663-668. [ Links ]
Cortés, J. 1992. Los arrecifes coralinos del Refugio Nacional de vida Silvestre Gandoca-Manzanillo, Limón, Costa Rica. Rev. Biol. Trop. 40: 325-333. [ Links ]
Cortés, J. 1998. Cahuita and Laguna Gandoca, Costa Rica: 107-113. In: B. Kjerfve (ed.), CARICOMP-Caribbean coral reef, seagrass and mangrove sites. UNESCO, Paris. [ Links ]
Cortés, J. 2007. Coastal morphology and coral reefs: 185-200. In: J. Bundschuh & G.E. Alvarado (Eds.). Central America: Geology, resources and hazards, volume 1. Taylor & Francis, London. [ Links ]
Cortés, J. & H.M. Guzmán. 1985. Arrecifes coralinos de la costa Atlántica de Costa Rica. Brenesia 23: 275-292. [ Links ]
Cortés, J. & M.J. Risk. 1985. A reef under siltation stress: Cahuita, Costa Rica. Bull. Mar. Sci. 58: 339-356. [ Links ]
Cortés, J., R. Soto, C. Jiménez & A. Astorga. 1992. Earthquake associated mortality of intertidal and coral reef organisms (Caribbean of Costa Rica). Proc. 7th Int. Coral Reef Symp., Guam 1: 235-240. [ Links ]
Cortés, J., A.C. Fonseca, M. Barrantes & P. Denyer. 1998. Type, distribution and origin of sediments of the Gandoca-Manzanillo National Wildlife Refuge, Limón, Costa Rica. Rev. Biol. Trop. 46 (Suppl. 6): 251-256. [ Links ]
Dawes, C. 1998. Biomass and photosynthetic responses to irradiance by a shallow and a deep water population of Thalassia testudinum on the west coast of Florida. Bull. Mar. Sci. 62: 89-96. [ Links ]
Duarte, C.M. 1990. Seagrass nutrient content. Mar. Ecol. Prog. Ser. 6: 201-207. [ Links ]
Duarte, C.M. 2002. The future of seagrass meadows. Environ. Conserv. 29: 192-206. [ Links ]
Duarte, C.M. & J. Kalff. 1987. Weight-density relationships in submerged macrophytes: the importance of light and plant geometry. Oecologia 72: 612-617. [ Links ]
English, S., C. Wilkinson & B. Baker (eds.). 1994. Survey manual for tropical marine resources. Aust. Inst. Mar. Sci., Townsville, Australia. [ Links ]
Fernández, C. & J.J. Alvarado. 2004. El arrecife coralino de Punta Cocles, costa Caribe de Costa Rica. Rev. Biol. Trop. 52 (Suppl. 2): 121-129. [ Links ]
Fonseca, A.C., v. Nielsen & J. Cortés. 2007. Monitoreo de pastos marinos en Perezoso, sitio CARICOMP en Cahuita, Costa Rica. Rev. Biol. Trop. 55: 55-66. [ Links ]
Fourqurean, J.W., J.C. Zieman & G.v.N. Powell. 1992. Relationships between porewater nutrients and seagrasses in subtropical carbonate environment. Mar. Biol. 114: 57-65. [ Links ]
Fourqurean, J.W., A. Willsie, C.D. Rose & L.M. Rut-ten. 2001. Spatial and temporal pattern in seagrass community composition and productivity in South Florida. Mar. Biol. 138: 341-354. [ Links ]
Grasshoff, K., K. Kremeling & M. Ehrhardt. 1983. Methods of seawater analysis. Wiley-vCH, Weilheim, Germany. [ Links ]
Hemminga, M.A. & C.M. Duarte. 2000. Seagrass ecology. Cambridge Univ. Press, Cambridge, UK. [ Links ]
Herrera, W. 1984. Clima de Costa Rica. Editorial Universidad Estatal a Distancia (UNED), San José, Costa Rica. [ Links ]
Jones, A.B., M.J. ODonohue, J. Udy & W.C. Dennison. 2001. Assessing ecological impacts of shrimp and sewage effluent: biological indicators with standard water quality analyses. Est. Coast. Shelf Sci. 52: 91-109. [ Links ]
Kamp-Nielsen, L., J.E. vermaat, I. Wesseling, J. Forum & O. Geertz-Hansen. 2002. Sediment properties along gradients of siltation in South-East Asia. Est. Coast. Shelf Sci. 54: 127-137. [ Links ]
Linton, D. & T. Fisher. 2004. Caribbean coastal marine productivity program: 1993-2003. Caribbean coastal marine productivity program (CARICOMP), Kingston, Jamaica. [ Links ]
McGlathery, K.J. 1995. Nutrient and grazing influences on a subtropical seagrass community. Mar. Ecol. Prog. Ser. 122: 239-252. [ Links ]
McGlathery, K.J. 2001. Macroalgal blooms contribute to the decline of seagrass in nutrient-enriched coastal waters. J. Phycol. 37: 453-456. [ Links ]
Milliman, J.D. & R.H. Meade. 1983. World-wide delivery of river sediment to the oceans. J. Geol. 91: 1-21. [ Links ]
Moore, D.R. 1963. Distribution of the seagrass, Thalassia, in the United States. Bull. Mar. Sci. Gulf Carib. 13: 329-342. [ Links ]
Neckles, H.A., R.L. Wetzel & R.J Orth. 1993. Relative effects of nutrient enrichment and grazing on epiphyte-macrophyte (Zostera marina L.) dynamics. Oecologia 93: 285-295. [ Links ]
Nielsen-Muñoz, v. & J. Cortés. 2008. Abundancia, biomasa y floración de Thalassia testudinum (Hydrocharitaceae) en el Caribe de Costa Rica. Rev. Biol. Trop. 56 (Supl. 4): 175-189. [ Links ]
Olesen, B. & K. Sand-Jensen. 1994. Biomass-density patterns in the temperate seagrass Zostera marina. Mar. Ecol. Prog. Ser. 109: 283-291. [ Links ]
Paynter, C.K., J. Cortés & M. Engels. 2001. Biomass, productivity and density of the seagrass Thalassia testudinum at three sites in Cahuita National Park, Costa Rica. Rev. Biol. Trop. 49 (Suppl. 2): 265-272. [ Links ]
Phillips, R.C. 1960. Observations on the ecology and distribution of the Florida seagrasses. Fla. Board Conserv. Mar. Lab. Prof. Pap. 2: 1-72. [ Links ]
Preen, A.R., W.J.L. Long & R.G. Coles. 1995. Flood and cyclone related loss, and partial recovery, of more than 1000km² of seagrass in Hervey Bay, Queensland. Aust. Aquat. Bot. 52: 3-17. [ Links ]
Robblee, M.B, T.R Barber, P.R. Carlson, M.J. Durako, J.W. Fourqurean, L.K. Muehlstein, D. Porter, L.A. Yarbro, R.T. Zieman & J.C. Zieman. 1991. Mass mortality of the tropical seagrass Thalassia testudinum in Florida Bay (USA). Mar. Ecol. Prog. Ser. 71: 297-299. [ Links ]
Scheffer, F. & H. Pajenkamp. 1952. Phosphatbestimmung in Pflanzenaschen nach der Molybdän-vanadin-Methode. Z. Pflanzenernähr. Bodenk. 56: 2-8 [ Links ]
Short, F.T. & D.M. Burdick. 1995. Mesocosm experiments quantify the effects of eutrophication on eelgrass, Zostera marina. Limnol. Oceanogr. 40: 740-749. [ Links ]
Short, F.T. & S. Wyllie-Echeverria. 1996. Natural and human-induced disturbance of seagrasses. Env. Conserv. 23: 17-27. [ Links ]
Smith, S.V., W.J. Kimmerer, E.A. Laws, R.E. Brock & T.W. Walsh. 1981. Kaneohe Bay seawage diversion experiment: Perspective on ecosystem responses to nutritional perturbation. Pac. Sci. 35: 278-396. [ Links ]
Suttle, C.A. & P.J. Harrison. 1988. Ammonium and phosphate uptake kinetics of size-fractionated plankton from an oligotrophic freshwater lake. J. Plankton Res. 10: 133-149. [ Links ]
Suttle, C.A., J.A. Fuhrman & D.G. Capone. 1990. Rapid ammonium cycling and concentration-dependent partitioning of ammonium and phosphate: Implications for carbon transfer in plankton communities. Limnol. Oceanogr. 35: 424-433. [ Links ]
Terrados, J., C.M. Duarte, M.D. Fortes, J. Borum, N.S.R. Agawin, S.S. Bach, U. Thampanya, L. Kamp-Nielsen, W.J. Kenworthy, O. Geertz-Hansen & J.E. vermaat. 1998. Changes in community structure and biomass of seagrass communities along gradients of siltation in SE Asia. Est. Coast. Shelf Sci. 46: 757-768. [ Links ]
Terrados, J., P. Ramírez-García, O. Hernández-Martínez, K. Pedraza & A. Quiroz. 2008. State of Thalassia testudinum Banks ex König meadows in the veracruz Reef System, veracruz, Mexico. Aquat. Bot. 88: 17-26. [ Links ]
Tomasko, D.A. & B.E. Lapointe. 1991. Productivity and biomass of Thalassia testudinum as related to water column nutrient availability and epiphyte levels: field observations and experimental studies. Mar. Ecol. Prog. Ser. 75: 9-17. [ Links ]
UNESCO. 1989. CARICOMP–Caribbean coral reef, seagrass and mangrove sites. Coastal Region and Small Island Papers 3, UNESCO, Paris, xiv, France. [ Links ]
Valiela, I., J. Costa, K. Forman, J.M. Teal, B.L. Howes & D. Aubery. 1990. Transport of groundwater-borne nutrients from watersheds and their effects on coastal waters. Biogeochemistry 10: 177-197. [ Links ]
Valiela, I., J. McClelland, J. Hauxwell, P.J. Behr, D. Hersh & K. Foreman. 1997. Macroalgal blooms in shallow estuaries: Controls and ecophysiological and ecosystem consequences. Limnol. Oceanogr. 42: 1105-1118. [ Links ]
Zieman, J.C. 1974. Methods for the study of growth and production of turtle grass, Thalassia testudinum Konig. Aquaculture 4: 139-142. [ Links ]
Zieman, J.C., J.W. Fourqurean & T.A. Frankovich. 1999. Seagrass die-off in Florida Bay (USA): long-term trends in abundance and growth of turtle grass, Thalassia testudinum. Estuaries 22: 460-470. [ Links ]
Internet references
Waycott, M., C.M. Duarte, T.J.B. Carruthers, R.J. Orth, W.C. Dennison, S. Olyarnik, A. Calladine, J.W. Fourqurean, K.L. Heck, Jr., A.R. Hughes, G.A. Kendrick, W.J. Kenworthy, F.T. Short & S.L Williams. 2009. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Nat. Acad. Sci. www.pnas. org_cgi_doi_10.1073_pnas.0905620106 [ Links ]

