










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 n.4 San José Dec. 2009
The effect of parasitism on the population dynamics of the macadamia nutborer Gymnandrosoma aurantianum (Lepidoptera: Tortricidae)
Helga Blanco-Metzler1*; Allan D. Watt2 & Derek Cosens3
1. Crop Protection Research Centre (CIPROC), University of Costa Rica. San José, Costa Rica; helgablanco@gmail.com
* Correspondence author
2. Centre for Ecology and Hydrology, Edinburgh, Scotland.
3. Zoology Department, Edinburgh University, Scotland.
Abstract: Biological control on crop infesting insects represent an useful method in modern agriculture. A search for parasitoids of the macadamia nutborer was carried out during a three year study, and their effect on the population fluctuation of the borer was determined. An egg parasitoid belonging to the family Trichogrammatidae and four larval parasitoids, Microgastrine I, Microgastrine II, Ascogaster sp. (Hymeoptera: Braconidae) and Pristomerus sp. (Hymeoptera: Ichneumonidae) were recovered. Parasitism percentage by Microgastrine I was 15% in 1991, 16% in 1992 and 4% in 1993; Microgastrine II was not collected in 1991, but accounted for a 4.3% of parasitism in 1992 and 3.7% in 1993; Ascogaster sp. was registered since 1992 with 3% parasitism (29% in 1993). We found an inverse relationship between total parasitism and the mean of damaged nuts. Parasitoids play an important role in the reduction of the G. aurantianum population. Rev. Biol. Trop. 57 (4): 1245-1252. Epub 2009 December 01.
Key words: Gymnandrosoma aurantianum (Ecdytolopha torticornis), biological control, parasitoids, macadamia, Costa Rica.
Nut trees of the genus Macadamia spp. (Proteaceae) grow naturally in rain forests of Australia and New South Wales (Ironside 1992). Macadamia trees have been introduced and planted in many parts of the world, particularly in Hawaii, South Africa, Malawi, Guatemala and Costa Rica.
The macadamia was introduced to Costa Rica in 1952, but it was not until 1965 that it started to be grown commercially as an alternative crop to coffee. The orchards were situated in plantations previously devoted to coffee (50- 1200). Macadamia exceeded all expectations in productivity. New orchards started to proliferate and soon it became a highly remunerative, non traditional export crop. At the present time, Costa Rica (2100 MT) is the world’s fourth largest producer of macadamia, behind Australia (33000 MT), United States (24494 MT), South Africa (11500 MT), Guatemala (9360 MT) and Kenya (5800 MT) (USDA 2006).
With the excellent ecological conditions that Costa Rica offers for nut production, and the attractiveness of its international price, various government agencies as well as private banks have adopted generous incentive schemes to stimulate macadamia nut production. As the area used to grow macadamia increased, so have reports by producers of pest damage. One of the causes of damage to the nuts is Gymnandrosoma aurantianum (former Ecdytolopha torticornis) (Meyrick) (Lepidoptera: Tortricidae) which tunnels into the nut and feeds on the kernel. The life cycle from egg to adult of the nutborer lasts nearly 36 days (Blanco-Metzler et al. 1993), so it is possible it could produce up to ten generations per year depending on the environmental conditions and food availability.
Appropriate pest management usually consists of a combination of two or more techniques (e.g. introduction or augmentation of biological control agents, the development of sampling methods, the correct timing and dosage of insecticides) which diminish the pest population without producing problems in the environment (Horn 1988). As G. aurantianum is relatively new to damaging macadamia nuts, there is a need to identify the natural control agents and their relationship to its host to devise management strategies for this pest.
There are nine species of macadamia: Macadamia integrifolia, M. tetraphylla, M. ternifolia, M. claudiensis, M. grandis, M. hildebrandii, M. jansenii, M. whelanii, M. neurophylla; of these species, only integrifolia and tetraphylla have commercial value. Present commercial varieties are mainly grafted Hawaiian selections of M. integrifolia. The tetraphylla types are inferior in nut quality and are not accepted by processors, and the remaining types have a small and bitter nut (MAG 1991). Of the many Hawaiian varieties developed, the most commonly grown in Costa Rica are clones Keahou (Haes 246), Kau (Haes 344), Kakea (Haes 508) and Keaau (Haes 660).
The aim of this study was to determine the presence of G. aurantianum egg and larval parasitoids, to record population fluctuations in parasitoids, and to quantify the suppression of the nutborer population achieved with the use of parasitoids.
Materials and methods
Parasitism was studied in forty damaged nuts per clone (246, 344, 508, 660) for a total of 160 nuts, harvested at random from the trees each month, at Oriente Farm, Turrialba, Costa Rica, 09°53’ N, 83°41’ W, 620 to 700m a.s.l. The mean annual rainfall during the period 1991–1993 was 2600mm; the mean temperature 22°C, and the relative humidity around 80%. Nuts were examined in the laboratory (23°C, 80% RH) at CATIE (Centre for Tropical Agricultural Research and Education), Turrialba, Costa Rica, and the following variables were recorded: shell diameter, husk diameter, number of holes per nut, location of the larvae in the nut, number of larvae and their instar, and the number and species of parasitoids. From April 1992, sample size was increased to 15 damaged nuts per clone per site (five sites) for a total of 300 nuts examined per month. The host larvae found were incubated individually in split nuts in 165cm3 plastic cups until emergence of either the adult moth or a parasitoid. Large holes in the lids were lined with muslin to provide ventilation. Nuts were tied up with tape and moistened absorbent filter paper was added to the bottom of each cup in order to reduce desiccation. Macadamia nut clone 344 was used as the food source, as this line was found to be the most susceptible to G. aurantianum (Blanco-Metzler 1994), and it was changed every two days.
Apparent percentage parasitism was estimated from the total number of parasitoids reared in relation to the total number of hosts either parasitized or successfully reared to the adult stage. The following formula was used:
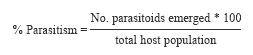
Differences in the parasitism rates were determined by Chi-square analysis for the two most abundant parasitoids and for differences in parasitism rates in the different macadamia clones. The accepted level of significance was P<0.05 in all instances.
Results
Six species of hymenopteran parasitoids were recovered from the nutborer for the first time (Table 1). An egg parasitoid belonging to the family Trichogrammatidae was recovered in 1992. Nuts containing fertile nutborer eggs collected from the field turned black in the laboratory, and a mean of 3.33 (SE=0.68; n=18) wasps per egg were observed. In view of its low occurrence, and the difficulties in finding unhatched nutborer eggs in the field, more emphasis was given to the study of larval parasitoids.
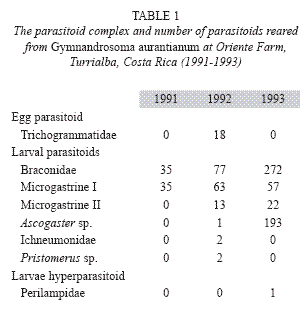
A total of three braconids, and one ichneumonid species were reared from the nutborer larvae between 1991 and 1993. Microgastrine I occurred each year, while Microgastrine II and Ascogaster sp. occurred only during the latter years. The Microgastrine species are named as I and II since specialists (see acknowledgements) were not able to identify them to the species level and biomolecular methods were not yet available making misidentifications possible. The difference between these two species is based on the colour of the legs and their size: Microgastrine I has brown legs and is larger, while Microgastrine II has black legs. Microgastrine I was the dominant parasitoid in terms of consistency. The larval parasitoid complex was dominated by Microgastrine I during the first two years and by Ascogaster during the final year. Microgastrine I was reared on 35 occasions in 1991, 63 in 1992 and 57 in 1993; Microgastrine II was reared 13 times in 1992 and 22 in 1993. The number of Ascogastger recorded was highest among all the parasitoid species followed by Microgastrine I and Microgastrine II which occurred sporadically, and in small numbers (Fig. 1, Table 1). There was a significant difference between the number of Microgastrine I, and Ascogaster sp. parasitoids reared (X2=4.36; df=1; P<0.05), (Chi-square analysis was only calculated for these two parasitoids due to the very low abundance of the other species). Pristomerus was reared only in two occasions during 1992. Although four parasitoids were reared from the nutborer larvae, multiple parasitism was not observed.
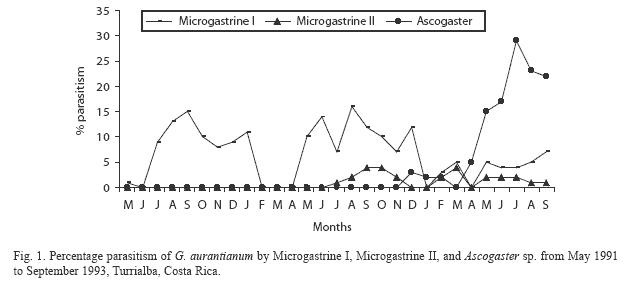
The percentage parasitism in each year of sampling is presented in Fig. 1. The number of nutborer larvae parasitized by Microgastrine I was 15% in 1991, 16% in 1992 and 4% in 1993; parasitism by Microgastrine II reached 4.3% in 1992 and 3.7% in 1993, while the level of parasitism by Ascogaster was 3% parasitism in 1992 and 29% in 1993.
A larval hyperparasitoid belonging to the family Perilampidae was found in 1993.
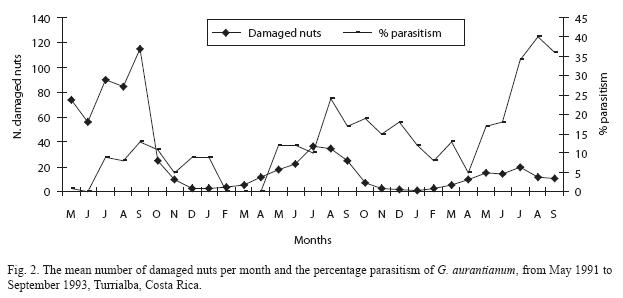
Total parasitism was related to the mean number of damaged nuts per month: there was an inverse relationship between the number of damaged nuts and percentage parasitism (Fig. 2). There were three peaks with highest parasitism: September 1991, August 1992 and August 1993. These peaks followed the months when nut damage was highest, except in 1991 where it coincided with the peak number of damaged nuts. However, even in 1991 there was evidence of parasitism lagging slightly behind nutborer damage because parasitism remained at a high level while nutborer damage declined. The parasitoid population peaked after nut damage reached its highest value, indicating a delayed density-dependent response of the parasitoid population to that of G. aurantianum. Percentage parasitism increased in 1992 and 1993, being highest at the end of the third year. However, parasitoids were not reared on every sampling date during the first two years, but were reared on every occasion during the third year of study, so for the early years percentage parasitism may be underestimated.
The percentage parasitism per clone is presented in Figs. 3 to 6. The monthly estimates of percentage parasitism were highest among larvae from clone 660 reaching a maximum of 33% in 1991, and 71% for the same clone in 1992; percentage parasitism was highest in clone 246 in 1993 (80%). Percentage parasitism for individual clones had a similar pattern to that of damaged nuts and total parasitism (Fig. 2). There was a highly significant difference among the number of parasitoids reared per clone, being highest in clone 246 (129 parasitoids), followed by clone 660 (97 parasitoids), 344 (96 parasitoids), and 508 (64 parasitoids) (X2=21.89; df=3; P<0.001). However, no significant difference was found for overall parasitism (X2=2.76; df=3; P=0.53). The overall parasitism was highest in clone 246 (19.9%, 649 larvae), followed by clone 660 (15.5%, 624 larvae, clone 344 13.1%, 732 larvae, and clone 508 (11.3% 565 larvae). Parasitism might have been higher than estimated since some of the small larvae died during rearing.
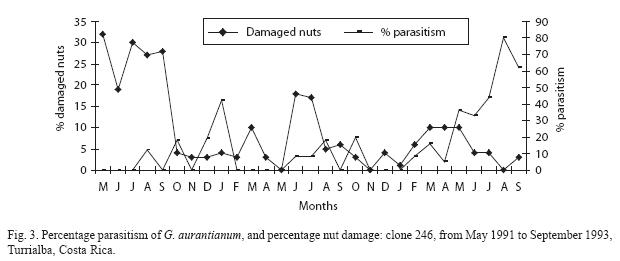
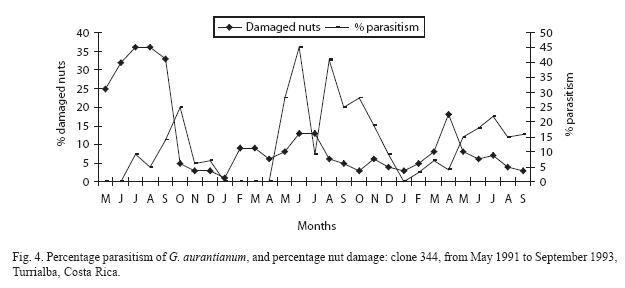
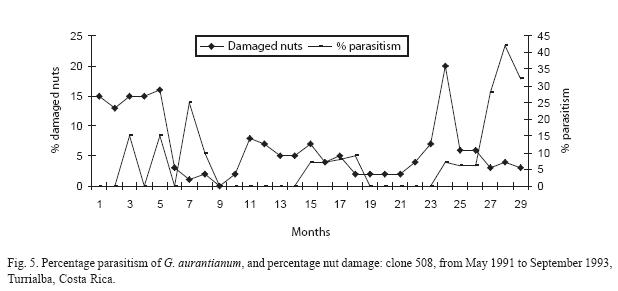
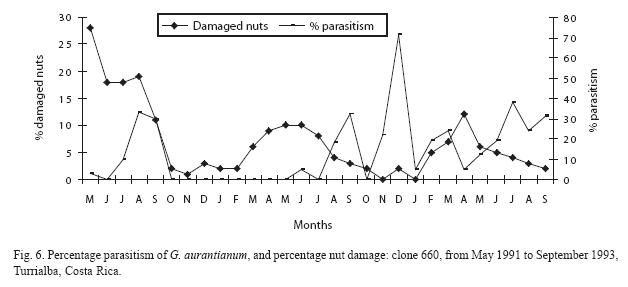
Discussion
The fact that six hymenopteran parasitoids were reared from the nutborer in the macadamia orchard, although only two reached high levels of parasitism, supports the view that G. aurantianum is likely to be an indigenous species which switched to a more abundant food source (Blanco-Metzler et al. 1993). However, it is not known whether the parasitoids are generalists or specialists on the nutborer, since they could not be identified to the species level. If the nutborer is an exotic species, there would not have been enough time for the host-parasitoid relationship to develop since macadamia is a relatively new introduction into Costa Rica (unless both the nutborer and its parasitoids were simultaneously introduced). Furthermore, the parasitoid genera collected in this study have also been reported in Cryptophlebia studies (La Croix & Thindwa 1988, van der Geest et al. 1992).
A similarity in population trends between the nutborer and its parasitoids is evident: as the nutborer population increased the parasitoid population also increased, but with a time lag strongly suggesting a delayed densitydependent response. This also indicates that the nutborer population may be regulated to some degree by natural enemies. Further evidence for this comes from the inverse relationship between nut damage and parasitism between years: parasitism increased during the second and third year, while nut damage decreased (Fig. 2). However, a longer study of nutborer parasitism is needed as the situation described here could change with time due to external factors affecting either population. For example, Münster-Swendsen (1980) found strong spatial density-dependent parasitism of lepidopteran larvae by the braconid Apanteles tedellae, but over a period of nine years, the abundance of adult parasitoids was unrelated to that of their hosts in the same year.
Although parasitoids are widely thought to cause spatially density-dependent mortality, Hassell (1978) reports that this pattern is clearly manifested in only a minority of instances. Strong et al. (1984) concluded that for herbivorous insects, natural enemies are of overriding importance as agents of densitydependent control and that intra-specific competition for food is, in contrast, quite rare. The opposite conclusion was reached by Dempster (1983) in reviewing natural control of lepidopteran populations because he found more evidence of density-dependence due to intraspecific competition than to natural enemies. In addition, Stiling (1988) found that out of 32 cases in which density-dependence has been demonstrated, in 19 cases (59.4%) it was due to factors acting from the trophic level below (the plant), whereas in only 13 (40.6%) was densitydependence found to be acting from the trophic level above (predators and parasitoids).
Microgastrine I can be considered to be an effective nutborer parasitoid because it was present throughout each of the three years of study, and also because the achieved level of parasitism was relatively high. The possible reasons why Ascogaster was only found at the end of the second year, and why it became the dominant parasitoid, thus suppressing Microgastrine I, are not clear. However, the aggressiveness of this parasitoid makes it an important biological control agent. Therefore, this should be taken into consideration when designing a pest management programme.
Due to the infrequent occurrence of Pristomerus and the hyperparasitoid, it is likely that they were transients rather than permanent inhabitants of the crop, and that the original hosts of these two parasitoids were probably other lepidopteran larvae from the surrounding forest. Since the Perilampidae is a hyperparasitoid its contribution to natural control must be considered negative.
There are various reasons which can explain the variability observed in the levels of parasitism: the death of small larvae during rearing which might have been parasitized, thus underestimating the percentage parasitism; the distribution of the nutborer and its parasitoids across the orchard and within the tree, relative to the sampling procedure used; and, finally, the frequency of sampling. A more regular sampling programme, for example at fortnight intervals, could have avoided or reduced some of the variability observed.
Although no differences were found between clones with respect to overall parasitism, the data suggest some pattern in parasitism of the nutborer. It was particularly high in the middle part of the study in clone 344 and towards the end of the study in clone 246. Apart from one sampling date, parasitism of larvae in clone 660 was relatively consistent and parasitism in clone 508 was lower than that found in other clones. It must be stressed that these are tentative interpretations and that further studies of parasitism are required. There is no clear explanation why clone 660 had the highest parasitism in two of the three years of study. It could have been an artefact due to the relatively high number of parasitoids reared from a small number of nutborer larvae (71% parasitism in 1992 from seven larvae). Van Driesche (1983) and van Drieshe & Bellows (1996) criticizes the use of peak percent parasitism such as this, as a statistic for comparing or assessing parasitoid impact on pest regulation. He considers that percentage parasitism is often of little or no biological significance, since depending on the relative phenological processes of the host stage and the parasitoid, peak percentage parasitism can be higher, lower, or the same as the generational percentage parasitism value. Nevertheless, it is more important to study the way that parasitism varies, than simply to record overall levels of parasitism, because it is the way that parasitism responds to host abundance that determines its ability to play a role in regulating its abundance.
The existence of nutborer parasitoids is important in planning pest management strategies. The presence of high larval nutborer populations is an indicator that egg parasitoids are inadequate in suppressing borer populations. Because of the importance of Trichogrammatidae in reducing the nutborer population from the egg stage, efforts should be made to increase the population of this parasitoid. A similar conclusion was reached by Ironside (1992) who strongly recommended the introduction of Trichogrammatidae to other macadamia growing areas, because of its strong affinity to macadamia in Malawi, and for their distribution at all heights within the tree. The benefits of mass releasing Trichogramma for the control of the codling moth Cydia pomonella and the summer fruit tortrix moth, Adoxophyes orana in apple orchards was reported by Hassan et al. (1988). They found that T. dendrolini reduced the damage of C. pomonella by 61% and of A. arena by 73%. Generally, a reduction of 67% in the number of damaged fruits is considered economic.
Acknowledgments
Thanks to Paul Hanson for the identification of parasitoids. This research was financed by the British Council and the Tropical Agricultural Research and Training Centre, CATIE.
Resumen
Durante tres años se realizó una búsqueda de los enemigos naturales del barrenador de la nuez de macadamia y se determinó el efecto sobre su población. Se registró un parasitoide de huevo perteneciente a la familia Trichogrammatidae y cuatro parasitoides de larva, Microgastrine I, Microgastrine II, y Ascogaster sp. (Hym: Braconidae) y Pristomerus (Hym: Ichneumonidae). El porcentaje de parasitismo por Microgastrine I fue del 15% en 1991, 16% en 1992 y 4% en 1993; Microgastrine II no se registró en 1991, pero alcanzó un 4.3% de parasitismo en 1992 y un 3.7% en 1993; Ascogaster sp. se registró a partir de 1992 con un 3% de parasitismo y con un 29% en 1993. Se encontró una relación inversa entre el parasitismo total y el promedio de nueces dañadas. Los resultados indican que los parasitoides juegan un papel importante en la disminución de la población de G. aurantianum.
Palabras clave: Gymnandrosoma aurantianum (Ecdytolopha torticornis), control biológico, parasitoides, macadamia, Costa Rica.
Received 15-X-2008. Corrected 25-V-2009. Accepted 26-VI-2009.
References
Blanco-Metzler, H., A.D. Watt & D. Cosens. 1993. Ciclo de vida y comportamiento de oviposición de Ecdytolopha torticornis (Lepidoptera: Tortricidae) barrenador de la nuez de macadamia. Revista Manejo Integrado de Plagas No. 29: 36-39. [ Links ]
Blanco-Metzler, h. 1994. The biology and ecology of the macadamia nutborer Ecdytolopha torticornis in Costa Rica . PhD Thesis, University of Edinburgh, Scotland. [ Links ]
Dempster, J.P. 1983. The natural control of populations of butterflies and moths. Biol. Rev. 58: 461-481. [ Links ]
Hassan, S.A., E. Kohler & W.M Rost. 1988. Mass production and utilization of Trichogramma: 10. Control of the codling moth, Cydia pomonella and the summer fruit tortrix moth, Adoxophyes orana (Lep.: Tortricidae). Entomophaga 33: 413-420. [ Links ]
Hassell, M.P. 1978. The dynamics of arthropod predatorprey systems. Monographs in Population Biology 13. Princeton University, New Jersey, USA. [ Links ]
Horn, D.J. 1988. Ecological approach to pest management. Guilford, New York, USA. [ Links ]
Ironside, D.A. 1992. Suppression of nutborer (Cryptophlebia batrachopha Meyrick and C. leucotreta Meyrick, Tortricidae) on macadamia in Malawi by Trichogramma cryptophlebiae, p. 38-40. In H.C. Bittenbender (Ed). Proceedings of the First International Macadamia Research Conference, July 28-30, 1992. Honolulu, University of Hawaii at Manoa, Hawaii, USA. [ Links ]
La Croix, E.A.S. & H.Z Thindwa. 1988. Macadamia pests in Malawi. III. The major pest. The biology of bugs and borers. Trop. Pest Manag. 32:11-20. [ Links ]
Ministerio de Agricultura y Ganadería (MAG). 1991. Macadamia: Macadamia integrifolia Proteaceae. Comisión Nacional de Investigación y Transferencia de Tecnología Agropecuaria, CONITA. Serie ITTA N. 8, Universidad Estatal a Distancia. San Jose, Costa Rica. [ Links ]
Münster-Swendsen, M. 1980. The distribution in time and space of parasitism in Epinotia tedella (Cl) (Lep: Tortricidae). J. Anim. Ecol. 5:373-383. [ Links ]
Stiling, P.D. 1988. The frequency of density-dependence in insect host parasitoid systems. Ecology 68:844-856. [ Links ]
Strong, D.R., J.H Lawton & R. Southwood. 1984. Insects on plants: community patterns and mechanisms. Blackwell, Oxford, England. [ Links ]
van der Geest, L.P.S., C.H Wearing & J.S Dugladale. 1991. Tortricids in miscellaneous crops, p. 563-577. In L.P.S. van der Geest & H.H Evenhuis (Eds). Tortricid pests: their biology, natural enemies and control. Elsevier, Amsterdam, Holland. [ Links ]
van Driesche, R.G. 1983. The meaning of "percent parasitism" in studies of insect parasitoids. Environ. Entomol. 12:1611-1622. [ Links ]
van Driesche, R.G. & T.S. Bellows. 1996. Biological Control. Chapman & Hall. New York, New York, USA. [ Links ]
Internet references
United States Department of Agriculture. 2006. Macadamia, Inshell Basis- Production, Consumption, Exports, and Imports Statistics. (Downloaded May 16 2009, http://indexmundi.com/en/commodities/agricultural/macadamia-inshell-basis/) [ Links ]

