










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 suppl.1 San José Nov. 2009
Patterns of nectar production and composition, and morphology of floral nectaries in Helicteres guazumifolia and Helicteres baruensis (Sterculiaceae): two sympatric species from the Costa Rican tropical dry forest
Loretta Goldberg
Escuela de Biología, Universidad de Costa Rica, 11501-2060, San José, Costa Rica. Fax 506-22809294; goldberg_loretta@yahoo.com
Abstract: Helicteres guazumifolia Kunth and Helicteres baruensis Jacq. (Sterculiaceae) are two sympatric species of shrubs common along the North Western tropical dry forest of Costa Rica. i recorded their nectar production within a 24 hour cycle. i also describe the morphology of extrafloral nectaries with scanning electron microscopy. in H. guazumifolia secretion was restricted to the first day of flower life span, shortly after anthesis (0600 hr - 1800 hr). Flowers secreted on average 15.63 ±8.45 µl (N=409). Nectar is composed of three main sugars: sucrose, fructose and glucose (mainly sucrose). A total of 17 free amino acids were identified: mainly proline, arginine, threonine and tyrosine, with a concentration above 70 Ng/µl. values were different for H. baruensis. Nectar secretion was confined to the second day after anthesis, starting at 1600 hr and ending at 0600 hr the following day. Flowers secreted on average 77.03 ±64.99 µl (N=163) of nectar. Nectar is also composed of three main sugars; however, it showed a tendency to be hexose-rich, having more fructose and glucose than sucrose. There were also 17 free amino acids, mainly proline, alanine, tyrosine, arginine and threonine. Patterns of nectar production are different between the two species for timing, and for amount and composition of nectar secretion. Rev. Biol. Trop. 57 (Suppl. 1): 161-177. Epub 2009 November 30.
Key words: Amino acids, Helicteres guazumifolia, Helicteres baruensis, floral nectary, nectar, pollination, reproductive biology, sugars, Costa Rica.
Nectar is one of the major primary attractants and rewards of angiosperm flowers to their pollinators (Baker & Baker 1973b, Gottsberger et al. 1984, Freeman et al. 1985, Stiles & Freeman 1993, Endress 1994, Galetto 1997). it plays a central role in plant reproduction by mediating plant-pollinator interactions due to its inherent features such as sugar concentra tion, volume, viscosity and chemical composition (Cruden 1976, Cruden et al. 1983, Galetto & Bernardello 1995, Perret et al. 2001, McDade & Weeks 2004). Since the pioneer works of Baker and Baker in the seventies, the nectar of hundreds of species from temperate and tropical regions has been studied in some detail (Gottsberger et al. 1984, Baker 1990). information is available for a few hun dred species for the most basic nectar traits such as sugar concentration and volume per flower (McDade & Weeks 2004). in addition, considerable attention has been given to eco logical, chemical and phylogenetic investigations of nectar (Gottsberger et al. 1984, Baker & Baker 1990, Gottsberger et al. 1990).
Sugars are present in all floral nectars in greater amount than any other constituent (Baker & Baker 1973b, 1976, Baker 1977). The three most common sugars found in nectar are sucrose, fructose and glucose. They dominate the total solutes and may be present in varying proportions (Freeman et al. 1985, Martínez & Baker del Río 1990, Stiles & Freeman 1993, Endress 1994, Proctor et al. 1996, Galetto 1997, Baker et al. 1998). Nectars may contain sucrose only or different combinations of these three sugars (Cruden et al. 1983, Proctor et al. 1996) and may vary between species (Murcia 2002).
Amino acids are the second most important group of components found in nectar. Essential amino acids are present in all species (Baker & Baker 1973a, b, 1976, Murcia 2002); however, their importance in the nutrition of pollinators is far from being clear. Also, it is not clear if there are regular patterns of amino acids content in nectar (Gottsberger et al. 1989, 1990). Other substances present in minor proportions are lipids, antioxidants, alkaloids, phenolic substances and glycosides (Baker 1977, Endress 1994).
Most studies on nectar chemistry have been designed to reveal convergence among ecological or geographical plant communities, but few concern nectar comparisons between species belonging to the same genus or tribe (Perret et al. 2001). Vogel (1990) discussed the parallel radiation of Neotropical plants into different pollination modes by describing the floral syndromes of the family Sterculiaceae and the corresponding pollinators. Members of the genus Helicteres provide an outstanding example of floral diversification and pollinator use. Floral syndromes include psychophily, sphingophily, ornitophily, and chiropterophily (vogel 1990, von Helversen & voigt 2002).
Two sympatric species within the genus Helicteres, with contrasting pollination syndromes, occur along the North Western tropical dry forest of Costa Rica. Helicteres guazumifolia Kunth which is hummingbird pollinated and Helicteres baruensis Jacq. known to be bat pollinated. The general objective of this study was to characterize nectar production patterns of both species according to the time of the day or the night in which nectar is secreted, in order to: (1) determine the quantity in terms of volume (µl) of nectar produced; (2) analyse the nectar composition by the identification and quantification of sugars and amino acids; (3) compare the type of nectar produced in these two species in the light of their contrasting pollination syndromes. Also, (4) the morphology of the floral nectaries of both species was analyzed by aids of scanning electron microscopy.
Materials and methods
Study site: This study was conducted in Santa Rosa National Park, Guanacaste Conservation Area (ACG), North Western Costa Rica (10°45 to 11°00 N and 85°30 to 85°45 W). Two life zones are present in the area and the study was conducted in a tropical dry forest, with a moist transition (Holdridge 1967, Hartshorn 1991). The park includes a mosaic of forests of different ages and abandoned pastures (Janzen 1986, Hartshorn 1991, Gerhardt 1993).
In the past, the zone was covered by extensions of tropical dry forest, the most threatened ecosystem of Mesoamerica, it originally covered about 550 000 km2 from Mazatlán in Mexico until Panama Canal. Actually, only 2% is maintained and 25% of the surface is protected, represented principally in the Guanacaste Conservation Area (Janzen 1986, Fernández Morillo 1998).
The climate is highly seasonal, with a well defined dry season that goes from late November to mid May. Annual rainfall ranges between 800 and 2 600 mm, with an annual mean of 1 423.4 mm. Annual mean temperature is 25.7 °C and annual mean relative humidity is 81% (Rojas Jiménez 2001).
Study species: Helicteres is a pantropical genus that contains approximately 60 species, native to the tropics of both hemispheres (Robyns & Cuatrecasas 1964, Gentry 1993, Cristóbal 2001a, Bayer & Kubitzki 2003). it is most abundant in America in which 38 species are distributed from Mexico, Central America, the Caribbean and South America through North Western Argentina and slightly South of the tropics line in Eastern Paraguay and Brazil (Sazima & Sazima 1988, Cristóbal 2001a, b). The members of the genus are shrubs or small trees of dry lowland areas (Sazima & Sazima 1988). They are characterized by having distinctive fruits, which are spiral capsules, many seeded and with a long androgynophore, free or fused (Robyns & Cuatrecasas 1964, Gentry 1993, Bayer & Kubitzki 2003). Two species of Helicteres are found in Santa Rosa National Park: H. guazumifolia and H. baruensis.
H. guazumifolia is widely spread and covers greatest area in America. it extends from Southern Mexico to Central America, North Western Cuba until Rondonia and West of Mato Grosso and neighboring zones of Bolivia (Cristóbal 2001a). it is a shrub or small tree 0.50-5 m high, ramificated from the base or erect with slender branches (Robyns & Cuatrecasas 1964, Cristóbal 2001a, b). Flowers are axillary, erect, actinomorphic and have a tubular corolla with a basal nectary. They have short red and spatulated petals, and the peduncle is aligned with the androgynophore (Robyns & Cuatrecasas 1964, Gentry 1993, Cristóbal 2001a,b). it is found on open, secondary and semideciduous forests, gallery forests, pastures and zones of periodic fires and clearings, also on dry or moist thickets, grassy or bushy slopes (Robyns & Cuatrecasas 1964, Cristóbal 2001a, b).
H. baruensis is also widely distributed in the Americas. it extends from the Pacific coast of Mexico, South of Sonora until Oaxaca, in Yucatan Peninsula, the Caribbean and South America until Colombia, venezuela, Suriname, Guyana and Brazil (Cristóbal 2001a). it is a shrub or slender tree 2-6 m high with dense foliage. Flowers are geniculated and are borne in axillary or oppositifolious inflorescences, usually three to five flowered and have a horizontal position. They are zygomorphic and have a tubular corolla with a basal nectary, and also have two or more nectaries at the base of the pedicels between the flowers. The petals are pale greenish and acintated, and the androgynophore is bent (Robyns & Cuatrecasas 1964, Cristóbal 2001a, b). This species is characteristic of dry caducifolious forests, holms-oak forests and gallery forests (Cristóbal 2001a, b).
Nectar production: Flower production of H. guazumifolia and H. baruensis was monitored from March 2003 to March 2004. H. guazumifolia produced flowers during March to late June 2003, and H. baruensis since July to late December 2003. Nectar volume was measured when each species produced flowers.
A total of five mature flower buds per plant were selected randomly and were bagged one day prior to anthesis using cheesecloth bags for each observation period. A total of 15 different plants were selected randomly from each population for every day of observation. The accumulated nectar was sampled each hour, on newly opened flowers over the course of each day. Measurements were conducted for H. guazumifolia within the period from 0600 hr to 1800 hr and for H. baruensis from 1600 hr to 0600 hr of the following day.
To determine nectar production, the volume (µl) of nectar secreted was measured using calibrated micropipettes (Drummond Scientific Company Wiretrol®, for H. guazumifolia of 10-20 µl, and for H. baruensis of 25-50 µl and 50+100 µl). Nectar production was determined as the volume of nectar secreted over the course of the day after anthesis. Floral nectar was measured only once on each flower; and they were manipulated with care preventing floral damage, contamination by pollen grains or tissue secretions. Nectar samples were stored in 1.5 ml vials containing 1 ml of 70% ethanol for preservation. They were kept frozen at -20 ºC until analysis.
Nectar composition: The study of the chemical composition of nectar was conducted in the Biology Laboratories, Ulm University, Germany. These analyses were conducted using High Performance Liquid Cromatography (HPLC) (Linskens & Jackson 1987). Sugar content was determined examining the concentration (micrograms/microliters) of three sugars: sucrose, glucose and fructose. in addition, amino acid concentration (nanograms/ microliters) was also determined from nectar samples from the two species of Helicteres. Sugar analyses with high performance liquid chomatography allow the determination of nectar composition with greater precision and accuracy (Freeman & Wilken 1987). The analytical methodology has been outlined previously by Elisens & Freeman (1988).
Morphology of floral nectaries: The study of the morphology of the floral nectaries was also conducted in the Biology Laboratories, Ulm University. Samples of the floral nectaries of both species were critically point dried with the critical point drying apparatus model E3000. Dried samples were mounted on microscope stubs and were vacuum coated sputtered with gold. The sputtering device used was model Balzers Union FL-9496. Micrographs were taken using a JSM-Si Scanning Microscope.
Statistical analyses: Statistical analyses were conducted using StatisticaTM (release 6.0). One way analysis of variance was used to compare mean nectar volume produced through the course of the day or night for each species. Multivariate analyses of variance were used to compare mean sugar concentration and mean amino acid concentration for each species.
Results
Nectar production
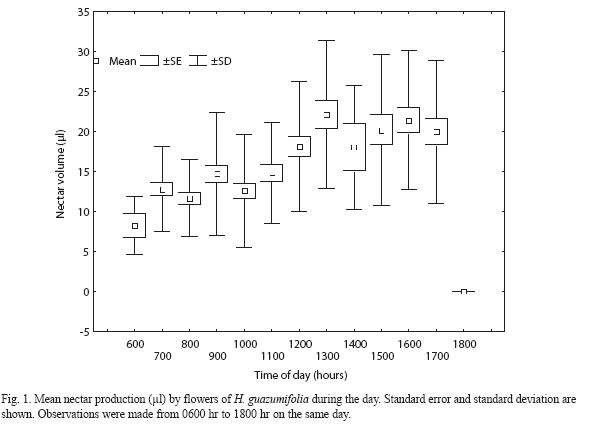
H. guazumifolia. Nectar production in flowers of H. guazumifolia begins shortly after their anthesis, approximately at 0600 hr and ceases at 1800 hr. it is produced only during the first day of flower life span. Daily nectar production ranged between 8.29 and 22.08 µl. Analysis of variance revealed significant variation in mean nectar production (µl) during the course of the day (F=12.30, df=12 and 396, p<0.0001). On average, a total of 15.63 ±8.45 µl (N=409) of nectar were secreted through the day.
Nectar secretion remained fairly constant until 1100 hr, it increased slightly at 1200 hr, reaching a maximum peak of 22.08 µl at 1300 hr. During the following afternoon hours high quantities of nectar secreted were maintained, ranging from 18.03 µl to 22.08 µl. Secretion ceased drastically by 1800 hr in most flowers sampled (Fig. 1).
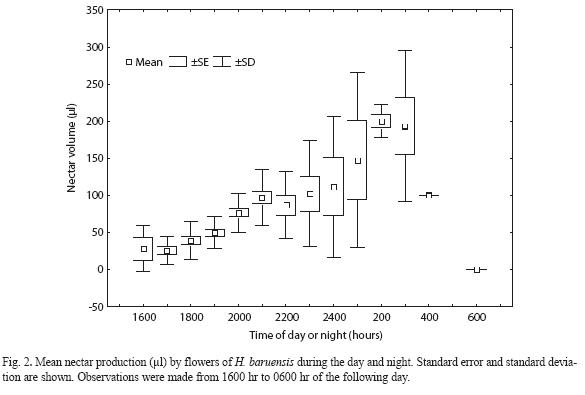
H. baruensis. Flowers of H. baruensis initiate nectar secretion during the second night after anthesis. Flowers secreted nectar only during one single night. Secretion started around 1600 hr and ceased around 0600 hr of the following day. Mean nectar volume (µl) produced varied significantly during the course of late afternoon and night times (F=13.64, gl=14 and 148, p<0.0001). Flowers secreted an average amount of 77.03 ±64.99 µl (N=163) of nectar.
Nectar volume secreted between 1600 hr and 1700 hr was the lowest ranging between 27.88 µl and 25.21 µl. An increase in nectar secretion occurred since 1900 hr. The maximum production was recorded around 0200 hr of the following day. On average a total of 199.82 µl of nectar were produced at this time. A reduction in secretion occurred after 0400 hr. At 0500 hr, mean nectar production was only 50 µl. At 0600 hr in the early morning most flowers have ceased nectar production (Fig. 2).
Sugar concentration
H. guazumifolia. Floral nectar of H. guazumifolia contained three main sugars: sucrose, fructose and glucose. Mean sugar composition varied significantly (F=2.37, df=33 and 106, p=0.0005) during the course of the day. Nectar differs in concentration and predominant sugar type. Sucrose predominates as the main sugar and it is significantly more concentrated than glucose and fructose (F=4.08, df=11 and 38; p=0.0005). The mean concentration of sucrose was 292.69 ±114.99 µg/µl (N=50).
Sucrose concentration in nectar was higher during the first morning hours, reaching a value of 650.76 µg/µl at 0600 hr (Fig. 3). it declines steadily until 0900 hr, reaching 219.93 µg/µl. A slight peak was recorded at 1300 hr and at 1600 hr; on average, 334.36 µg/µl and 335.94 µg/µl were produced respectively. Sucrose concentration remained fairly constant during the rest of the day.
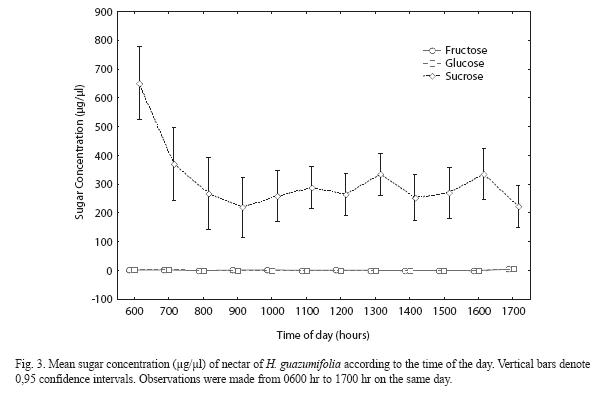
Mean concentration of fructose and glucose was low during the course of the day (1.68 ±2.87 µg/µl and 1.02 ±2.54 µg/µl, F=1.35, df=11 and 38; p=0.24 and F=1.98, df=11 and 38; p=0.06, respectively). Sucrose was present in all nectar samples regardless of the time of the day. in contrast, fructose was not found in samples taken at 0800 hr and 1600 hr. No glucose was found in samples from 0800 hr, 0900 hr, 1100 hr, 1200 hr, 1400 hr and 1500 hr.
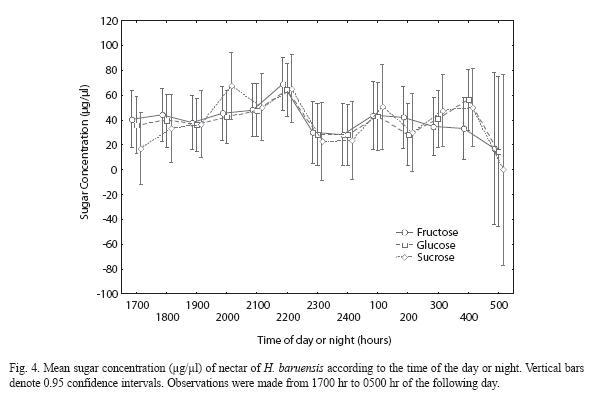
H. baruensis. Floral nectar of H. baruensis also contained three main sugars: sucrose, fructose and glucose. There were no significant differences in mean concentration between the three types of sugars (F=1.12, df=36 and 204, p=0.30). Nectar concentration, as well as the predominant sugar type did not vary significantly through sampling time. Overall mean sugar concentration for sucrose, fructose, and glucose were 41.52 ±39.31 µg/µl, 41.94 ±30.32 µg/µl, and 41.11 ±30.15 µg/µl, respectively. However, a tendency for nectar to have more fructose and glucose over sucrose was observed at the following hours: 1700 hr, 1800 hr, 2200 hr, 2300 hr, 2400 hr, 0200 hr, 0400 hr and 0500 hr (Fig. 4).
Amino acid concentration
H.s guazumifolia. The nectar of H. guazumifolia contained a total of 17 different free amino acids. There were no significant differences between amino acid concentrations and the different sampling times. On average, proline was the most abundant amino acid, with a mean concentration of 554.22 ±391.64 Ng/ µl (F=3.69, df=3 and 20, p=0.03). As shown in Table 1, arginine, threonine and tyrosine were significantly more abundant (more than 70 Ng/ µl) than the rest of the amino acids found in the nectar.
The following amino acids were found in low concentrations (less than 50 Ng/µl) on every sampling period: methionine, lysine, serine, valine, histidine, aspartic acid, leucine and glycine. Cysteine was the only amino acid absent from every sample period. Meanwhile, glutamic acid, alanine, isoleucine and phenylalanine were found only in some samples.
H. baruensis. Similar to its related species H. guazumifolia, floral nectar of H. baruensis contained a total of 17 different free amino acids (Table 2). There were no significant differences in amino acid concentration between sampling times. Proline, alanine, tyrosine, arginine, and threonine were more concentrated than the other amino acids (more than 20 Ng/µl). Proline showed the highest concentration (146.56 ±100.82 Ng/µl) among these amino acids.
Even though they were found in low concentrations (less than 10 Ng/µl), valine and lysine were found in every sampling period. Glycine, methionine, and leucine were absent in every sample. Aspartic acid, serine, glutamic acid, histidine, cysteine, isoleucine and phenylalanine were absent in two sampling periods (1700-1800 hr and 2100-2200 hr).
Both Helicteres species share the same types of amino acids that were found to be the most concentrated, with the exception of alanine, which was present on H. baruensis and absent from H. guazumifolia on sampling time 1200-1400 hr.
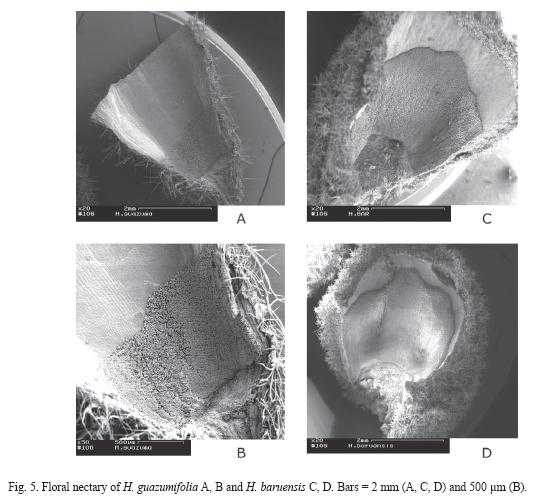
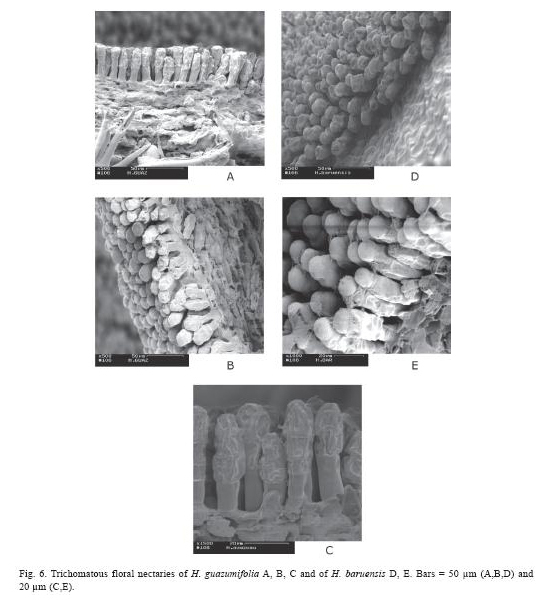
Floral nectaries morphology: The floral nectary of H. guazumifolia is located at the base of the calyx, and has an average length of 1.54 ±0.34 mm (N=5) with respect to the total length of the corolla (Fig. 5. A-B). As in H. guazumifolia, the floral nectary of H. baruensis is also located at the base of the calyx, and has an average length of 2.83 ±0.70 mm (N=11) with respect to the total length of the corolla (Fig. 5. C-D). in both species, the floral nectary is formed by a distinct group of trichomes that form a particular carpet (Fig. 6.).
Discussion
Nectar production
Nectar production is a dynamic process that involves continuous processes in the lifespan of every flower such as secretion, reabsorption and evaporation (Gottsberger et al. 1989, 1990). An enormous amount of descriptive work has been published on patterns of floral nectar production. Nectar production may be affected by time of day or season, flower age, size or stage, flower location on the plant, defoliation, soil moisture and weather conditions (Gottsberger et al. 1984, Zimmerman 1988, Witt et al. 1999). Other selective pressures that can influence nectar production besides pollinator class are flower density, habitat, nectar thieves and breeding system (Cruden et al. 1983). As a result, rates of nectar production among plants in populations have been found to differ in variability (Zimmerman 1988). The two Helicteres species showed differences in the amount and composition of the nectar produced, as it varies between different species (Witt et al. 1999, Murcia 2002).
Helicteres guazumifolia: it has generally been stated that flowers pollinated by high energy requiring animals such as hummingbirds and bats tend to produce high amounts of nectar (Cruden et al. 1983, Stiles & Freeman 1993, Proctor et al. 1996), such as the ones registered for each Helicteres species. Patterns of nectar production are clearly distinguished between the two species. Nectar is secreted in different amounts and at different times of the day and night; in addition, both species provide a great energetic reward.
Flowers pollinated by diurnally active animals produce nectar during the day (Cruden et al. 1983) and this pattern was observed for H. guazumifolia flowers which started secretion at 0600 hr. Regardless of quantification of hummingbird visits to the flowers, several individuals of the hummingbird Amazalia rutila were observed hovering at flowers early in the morning after 0700 hr and late in the afternoon after 1500 hr. After noon and during the rest of the day, flowers contained higher amounts of nectar that could be offered to them. in most species nectar is secreted at a constant rate until some critical amount has accumulated, then ceases at approximately the same time that hummingbirds cease daily activity, approximately at 1830 hr (Cruden et al. 1983). This may explain why secretion was stopped by 1800 hr after having nectar available during most of the day.
In unbagged flowers exposed to pollinators, it is possible that nectar secretion ceases if pollinators are inactive or that is being reabsorbed in old or pollinated flowers (Cruden et al. 1983). Reabsorption could occur at the end of the day, when hummingbirds are not active anymore. Further studies under these conditions could help to understand if H. guazumifolia flowers show this type of adaptive pattern on the production of nectar. Galetto & Bernardello (1995) in their study of nectar secretion of two Lycium species, showed that nectar was reabsorbed at the end of flower lifetime. A nectar production pattern with no reabsorption may have an impact on reproductive biology (Zimmerman 1988, Galetto & Bernardello 1995). The seed numbers of a plant may decrease due to the costs of producing nectar (Witt et al. 1999). So, plants reabsorb nectar from aging flowers and utilize its carbon in developing seeds with a consequent reproductive advantage (Zimmerman 1988, Galetto & Bernardello 1995).
Helicteres baruensis: This species belongs to the group of plants specifically adapted to the pollination by bats of the subfamily Glossophaginae, Glossophaga soricina (von Helversen & voigt 2002). According to Cruden et al. (1983), flowers that are pollinated by nocturnally active animals produce nectar at night and this was the nectar secretion pattern registered for the species. Secretion of nectar is much greater in bat flowers than in all other pollination syndromes (von Helversen 1993, Endress 1994, Murcia 2002, Tschapka & Dressler 2002). H. baruensis flowers secreted nectar only during a single night, and produced on average 77.03 +64.69 µl (N=163), which is considered a high amount. Nectar glands of this species are voluminous in comparison to those of related species that are not bat pollinated (von Helversen 1993). Large flowers with a deeper corolla tube produce a higher volume of more diluted nectar than smaller flowers (Cruden & Herman 1983, Galetto & Bernardello 1995), and that is why H. baruensis secreted more nectar than its related species H. guazumifolia. The relatively large flowers and large amounts of nectar are among the traits associated with the syndrome of chiropterophily (Tschapka et al. 1999).
The timing of nectar production is correlated with the activity cycle of the pollinator (Cruden 1976, Cruden et al. 1983). Nectar secretion started at 1600 hr and incremented hourly since 1900 hr, initiation of secretion may occur over a period of an hour or more (Cruden & Herman 1983). initiation of secretion is so timed that sufficient amounts of nectar are present when pollinators become active (Cruden et al. 1983). H. baruensis flowers initiated nectar secretion around two to four hours prior to bat activity, which was registered to occur from 1800 hr to 2300 hr by von Helversen & voigt (2002). As shown in Figure 2, maximum amounts of nectar started to be produced from 2100 hr, five hours later since secretion started. From this time on flowers contained maximum amounts of nectar available for its main pollinator, reaching a peak of 199.82 µl.
After flowers had accumulated critical amounts of nectar available for its pollinator, reduction in secretion started to occur, again while pollinators were inactive. von Helversen & voigt (2002) proposed that sugar was probably reabsorbed actively by H. baruensis flowers in the morning hours as they had little or no nectar during the day, even in flowers that had been bagged during the whole preceding night. By 0500 hr, flowers contained only 50 µl, and none of the flowers sampled at 0600 hr contained nectar.
Some H. guazumifolia plants were located in an open area close to few individuals of H. baruensis. Between 1600 hr and 1700 hr when the timing of nectar production for the latter species started, individuals of A. rutila were seen visiting their flowers, probably consuming the smaller amounts of nectar available at those hours. Hummingbirds are known to visit several species of chiropterophilous flowers during late afternoon, but usually they act as nectar thieves (Sazima et al. 1994, Muchhala 2003).
Sugar concentration
H. guazumifolia: The chemical constituents of floral nectar are known to vary according to the type of pollinator attracted to the flowers (Gottsberger et al. 1989, Baker & Baker 1990, Stiles & Freeman 1993, Baker et al. 1998, Witt et al. 1999), and in turn may affect the visiting behaviour of potential pollinators (Baker & Baker 1976). The three sugars sucrose, glucose, and fructose, are by far the most common and abundant sugars in nectars (Baker et al. 1998) and were contained in the nectar of both Helicteres species. However, nectars differed in the concentration of their sugars (Wells et al. 1992).
Sucrose is the predominant sugar in the nectars of New World hummingbird pollinated species (Stiles & Freeman 1993, Baker et al. 1998) and it is the main sugar in nectar of H. guazumifolia. Sucrose was present more than 200 times as much as glucose and fructose at every sampling time. The sugar concentration of the nectar of some hummingbird pollinated species increased during the morning (Cruden et al. 1983), and the highest sucrose concentration values were recorded during the first hours of the day. Nectar contained a low content of fructose and glucose; however, the hexoses were found to be fructose-glucose balanced, also typical for hummingbird pollinated flowers (Freeman et al. 1985, Stiles & Freeman 1993).
Baker et al. (1998) stated that the low sucrose content in nectar represents the ancestral condition for taxa pollinated by volant vertebrate animals. Most likely, the ancestral condition of nectars with low sucrose content was breached by species in the New World where they were in contact with hummingbirds. The nectar chemistry may have shifted from low sucrose content to high more than once, with the repeated evolution of hummingbird-pollinated species. Adaptive convergence in sugar composition presumably reflects taste preferences of hummingbirds. Nestlings will be imprinted with sucrose when they are fed such a predominantly sucrose diet. Due to the establishment of a long lasting preference for sucrose, hummingbirds may actively seek sucrose-rich food sources (Stiles & Freeman 1993, Baker et al. 1998), and this type of source was highly available at the flowers during the whole period of nectar secretion.
It is expected that secretion of sucrose rich nectar would be more economical than hexose rich nectar, because most sugar is translocated within the plant as sucrose, which is the major sugar in the phloem sap (the source of sugar in nectar) (Stiles & Freeman 1993).
H. baruensis: Flowers pollinated by neotropical bats are seem to be dominated by the hexose sugars, fructose and glucose (Baker & Baker 1990, von Helversen 1993). Although H. baruensis nectar had a similar overall sugar concentration between sucrose, fructose and glucose, it did showed a tendency for it being composed more of fructose and glucose in the majority of time hours of nectar secretion. H. baruensis nectar is one of the principal components in the diet of its main pollinator Glossophaga soricina; as it is the only species that provides nectar during most of the rainy season at Santa Rosa (Fernández Morillo 1998).
Glossophagine bats with weight ranges between 7-35 g are animals with an unusually high energy turn-over. The minimum requirement for a small Glossophaga is in the range of 1 mg of sugar (or about 5 µl of a 20 % nectar) average reward for one flower visit (von Helversen 1993). The glossophagines´ ability to hover leads to the exploitation of the flowers that they visit. Hovering visits of bats generally last less than a second and are of a shorter duration than in hummingbirds. Although hovering is an expensive mode of flight, it allows bats, like hummingbirds, to visit a larger number of flowers per time unit and therefore improves total foraging efficiency (von Helversen 1993, Tschapka & Dressler 2002). Sugars encountered in H. baruensis nectar may tend to compensate the high energetic demands imposed by their flight mode. Also, bats use nectar as an additional water source when water is in short supply under seasonally arid environments like the dry forest (Tschapka & Dressler 2002).
Even though, pollinator visits were not quantified in this study, G. soricina was observed visiting the flowers since 2200 hr until approximately 0300 hr. G. soricina visits flowers of the same bush or group of bushes consecutively every 15 to 40 min during the first half of the night carrying pollen of the plant in its fur (Fernández Morillo 1998, von Helversen & voigt 2002).
Amino acid concentration
The amounts of amino acids present in the nectar of most flowering plants, although small, are sufficient to provide pollinators with a useful nitrogen supply (Baker and Baker 1973a). The overall concentration of amino acids differs among species and appears to be related to their principal pollinator (Gottsberger et al. 1989, 1990; Dress et al. 1997). in general, it has been argued that amino acid concentration is lower if the principal pollinator has alternate sources of amino acids in its diet (Baker 1977, Gottsberger et al. 1984, Dress et al. 1997). Pollinators such as birds and bats, which normally eat pollen or insects, do not need to rely entirely on nectar to obtain all amino acids needed for their nutrition (Gottsberger et al. 1984, 1990). Nectars are typically expected to be richer in amino acids if flowers are pollinated by settling moths, butterflies and many wasps which, as adults, do not have alternative sources of protein building materials (Baker 1977, Gottsberger et al. 1990). This study reveals that there are significant amounts of amino acids in the nectar of these two species, suggesting that their presence may affect the flower-visiting behaviour of potential pollinators (Baker & Baker 1973a, 1976).
H. guazumifolia: it has been shown that hummingbird pollinated flowers often show little amino acid in the nectar (Baker & Baker 1973a, b, Baker 1977, Endress 1994). These results are understandable because hummingbirds can use an alternative source of protein building materials in the insects that they catch (Baker & Baker 1973a, b, Baker 1977). Even though, the nectar of this species contained a total of 17 different amino acids, only four were found to have a concentration above 70 Ng/µl. in contrast, eight amino acids showed concentrations of less than 50 Ng/µl. These findings indicate that H. guazumifolia, as a hummingbird pollinated plant, has nectar with low amino acid content.
Other studies have shown that arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine are essential nutrients for many insects. Proline and glycine are also essential for some insect species. Other amino acids, while not essential, do increase insect growth like alanine, aspartic acid, glutamic acid, glycine and serine (Dress et al. 1997). Several of these essential amino acids for insects, were found in lower concentrations in nectar, e.g., histidine, leucine, lysine, methionine, valine and glycine. isoleucine and phenylalanine were absent in the nectar of H. guazumifolia. in addition, two non essential amino acids for insects, aspartic acid and serine, were present in low concentration in nectar. Lastly, alanine and glutamic acid were absent in the nectar. As a consequence, hummingbird pollinators could actually obtain these amino acids from insects and this could explain why H. guazumifolia´s floral nectar is low in amino acid composition.
H. baruensis: The nectars of bat pollinated plants are low in amino acids content (Baker 1977). Bats need substantial quantities of protein building materials, but they can use pollen, fruits, and insects as alternate sources of nitrogen (Baker & Baker 1973b, Baker 1977, von Helversen 1993). Similar to H. guazumifolia, H. baruensis floral nectar contained 17 different amino acids, but only five were found with a concentration over 20 Ng/µl. Only two amino acids, valine and lysine had concentrations lower than 10 Ng/µl. Ten out of the 17 total amino acids were not present in all samples: glycine, methionine, leucine, histidine, isoleucine, phenylalanine, aspartic acid, glutamic acid, serine and cysteine. These findings indicate that, with the exception of cysteine, the amino acids present in the nectar of this species match the group of essential and non essential amino acids for insects.
Fernández Morillo (1998) determined the feeding behavior of G. soricina during the dry and the rainy seasons at Santa Rosa. During the dry season bats feed predominantly on pollen and nectar, since these resources are in major abundance from late December and May. von Helversen (1993) stated that within the evolution of Glossophaginae pollen has become the main protein source, suggesting that pollen of chiropterophilous flowers can satisfy the amino acids needed by bats (Fernández Morillo 1998). Future studies of H. baruensis are needed to determine amino acid composition of pollen, and compare it with that of its nectar. in this sense it may be expected that some amino acids in pollen grains have higher concentrations, meanwhile in the nectar have lower concentrations or are absent.
During the rainy season bats consumed insects (lepidoptera, hawkmoths, flies and beetles), fruits (Muntingia calabura), pollen and nectar of H. baruensis in similar proportions. The amino acids present in insects can satisfy the requirements of flight, pregnancy and lactancy. Lepidoptera are rich in lipids, which provide more energy than sugars in nectar (Fernández Morillo 1998). H. baruensis has nectar with low amino acid composition, and the amino acids in nectar that were found in low concentration or absent may be obtained directly from insect consumption.
Nectar amino acid complements of closely related species show a high degree of constancy, and they tend to show similar but not identical complements (Baker 1977, Baker & Baker 1979, Cruden & Hermann 1983). Floral nectar of both Helicteres species share four types of amino acids which were the ones found in high concentration: proline, arginine, threonine and tyrosine. They also have two amino acids in common that had lower concentration: lysine and valine. Finally, they also shared three amino acids that were absent from some samples: glutamic acid, isoleucine and phenylalanine. Amino acids can be valuable in taxonomic and phylogenetic studies (Baker 1977, Baker et al. 1998). The results obtained in this study agree with those suggested by Gottsberger et al. (1989), in the sense that within each species certain amino acids predominate while others are absent or appear only in low concentrations.
Floral nectaries morphology: Members of the family Sterculiaceae possess trichomatous floral nectaries, consisting of multicellular clavate hairs, which release nectar from the top and are usually aggregated in cushions or carpets (Endress 1994, vogel 2000). Tricome nectaries seem to be relatively rare (Endress 1994); but it is one of the features characterizing core Malvales (Bombacaceae, Malvaceae, Sterculiaceae and Tiliaceae). in Helicteroids floral nectaries maintain a radial arrangement and carpets are mostly calyx-borne (vogel 2000).
As shown in Figures 5 and 6 each floral nectariferous trichome can be distinguished separately and clearly because they do form a compact carpet, on both Helicteres species. They are always multicellular, uni- to masonry-like pluriseriate, with a basal cell rooting in the epidermis, one neck cell, and a filiform, clavate or fusiform glandular body. All nectar carpets are supplied with special innervation and with additional glandular tissue in the subjacent mesophyll, lending the complex an integrated organ like character (vogel 2000). The glandular tissue is also shown in Figure 6 as a thick layer underneath the layer formed by the trichomatous nectaries.
The dimension of these carpets varies enormously depending on floral and pollinator type (vogel 2000). As flowers of H. baruensis are larger and wider than those of H. guazumifolia, consequently there is more area occupied by the floral nectary of H. baruensis which almost doubles the size compared to the one of H. guazumifolia. Accordingly, this leads to the production of more quantities of nectar by H. baruensis flowers. Nectar glands in bat flowers are often voluminous in comparison to those of related species that are not bat-pollinated, and that is why secretion of nectar is much greater in bat flowers than in all other pollination syndromes (von Helversen 1993).
Acknowledgments
I would like to thank Julio Sánchez and Jorge Leiva for their support and assistance during the field work. i am very grateful to Gerhard Gottsberger and Hans Malchus for their comments and guidance with the laboratory work at University of Ulm, Germany. The Andrew W. Mellon Foundation provided financial support at Santa Rosa National Park.
Resumen
Helicteres guazumifolia Kunth y Helicteres baruensis Jacq. (Sterculiaceae) son dos especies simpátricas de arbustos comunes en el bosque tropical seco de la zona noroeste de Costa Rica. Registré los patrones de producción de néctar de las dos especies según la hora del día o de la noche cuando hubo secreción de néctar. En H. guazumifolia se limitó al primer día del período de vida floral, desde el inicio de la antesis a las 0600 hr hasta las 1800 hr. Las flores secretaron en promedio 15.63 ±8.45 µl (N=409) de néctar. El néctar está compuesto por tres azúcares principales: sacarosa, fructuosa y glucosa, y es rico en sacarosa. Se identificó un total de 17 aminoácidos diferentes en el néctar floral. Prolina, arginina, treonina y tirosina fueron los aminoácidos más abundantes con una concentración mayor a 70 Ng/µl. En contraste, las flores de su pariente H. baruensis, secretaron néctar en el segundo día de vida de la flor, después de la antesis; se inició a las 1600 hr y cesó a las 0600 hr del día siguiente. Las flores secretaron en promedio 77.03 ±64.99 µl (N=163) de néctar. El néctar también está compuesto por tres azúcares principales; no obstante, tiende a ser rico en hexosas, con más fructuosa y glucosa que sacarosa. También contiene 17 aminoácidos libres, siendo los más concentrados prolina, alanina, tirosina, arginina y treonina. Se observan claramente patrones diferentes de producción de néctar entre las dos especies según la hora, la cantidad y la composición del néctar. También describí la morfología de los nectarios florales usando fotomicrografías tomadas con microscopio electrónico.
Palabras clave: Amino acidos, Helicteres guazumifolia, Helicteres baruensis, nectario floral, néctar, polinización, biología reproductiva, azúcares, Costa Rica.
Received 09-X-2007. Corrected 10-VIII-2009. Accepted 04-X-2009.
References
Baker, H.G. & I. Baker. 1973a. Amino acids in nectar and their evolutionary significance. Nature 241: 543-545. [ Links ]
Baker, H.G. & I. Baker. 1973b. Some anthecological aspects of the evolution of nectar producing flowers, particularly amino acid production in nectar, p. 243-263. In v.H. Heywood (ed.). Taxonomy and Ecology. Academic, London and NewYork, USA. [ Links ]
Baker, I. & H.G. Baker. 1976. Analyses of amino acids in flower nectars of hybrids and their parents with phylogenetic implications. New Phytol. 76: 87-98. [ Links ]
Baker, H.G. 1977. Non-sugar chemical constituents of nectar. Apidologie 8: 349-356. [ Links ]
Baker, I. & H.G. Baker. 1979. Chemical constituents of the nectars of two Erythrina species and their hybrid. Ann. Missouri Bot. Gard. 66: 446-450. [ Links ]
Baker, H.G. & I. Baker. 1990. The predictive value of nectar chemistry to the recognition of pollinator types. isr. J. Bot. 39: 157-166. [ Links ]
Baker, H.G., I. Baker & S.A. Hodges. 1998. Sugar composition of nectars and fruits consumed by birds and bats in the tropics and subtropics. Biotropica 30: 559-586. [ Links ]
Bayer, C. & K. Kubitzki. 2003. Flowering plants, dicotyledons: Malvales, Capparales, and non-betalain Caryophyllales, p. 260. In K. Kubitzki (ed.). The families and genera of vascular plants. Berlin, Heidelberg, Germany. [ Links ]
Cristóbal, C.L. 2001a. Taxonomía del género Helicteres (Sterculiaceae). Revisión de las especies americanas. Bonplandia 11: 1-206. [ Links ]
Cristóbal, C.L. 2001b. Sterculiaceae, p. 1911-2666. In W.D. Stevens, C. Ulloa, A. Pool & O.M. Montiel (eds.). Flora de Nicaragua. Monogr. Syst. Bot. Missouri Bot. Gard., Missouri, USA. [ Links ]
Cruden, R.W. 1976. Intraspecific variation in pollen-ovule ratios and nectar secretion-preliminary evidence of ecotypic adaptation. Ann. Mo. Bot. Gard. 63: 277-289. [ Links ]
Cruden, R.W., S.M. Hermann & S. Peterson. 1983. Patterns of nectar production and plant-pollinator coevolution, p. 80-125. In B. Bentley & T. Elias (eds.). The biology of nectars. Columbia, New York, USA. [ Links ]
Cruden, R.W. & S.M. Hermann. 1983. Studying nectar ? Some observations on the art, p. 222-241. In B. Bentley & T. Elias (eds.). The biology of nectars. Columbia, New York, USA. [ Links ]
Dress, W.J., S.J. Newell, A.J. Nastase & J.C. Ford. 1997. Analysis of amino acids in nectar from pitchers of Sarracenia purpurea (Sarraceniaceae). Am. J. Bot. 84: 1701-1706. [ Links ]
Elisens, W.J. & C.E. Freeman. 1988. Floral nectar sugar composition and pollinator type among New World genera in tribe Antirrhineae (Scrophulariaceae). Am. J. Bot. 75: 971-978. [ Links ]
Endress, P.K. 1994. Diversity and evolutionary biology of tropical flowers. Cambridge, Cambridge, USA. [ Links ]
Fernández-Morillo, M.T. 1998. Variación estacional de la dieta y la disponibilidad de alimento de Glossophaga soricina (Chiroptera) en el bosque seco tropical de Santa Rosa. MSc. Thesis, Heredia, Costa Rica. [ Links ]
Freeman, C.E., R.D. Worthington & R.D. Corral. 1985. Some floral nectar-sugar compositions from Durango and Sinaloa, Mexico. Biotropica 17: 309-313. [ Links ]
Freeman, C.E. & D.H. Wilken. 1987. Variation in nectar sugar composition at the intraplant level in Ipomopsis longiflora (Polemoniaceae). Am. J. Bot. 74: 16811689. [ Links ]
Galetto, L. & G. Bernardello. 1995. Characteristics of nectar secretion by Lycium cestroides, L. ciliatum (Solanaceae), and their hybrid. Plant Species Biol. 11: 157-163. [ Links ]
Galleto, L. 1997. Flower structure and nectar chemical composition in three Argentine Apocynaceae. Flora 192: 197-207. [ Links ]
Gentry, A.H. 1993. Field guide to the families and genera of woody plants of North Western South America. Conservation international, USA. [ Links ]
Gerhardt, K. 1993. Tree seedling development in tropical dry abandoned pastures and secondary forest in Costa Rica. J. Veg. Sci. 4: 95-102. [ Links ]
Gottsberger, G., J. Schauen & H.F. Linskens. 1984. Amino acids and sugars in nectar, and their putative evolutionary significance. Plant Syst Evol. 145: 55-77. [ Links ]
Gottsberger, G., T. Arnold & H.F. Linskens. 1989. Intraspecific variation in the amino acid content of floral nectar. Bot. Acta. 102: 141-144. [ Links ]
Gottsberger,G., T. Arnold & H.F. Linskens. 1990. Variation in floral nectar amino acids with aging of flowers, pollen contamination, and flower damage. isr. J. Bot. 39: 167-176. [ Links ]
Hartshorn, G.S. 1991. Plantas, p. 119-353. In D.H. Janzen (ed.). Historia natural de Costa Rica. Universidad de Costa Rica, San José, Costa Rica. [ Links ]
Holdridge, L.R. 1967. Life zone ecology. Tropical Science Center, San José, Costa Rica. 213 p. [ Links ]
Janzen, D.H. 1986. Parque Nacional Guanacaste, restauración ecológica y cultural en el trópico. Servicio de Parques Nacionales, San José, Costa Rica. [ Links ]
Linskens, H.F. & J.F. Jackson. 1987. High Performance Liquid Cromatography in Plant Science. Mod. Meth. Plant Anal., n.s. 5: 1-250. [ Links ]
Martínez del Río, C. 1990. Sugar preferences in hummingbirds: the influence of subtle chemical differences on food choice. Condor 92: 1022-1030. [ Links ]
McDade, L.A. & J. Weeks. 2004. Nectar in hummingbird-pollinated neotropical plants i: Patterns of production and variability in 12 species. Biotropica 36: 196-215. [ Links ]
Muchhala, N. 2003. Exploring the boundary between pollination syndromes: bats and hummingbirds as pollinators of Burmeistera cyclostigmata and B. tenuiflora (Campanulaceae). Oecologia 134: 373-380. [ Links ]
Murcia, C. 2002. Ecología de la polinización, p. 493-530. In M.R. Guariguata & G.H. Kattan (eds.). Ecología y conservación de bosques neotropicales. LUR, Costa Rica. [ Links ]
Perret, M., A. Chautems, R. Spichiger, M. Peixoto & v. Savolainen. 2001. Nectar sugar composition in relation to pollination syndromes in Sinningieae (Gesneriaceae). Ann. Bot. 87: 267-273. [ Links ]
Proctor, M., P. Yeo & A.J. Lack. 1996. The natural history of pollination. Timber, Portland, Oregon, USA. [ Links ]
Robyns, A. & J. Cuatrecasas. 1964. Sterculiaceae, p. 69-107. In R.E. Woodson & R.W. Schery (eds.). Flora of Panama. Ann. Missouri Bot. Gard. 51: 69-107. [ Links ]
Rojas Jiménez, K. 2001. Fenología de la copa y del sistema de raíces finas de Enterolobium cyclocarpum (guanacaste) un árbol de brotación temprana en el bosque tropical seco. MSc. Thesis, San José, Costa Rica. [ Links ]
Sazima, M. & I. Sazima. 1988. Helicteres ovata (Sterculiaceae), pollinated by bats in southeastern Brazil. Bot. Acta. 101: 269-271. [ Links ]
Sazima, M., I. Sazima & S. Buzato. 1994. Nectar by day and night: Siphocampylus sulfureus (Lobeliaceae) pollinated by hummingbirds and bats. Pl. Syst. Evol. 191: 237-246. [ Links ]
Stiles, F.G. & C.E. Freeman. 1993. Patterns in floral nectar characteristics of some bird-visited plant species from Costa Rica. Biotropica 25: 191-205. [ Links ]
Tschapka, M., O. Von Helversen & W. Barthlott. 1999. Bat pollination of Weberocereus tunilla, an epiphytic rain forest cactus with functional flagelliflory. Pl. Biol. 1: 554-559. [ Links ]
Tschapka, M. & S. Dressler. 2002. Chiropterophily: on bat flowers and flower bats. Curtiss Bot. Mag. (ser 6). 19: 114- 125. [ Links ]
Vogel, S. 1990. Radiación adaptativa del síndrome floral en las familias neotropicales. Bol. Acad. Nac. Cien. 59: 5-30. [ Links ]
Vogel, S. 2000. The floral nectaries of Malvaceae sensu lato- a conspectus. Kurtziana 28: 155-171. [ Links ]
Von Helversen, O. 1993. Adaptations of flowers to the pollination by glossophagine bats, p. 167-174. In W. Barthlott, C.M. Naumann, K. Schmidt- Loske & K.L. Schuchmann (eds.). Animal-plant interactions in tropical environments. Bonn Museum Alexander Koenig, Germany. [ Links ]
Von Helversen, O. & C.C. voigt. 2002. Glossophagine bat pollination in Helicteres baruensis (Sterculiaceae). Ecotropica 8: 23-30. [ Links ]
Witt, T., A. Jurgens, R. Geyer & G. Gottsberger. 1999. Nectar dynamics and sugar composition in flowers of Silene and Saponaria species. Caryophyllaceae. Pl. Biol. 1: 334-345. [ Links ]
Wells, H., P.S. Hill & P.H. Well. 1992. Nectarivore foraging ecology: rewards differing in sugar types. Ecol. Entomol. 17: 280-288. [ Links ]
Zimmerman, M. 1988. Nectar production, flowering phenology, and strategies for pollination, p. 157-178. In J.L. Doust & L.L. Doust (eds.). Plant reproductive ecology, patterns and strategies. Oxford, New York, USA. [ Links ]

