










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 suppl.1 San José Nov. 2009
Anatomía de la agalla en Ficus benjamina (Moraceae) asociada a "thrips" (Tubulifera: Phlaeothripidae)
Axel P. Retana-Salazar & Ethel Sánchez-Chacón
1. Centro de Investigación en Estructuras Microscópicas (CIEMic), Ciudad de la Investigación, Universidad de Costa Rica. 11501-2060, San José, Costa Rica; axel.retana@ucr.ac.cr
Abstract: Anatomy of the gall in Ficus benjamina (Moraceae) associated with thrips (Tubulifera: Phlaeothripidae). The galls are generally recognized as abnormal growths of tissues affected by insects when lay the eggs or feed on plant tissues. In, Ficus benjamina the insect Gynaikothrips garitacambroneroi, when feeding on leaf tissues, causes a gall which consists on the bending of the leaves. In this paper we analyzed the ultrastructure of sections of healthy leaves, and leaves with galls of F. benjamina using scanning electron microscopy (SEM). We analyzed the number of stomata per area and found no significant changes, although there is less cuticular wax on the surface of the affected area by the barb. In the leaf area affected by G. garitacambroneroi, bacilli and fungi were observed and eggs of other organisms as possible invaders. Moreover, the inner tissues of leaves with galls had some problems in the differentiation of palisade and spongy mesophylls, and only parenchymatous tissue was observed. Rev. Biol. Trop. 57 (Suppl. 1): 179-186. Epub 2009 November 30.
Key words: Gall, Ficus benjamina, Gynaikothrips spp., SEM, ultraestructure, leaf, thrips, Costa Rica.
Ficus benjamina es una de las especies ornamentales más cultivadas en el mundo. En Asia se halla ampliamente distribuida donde se exporta y utiliza como ornamental (Mound et al. 1995). Su cultivo, para exportación, se ha distribuido por todo el mundo incluyendo Costa Rica y otros países del área centroame ricana. Esta especie presenta una distribución natural desde India, el sur de China, el sudeste asiático, Malasia, Filipinas, el norte de Australia y las islas del Pacífico Sur (Riffle 1998). Ha sido introducido su cultivo en América del cano, las Islas Marshall (PIER 2001).
Taxonómicamente, F. benjamina es una insecto especie compleja que ha sido varias veces cambiada de estatus por su dificultad para identificarla con respecto a otras especies emparentadas como F. microcarpa y F. retusa. Actualmente se acepta que las especies F. nitida y F. waringiana son sinónimas de esta (Bailey & Bailey 1976, GRIN 2002). Esta misma dificultad se presenta en la diferenciación de las especies de thrips del género Gynaikothrips asociadas a estos hospederos (Garita-Cambronero & Lizano-Fallas 2006; Retana-Salazar 2006). Mound (1994) señala que al parecer las evidencias indican que las especies de thrips de este género suelen ser muy específicas con respecto a sus hospederos.
Las agallas en F. benjamina por doblez adaxial de la hoja, son consideradas crecimien-Norte y del Sur como en el istmo centroameritos alterados de los tejidos afectados que con trolan la secuencia de diferenciación celular a través de la acción de alimentación del que la produce. Los Thysanoptera, habitual mente conocidos como "thrips", son esencial mente fitófagos, lo cual evidencia una gran habilidad para explotar a sus hospederos. En el caso particular de estos insectos la formación de una agalla depende del tiempo de alimentación y del tamaño de la población dentro de la agalla (Ananthakrishnan 1981, Raman & Anathakrishnan 1986).
Los insectos inductores de agallas son herbívoros altamente especializados que se alimentan del tejido vegetal (Shorthouse & Rohfritsch 1992). Por lo general utilizan un único tipo de tejido al que pueden inducir hiperplasia o hipertrofia (Gätjens-Boniche 2007). Esta capacidad ha surgido varias veces en los insectos, lo que explica la gran diversidad de grupos no emparentados que producen agallas (Nyman & Julkunen 2000).
Sin embargo, los mecanismos precisos mediante los cuales los thrips inducen la formación de la agalla no se conocen. Los estudios taxonómicos de los grupos de thrips productores de agallas indican un avanzado grado de especificidad (Raman & Anathakrishnan 1986). El aislamiento debido a la especificidad de hospedero en los thrips galígenos determina un flujo genético muy limitado entre las poblaciones (Ananthakrishnan 1979).
Desde el punto de vista de la especialización adaptativa, tanto morfológica como funcional es sobresaliente la subfamilia Phlaeothripinae, en la que se incluyen cerca de 300 especies que inducen agallas en 200 especies de angiospermas, pertenecientes a 60 órdenes de plantas. Además de la habilidad del thrips para inducir la formación de la agalla, también existe una acción recíproca de la planta hospedera, que forma diferentes tipos de agallas que involucran diferentes grados del fenómeno de la morfogénesis de la agalla como una respuesta de la planta (Raman & Ananthakrishnan 1986).
Este estudio nació como una necesidad de algunos productores nacionales de la especie F. benjamina al enfrentarse al problema provocado por este thrips a las plantaciones, esta especie en muchos casos obligó a una reconversión agrícola, teniendo que cambiar el sistema de cultivo básico de producción, debido a que el mercado de este ornamental fue cerrado ante la presencia de esta plaga (Held et al. 2005). El principal objetivo de este trabajo fue describir, mediante el uso de la microscopia electrónica, de barrido los posibles cambios ocurridos en los tejidos de las hojas de F. benjamina provocados por G. garitacambroneroi y de esta forma tratar de colaborar con el control de esta plaga. En el trabajo de Goldazarena et al. (2008), los autores consideran que G. garitacambroneroi es sinónimo de G. uzeli.
Materiales y métodos
Las muestras se obtuvieron de árboles de F. benjamina en los alrededores de San Pedro de Montes de oca, San José, Costa Rica. Para ello se tomaron hojas sanas como control y hojas con agallas bien desarrolladas, las cuales se liberaron de thrips y otros organismos. Las hojas se limpiaron de thrips mediante la recolección con pincel humedecido en alcohol al 80%, los especimenes tanto hembras como machos e inmaduros se guardaron en alcohol al 85% como testigos de la investigación, los mismos se hallan depositados en la colección del Centro de Investigación en Estructuras Microscópicas (CIEMic), así como en la del Museo de Insectos de la Universidad de Costa Rica. Se montaron 25 thrips siguiendo el método propuesto por Mound & Marullo (1996) y se procedió a la identificación utilizando las claves para el Neotrópico, de Mound & Marullo (1996) para identificación de géneros y especies. Se confirmó la identificación mediante comparación con especímenes depositados en la colección del CIEMic, y siguiendo la descripción original de G. garitacambroneroi Retana-Salazar. Los demás organismos se removieron manualmente en observación al microscopio estereoscópico Leica MZ8, en los laboratorios del CIEMic.
Se evaluó la cobertura de estomas en hojas control y agallas utilizando un área de 150000µm2 la cual se dividió en 32 sectores de 4624µm2 (68x68). Se evaluó la cantidad de estomas por cuadrícula de muestreo y los resultados se compararon mediante una prueba de χ2. Las muestras para Microscopia Electrónica de Barrido (MEB) fueron preparadas según protocolo establecido por Sánchez et al. (2006) para muestras vegetales. Las preparaciones se hicieron en el CIEMic.
Resultados
Las superficies (adaxial y abaxial) de las hojas control de F. benjamina observadas revelaron cantidad de cera cuticular en forma de capas compactas, cubiertas por bastoncillos de cera epicuticular, esta capa es muy homogénea sobre toda la superficie (Fig. 1A, C). Mientras que las superficies de las hojas con agallas mostraron zonas de cera con bastoncillos y zonas carentes de estos (Fig. 1B, D).
En relación a los estomas, F. benjamina es una especie anfiestomática (posee estomas en ambas superficies). Las superficies de los controles parecían presentar un mayor número de estomas que las superficies de las hojas con agallas (Fig. 1A, B), pero una cuantificación por área no evidenció cambios estadísticamente perceptibles ( χ2=0.1, gl=1, p>> 0. 05).
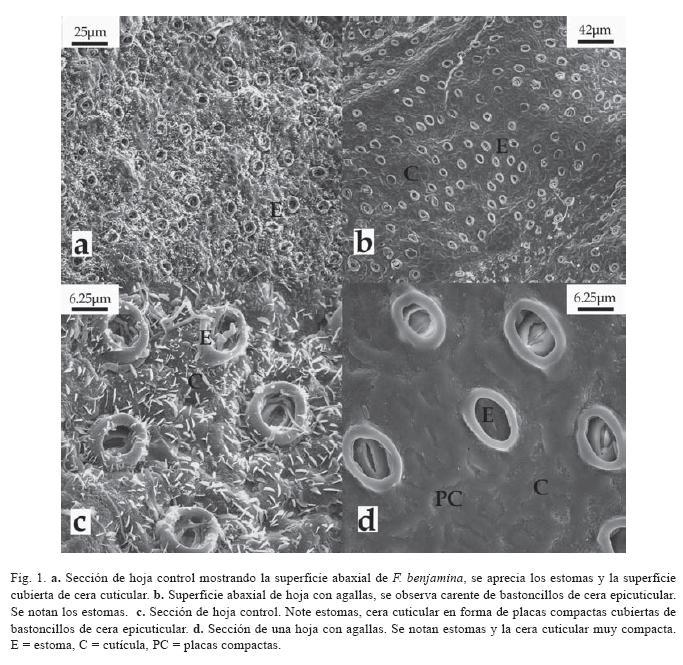
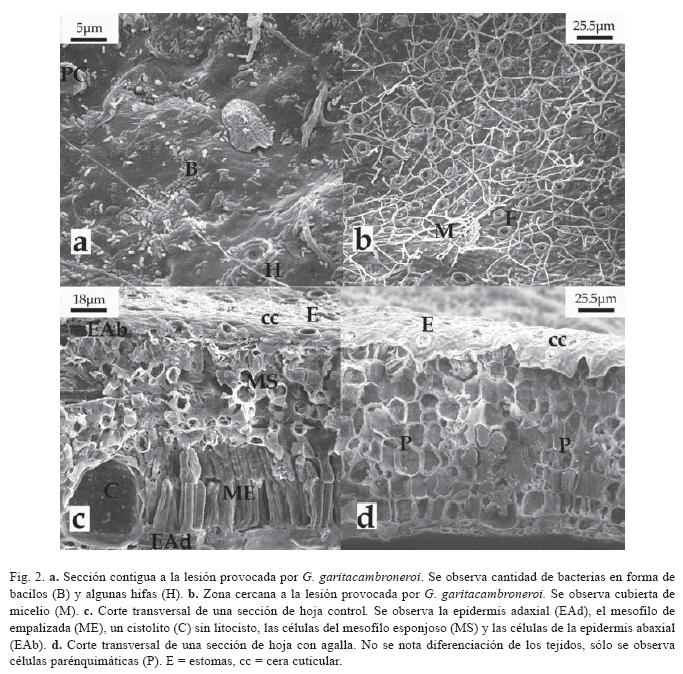
En la superficie adaxial de las hojas con agalla y los alrededores de la misma mostraron gran cantidad de bacterias baciliformes, hifas (Fig. 2A) y otros organismos, como un Coccoidea (Fig. 2A), también, algunas áreas muy cercanas a estas zonas dañadas se observaron cubiertas de micelio (Fig. 2B). La superficie abaxial no presentó estas características (Fig. 1B).
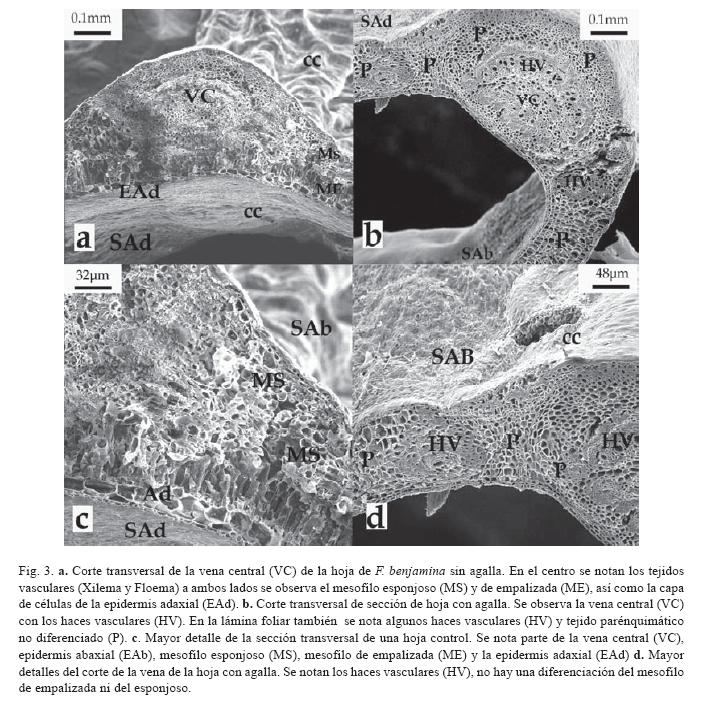
Los cortes transversales de hojas de F. benjamina no afectadas, evidenciaron una distribución y diferenciación normal de los tejidos epidérmicos (adaxial y abaxial), mesofilo de empalizada, haces vasculares y mesofilo esponjoso (Fig. 2C), por el contrario las secciones de hojas con agalla mostraron una desdiferenciación de los mesofilos de empalizada y esponjoso (Fig. 2D), aunque los tejidos epidérmicos y las venas centrales y menores conservaron su estructura normal (Fig. 3B, D). Al parecer, fue más evidente la presencia de látex en las secciones controles que las secciones con agallas, ya que cerca de las venas y otras zonas de las hojas control era difícil detallar las diferentes estructuras debido al látex expuesto al realizar el corte, pero aún así, se pudo diferenciar los diferentes tejidos que componen una hoja normal, como son: la epidermis adaxial, mesofilo en empalizada, haces vasculares, mesofilo esponjoso y epidermis abaxial (Fig. 3A, C)
Discusión
Las agallas producidas por los thrips son generalmente abiertas, excepto en el caso del género Johansenthrips Retana-Salazar y Soto-Rodríguez hallado en el Neotrópico (Retana-Salazar & Soto-Rodríguez 2007). Las agallas abiertas contienen poblaciones de la especie inductora, donde se registran todos los estadios de desarrollo. También se hallan con frecuencia otras especies de insectos y artrópodos que pueden ser inquilinos o depredadores de los inductores de la agalla (Crespi et al. 2003, Heds et al. 2005). La presencia de múltiples especies convierte a las agallas en comunidades con relaciones complejas. Los thrips generadores de agallas son más abun dantes en las zonas tropicales del Viejo Mundo y Australia. Sin embargo, estos grupos perte necen a linajes muy diversos mientras que los inductores de agallas en el Neotrópico parecen pertenecer todos ellos al linaje de los Liothrips, esto nos da la ventaja en el estudio de la traza filogenética del proceso de inducción de agallas (Mound & Morris 2005).
El proceso de morfogénesis de las agallas es complejo ya que involucra la recanalización del desarrollo de la planta por parte del insecto inductor (Ananthakrishnan 1998). Los estudios de la morfogénes de las agallas se han centrado en aquellas que se consideran de mayor complejidad, es decir las agallas cerradas producidas por Cecidomyiidae o Tephritidae. En estos casos se ha demostrado que los insectos inductores de las agallas activan regiones específicas de la planta hospedera, provocando un gradiente bioquímico y citológico a partir de las células más cercanas al sitio donde se halla el insecto (Shorthouse & Rohfritsch 1992).
Tradicionalmente se ha considerado que las agallas generadas por la acción de thrips y chinches son pequeños crecimientos anormales conformados por tejido parenquimatoso, lo que confirma los cambios observados en este estudio. Además, algunas especies producen enrollamiento de las hojas acompañado de hipertrofia celular (Gätjens-Boniche 2007). Esta aparente simpleza estructural ha generado que estas agallas sean menos estudiadas aunque los daños producidos en especies ornamentales de exportación son cuantiosos.
Ananthakrishnan y Raman (1989) en sus trabajos con agallas que producen enrollamiento dedican los estudios estructurales a aquellas agallas que generan modificaciones del crecimiento del tejido. Esto ha hecho que grupos que generan agallas por doblez y enrollamiento de la lámina foliar sean menos estudiados, debido a que las deformaciones producidas son menos evidentes.
Gynaikothrips garitacombroneroi, descrita recientemente dentro del complejo uzeli-ficorum, es una especie agresiva que ha reducido la producción nacional de Ficus ornamentales. Esto ha motivado el estudio pormenorizado de la morfogénesis de las agallas inducidas por esta especie de thrips donde los resultados de la ultraestructura demuestran que este proceso es bastante más complejo que el informado por Ananthakrishnan & Raman (1989) para especies de la misma familia con formaciones de agallas similares.
Según Ananthakrishnan & Raman (1989), la hoja con agallas no presenta zonas de crecimiento anómalo, como se ha observado con algunas especies de Liothrips y Gynaikothrips. Sin embargo, los resultados obtenidos en este estudio, indican que en efecto, hay un cambio en la estructura de las células que constituyen la lámina foliar al ser comparadas con la ultraestructura de hojas sanas donde sí hay una división clara de los diferentes tejidos que componen la hoja.
Lo anterior nos lleva a replantear las consideraciones tradicionales acerca de la polaridad de las agallas como caracteres evolutivos de los grupos de insectos. Se considera que las agallas que inducen deformación de los tejidos foliares son las más derivadas (Gätjens-Boniche 2007). Sin embargo, las evidencias ultraestructurales presentadas en este trabajo indican que es posible que las agallas más derivadas sean aquellas en las que se da una regulación total del tejido de la lámina foliar y no una modificación parcial de este tejido.
El que una especie de insecto pueda promover que todo un órgano vegetal mantenga una determinada estructura celular, indica la presencia de un sistema de regulación mucho más complejo que el estudiado en las agallas que evidencian crecimientos particulares, donde la estratificación o diferenciación de los tejidos se da por el gradiente de difusión a través de las capas celulares de sustancias que a menor concentración determinan mayor madurez del tejido (Gätjens-Boniche 2007). Un insecto que presenta la habilidad de mantener un tipo de tejido en determinadas condiciones particulares de madurez, y que además este tejido no pierda la funcionalidad para la planta, y que sea al mismo tiempo nutritivo para el insecto, parece involucrar regulaciones más complejas y puede tener un valor evolutivo y adaptativo mayor.
La especie de thrips que induce estas agallas es compleja en su determinación y se segrega por diferencias a nivel de genitalia del macho (Retana-Salazar 2006). Goldarazena et al. (2008) sinonimizan G. garitacambroneroi con G. uzeli de Asia, con base en la revisión de un paratipo de hembra de G. garitacambroneroi (depositado en el Museo de Historia Natural de Londres), y en haber encontrado en algunos especímenes de G. uzeli del sur-este de Asia e Islas Maldivas variación morfológica que se traslapa con la informada por Retana-Salazar (2006) para G. garitacambroneroi. Aunque estos no son criterios suficientes para establecer la sinonimia, los resultados acerca del desarrollo de la agalla inducida por esta especie contrastan grandemente con lo informado en la literatura acerca de la formación de agallas del mismo tipo por otras especies del género Gynaikothrips (Ananthakrishnan & Raman 1989). Esto motiva aún más el proponer criterios complementarios que permitan la identificación rápida y segura de estos insectos, en particular aquellos asociados a agallas.
Agradecimientos
Los investigadores agradecen a la Vicerrectoría de Investigación de la Universidad de Costa Rica, ya que mediante los proyectos 810A6-228 y 810-A6-239 de esta Vicerrectoría fue posible la realización de este trabajo.
Resumen
Las agallas son reconocidas generalmente como crecimientos anormales de los tejidos afectados por insectos cuando estos realizan la deposición de los huevecillos o se alimentan de los tejidos de la planta. En Ficus benjamina la acción del thrips Gynaikothrips garitacambroneroi al alimentarse de los tejidos de la hoja, provoca una agalla que consiste en el doblamiento de la hoja. En este trabajo se analizó la ultraestructura de secciones de hojas sanas y hojas con agallas de F. benjamina mediante el uso de la microscopia electrónica de barrido. Se analizó la cantidad de estomas por área y no se determinó alteración significativa, aunque se observa menor cantidad de cera cuticular en la superficie del área afectada por la agalla. En la zona de la hoja afectada por G. garitacambroneroi se observó bacilos y hongos y huevecillos de otros organismos, aparentemente invasores. Además, en los tejidos internos de las hojas con agallas se pudo determinar problemas de diferenciación de los mesófilos de empalizada y esponjoso, observándose sólo tejido parenquimático.
Palabras clave: Agalla, Ficus benjamina, Gynaikothrips spp., microscopia de barrido, ultraestructura, hoja, thrips, Costa Rica.
Recibido 04-X-2007. Corregido 20-IX-2009. Aceptado 21-X-2009.
Referencias
Ananthakrishnan, T.N. 1979. Biosystematics of Thysanoptera. Ann. Rev. Entomol. 24: 159-183. [ Links ]
Ananthakrishnan, T.N. 1981. Thrips-plant gall association with special reference to patterns of gall diversity in relation to varying thrips populations. Proc. Indian Nat. Sci. Acad. 47: 41-46. [ Links ]
Ananthakrishnan, T.N. 1998. Insect gall systems: Patterns, processes and adaptative diversity. Current Sci. 75: 672-676. [ Links ]
Ananthakrishnan, T.N. & A. Raman. 1989. Thrips and gall dynamics. oxford & IBH Publishing Co. PVT. Ltd., Nueva Delhi, India. [ Links ]
Bailey, L.H. & E.Z. Bailey. 1976. Hortus. Macmillan General Reference, Nueva york, EEUU. [ Links ]
Crespi, B.J., D.C. Morris & L.A. Mound. 2003. Evolution of Ecological and Behavioural Diversity: Australian Acacia Thrips as Model organisms. Australian Biological Resources Study & Australian National Insect Collection, CSIRo, Canberra, Australia. [ Links ]
Garita-Cambronero. J. & J. Lizano-Fallas. 2006. Determinación de dimorfismo sexual de Gynaikothrips garitacambroneroi (Thysanoptera: Phlaeothripidae) inductos de agallas en Ficus benjamina. Métodos Ecología Sistemática 1: 9-13. [ Links ]
Gätjens-Boniche, o. 2007. Agallas de plantas inducidas por insectos: un sistema natural de transformación genética en células eucarióticas. Tesis de Maestría en Biología, Universidad de Costa Rica, San José, Costa Rica. [ Links ]
Goldarazena, A., L.A. Mound & R. zur Strassen. 2008. Nomenclatural problems among Thysanoptera (Insecta) of Costa Rica. Rev. Biol. Trop. 56: 961-968. [ Links ]
Held, D.W, D. Boyd, T. Lockley & G.B.E. Dwards. 2005. Gynaikothrips uzeli (Thysanoptera: Phlaeothripidae) in the Southeastern United States: distribution and review of biology. Florida Entomol. 88: 538-540. [ Links ]
Mound, L.A., C.L. Wang & S.Okajima. 1995. Observations in Taiwan on the identity of the Cuban laurel thrips (Thysanoptera, Phlaeothripidae). J. New york Entomol. Soc. 103: 185-190. [ Links ]
Mound, L.A., & R. Marullo. 1996. The thrips of Central and South America: an introduction. Mem. Entomol. Internat. 6: 1-488. [ Links ]
Mound, L.A. & D.C. Morris. 2005. Gall-inducing Thrips: An Evolutionary Perspective, p. 59-72. In A. Raman, C.W. Schaefer & T.M. Withers (eds.). Biology, ecology, and evolution of gall-inducing arthropods. Science Publishers, New Hampshire, EEUU. [ Links ]
Nyman, T. & T. Julkunen. 2000. Manipulation of phenolic chemistry of willows by gall-inducing sawflies. Proc. Nat. Acad. Sci. USA 97:13184-13187. [ Links ]
Raman, A. & T.N. Ananthakrishnan. 1986. Biology of Gall Thrips (Thysanoptera: Insecta), p. 107-127. In T.N. Ananthakrishnan (ed.). Biology of gall insects. Edward Arnold, Londres, Inglaterra. [ Links ]
Retana-Salazar, A. 2006. Variación morfológica del complejo Gynaikothrips uzeli-ficorum (Phlaeothripidae: Tubulifera). Métodos en Ecología y Sistemática 1: 1-9. [ Links ]
Retana-Salazar, A.P. & G.A. Soto-Rodríguez. 2007. Revisión taxonómica del grupo Haplothrips-Karnyothrips (Thysanoptera: Phlaeothripidae). Rev. Biol. Trop. 55: 627-635. [ Links ]
Riffle, R.L. 1998. The tropical look. Timber, Inc., Portland, oregon, EEUU. [ Links ]
Sánchez, E., T. Quesada & A.M. Espinoza. 2006. Ultrastructure of the wild rice Oryza grandiglumis (Gramineae) in Costa Rica. Rev. Biol. Trop. 54: 377-385. [ Links ]
Shorthouse J.D. & o. Rohfritsch. 1992. Biology of the insect-induced galls. oxford University, Nueva york, EEUU. [ Links ]
Referencias de internet
GRIN (Germplasm Resources Information Network). 2001. online Database.
United States Department of Agriculture, Agricultural Research Service, National Germplasm Resources Laboratory, Beltsville, MD. (Accessed: February 7, 2002; http://www.ars-grin. gov/).
PIER (Pacific Island Ecosystems at Risk). 2001. Invasive plant species: Ficus benghalensis L., Moraceae. Pacific island ecosystems at risk. (Accessed: January 16, 2002; http://www.hear.org/pier/). [ Links ]

