










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 n.3 San José Sep. 2009
Forum
Conceptual and statistical problems associated with the use of diversity indices in ecology
Gilbert Barrantes & Luis Sandoval
Escuela de Biología, Universidad de Costa Rica, Ciudad Universitaria Rodrigo Facio, San José, Costa Rica. Fax: 2207-4216; gilbert.barrantes@gmail.com
Abstract: Diversity indices, particularly the Shannon-Wiener index, have extensively been used in analyzing patterns of diversity at different geographic and ecological scales. These indices have serious conceptual and statistical problems which make comparisons of species richness or species abundances across communities nearly impossible. There is often no a single statistical method that retains all information needed to answer even a simple question. However, multivariate analyses could be used instead of diversity indices, such as cluster analyses or multiple regressions. More complex multivariate analyses, such as Canonical Correspondence Analysis, provide very valuable information on environmental variables associated to the presence and abundance of the species in a community. in addition, particular hypotheses associated to changes in species richness across localities, or change in abundance of one, or a group of species can be tested using univariate, bivariate, and/or rarefaction statistical tests. The rarefaction method has proved to be robust to standardize all samples to a common size. Even the simplest method as reporting the number of species per taxonomic category possibly provides more information than a diversity index value. Rev. Biol. Trop. 57 (3): 451-460. Epub 2009 September 30.
Key words: diversity indices, multivariate analyses, species richness, species abundance, highland avifauna.
Patterns of species diversity are the result result of historical, evolutionary, and ecological processes combined, which influence in shaping such pattern vary across geographical regions, and temporally within each region (Mac Arthur 1961, 1972, May 1975, Ricklefs and Schluter 1993, Monge-Nájera 1994, Rosenzweig 1995). Hence, understanding the processes responsible for diversity patterns at different geographical scales has been a central issue in ecology and biogeography (Mac Arthur and Mac Arthur 1961, Mac Arthur et al. 1962, Lloyd and Ghelardi 1964, Shapiro 1975, Cody 1993). A necessary process in understanding the causes shaping patters of diversity is to measure diversity at different scales (Whittaker 1972): gamma, beta, and alpha diversity. Community ecologists, who perceived diversity as the of competition and other ecological interactions, have primarily focused at local patterns of diversity (Hutchinson 1959, Mac Arthur and Mac Arthur 1961, Mac Arthur et al. 1962, Schoener 1974, May 1975, Cody 1993, Singh et al. 2005). Community ecologists implemented numerous methods that incorporated not only the number of species but each species abundance to measure alpha diversity (Magurran 1988). These methods have been widely used, though; most of them have severe conceptual and statistical flaws (Wiens 1989, Gotelli and Graves 1996).
The publication of papers using diversity indices to analyze alpha diversity has fluctu ated over time. A few decades ago, community ecologists used diversity indices in nearly any paper that deal with alpha diversity. At present and despite the serious drawbacks inherent to many of the diversity indices, as pointed out for several authors (Peet 1975, Green 1979, Gotelli and Graves 1996), these indices continue appearing in a fairly large number of papers (García Novo 2003, Chao and Shen 2003, Singh et al. 2004, Pinkus Rendón et al. 2006, Castro-Franco et al. 2006, Rawlinson et al. 2005, Forster et al. 2006), texts of general ecology (Begon et al. 2006, Cain et al. 2008), and are regularly taught in class rooms worldwide.
The goal of this paper is to highlight and discuss some of the problems inherent to the indices of measuring alpha diversity, particularly Shannon-Wiener possibly the most widely used diversity index. Some of these problems have already been pointed out and discussed in some extent by Peet (1975), Green (1979), Washington (1984) and Gotelli and Graves (1996). First, we provide a general review and discussion of the: (1) conceptual problems, (2) problems associated to examine each species ecological performance, and (3) statistical problems. Second, we analyze a data set using the Shannon-Wiener index and some other alternative statistical techniques. Finally we discuss the disadvantages of using the Shannon-Wiener index over other statistical methods.
Conceptual issues: Most indices used to measure α diversity (e.g., Shannon-Wiener, Brillouin, Morisita) combine two unrelated aspects: species composition and abundance of species. The species composition in a given area is mostly explained by historical factors such as dispersal events, geographical isolation, and extinction due to past climatic and geological events (MacArthur and Wilson 1967, Haffer 1974, Prance 1987, Roughgarden and Pacala 1989, Humphries and Parenti 1999, Wiens 1989, Hubbell 2001, Lourenço 2003, viloria 2003), and in much less extend by some ecological factors such as competition and predation (Mayr 1963, Grant 1986, Simpson 1953, Schluter 2000). On the other hand, abundance or density of individuals per species in a given area results from the combination of each species life history traits (e.g. reproductive rate), with intra-and interspecific interactions. Predation, parasitism, and competition are density-dependent factors that play an important role in regulating the abundance of populations within a community (Roughgarden and Diamond 1986, Wiens 1989, Howe and Westley 1988). There are well documented examples on the regulatory effects of predation, parasitism, and intra- and inter-specific competition on population size (Robertson 1996). However, these factors rarely are the cause of local or total species extinctions.
Diversity indices combine the number of species (or species richness), an attribute of the community related to historical factors, and species abundance which is an attribute of the population determined by the reproductive potential and survival rates of each species, and population regulatory interactions (Andrewartha and Birch 1954, Hubbell 2001). The application of an index, such as the Shannon-Wiener index, results in a value of difficult interpretation that has little or no meaning by itself. Furthermore, when diversity indices are applied to a set of data (i.e., a group of species and their abundances) the species lose their "identity", therefore, little can be inferred of their role in the community, and even less can be inferred on the evolutionary or biogeographical patterns of the species involved (e.g. Losos and Miles 1994, Roig-Juñent and Flores 2001). The reduction in the information of species richness and abundances is comparable to the sacrificial pseudoreplication pointed out by Hurlbert (1984). in both cases, application of indices of diversity and sacrificial pseudoreplication, a great effort is made to gather good quality information (or a good sampling) of each species and then nearly all the information is lost when all species are combined and their identity are lost.
The combination of species richness and abundance has confounded the definition of diversity. Thus, it is common that diversity refers exclusively to species richness or to the combination of species richness and their abundance – evenness. Fortunately, several authors have in last years focused on species richness to address studies of diversity. Consequently, some interesting ecological and evolutionary patterns on species distributions have emerged at a local and at a much larger geographical scales (e.g., Ricklefs and Schluter 1993, Rosenzweig 1995, Sax and Gaines 2003, Mac Nally et al. 2004).
Ecological function of species: Studies on species functional ecology have become increasingly common in last years. This concept has often been treated without enough rigorously in the sense that species are considered equal and therefore substitutable for other species with similar function (Peterson et al. 1998, Statzner and Moss 2004, Sekercioglu 2006). it is well known that there is not a single pair of species (even sister species) that feed on the same items or use exactly the same habitat (Bouton et al. 1999, Schluter 1993). Although, it is also true that the functional role of species that feed upon similar food resources in similar habitats partially overlap. For example, different species of hummingbirds or fruit-eating birds that feed on nectar or fruits of the same plant species in different communities may well serve as pollinators and dispersers in both communities (Stiles 1985, Moermond and Denslow 1985). However, comparisons across communities using diversity indices prevent revealing and/or inferring the "ecological role" or functional performance of each species in the community, and it is at species level (or lower levels: population and individuals) that comparisons of the functional role is useful in a community.
Some authors have used the diversity indices to compare the diversity between or among particular taxonomic or ecological groups of species (e.g., the so called guilds) (Bethke and Nudds 1993), but focusing on a particular taxonomic or ecological group does not prevent the problem of combining species richness and species abundances in a single value. An additional problem was discussed by Peet (1975), Krebs (1989), and Gotelli and Graves (1996). These authors pointed out that most diversity indices do not recognize intraspecific differences among individuals of a particular species. individuals of different ages or different sexes could use different resources (e.g. food) or different habitats in the community (Polis et al. 1989). including different age classes and sexes in the same "bag" result in lost of information, particularly for studies of the functional role of the organisms in the community. However, considering such categories as different species, as suggested by Hendrickson and Ehrlich (1971), overestimate the species richness in the community, and consequently affect the calculated value of the diversity index (Gotelli and Graves 1996).
Statistical problems: Several authors have discussed in some extend the statistical problems associated to indices for measuring diversity (see Peet 1975, Washington 1984, Gotelli and Graves 1996 for reviews). Thus, we will not go through all the details, but instead will outline some of these problems here.
1. Heterogeneity in sampling units. in an attempt to correlate species diversity with performance of species in the community, some authors have used biomass, cover, or productivity instead of species as sampling units in measuring diversity (Dickman 1968). Using different sampling units, other than species, makes it impossible comparisons of diversity among different study sites.
2. Diversity indices are very sensitive to sample size. Nearly all indices used to measure diversity include number of species (S) as one of their terms. Hence, they are sample size (S) dependent (Peet 1975, Green 1979), and populations with different sample size affect greatly the result.
3. Diversity indices lack a probabilistic basis. This occurs because it is impossible to know the expected distribution of indices, making it impossible to assign a probability value to a given index value (Ghent 1991), and to evaluate differences in diversity values between communities (Gotelli and Graves 1996).
Jacknife technique has been used to produce confidence intervals around a value of a diversity index (Adams and McCune 1979). However, as it has been clearly stated by Gotelli and Graves (1996), the repeated sampling is not sufficient to eliminate the effect of the high sensitivity of most diversity indices to the number of species and individuals in the sample. Peet (1975) proposed to re-scale algebraically the index by bounded it from 0.0 to 1.0. However, this procedure neither removes the sampling bias caused by difference in sample size (Gotelli and Graves 1996).
Materials and methods
Study case: We use the Shannon-Wiener index and other alternative methods on a simple data set. The data consist of census of resident birds on six different localities in the Costa Rica mountain ranges: villa Mills, Reserva Biológica Cerro de la Muerte, volcán irazú, volcán Barva, volcán Poas, and Monteverde. Four censuses were conducted along 2 km transects in each locality from 05:45 to 09:00 h. Our goal is to describe the patterns of species richness and species abundances of Costa Rican highland birds. Additionally, we test whether species richness decreases with elevation or habitat available across localities.
Focal species: The Costa Rican highland bird species were defined by Wolf (1976) as those species that occur at the highest elevations of the four mountain ranges: Talamanca, Central, Tilarán, and Guanacaste. Nearly 40% of these species are endemic to the high mountains of Costa Rica and western Panama (Slud 1964, Barrantes, in press) where they inhabit the paramo, elfin forest, and high mountain forest. The area that these vegetation types covers on the different mountain ranges decreases from the Talamanca mountain range (the southern most mountain range) to Guanacaste mountain range (the northern most mountain range), following their reduction in elevation (Barrantes and Loiselle 2002).
Statistical analyses: We used the Shannon-Wiener index (despite the numerous statistical flaws associated to the index) to compare the diversity (combination of species richness and their abundances) of highland bird species among communities or localities. We also calculated the Jackard index and Morisita index to compare the species composition and abundance across communities and used Cluster analyses to represent the pattern of species richness and species abundances across localities. Correlations (Spearman correlations) were used to evaluate the associations between (a) values of diversity and species richness, (b) species richness and elevation (Barrantes, in press), and area of habitat available (Barrantes and Loiselle 2000) across localities. A Principal Component Analysis was used to graphically display the relation among localities based on the number of species and their abundances.
Results
The species diversity (H) for all species varied across localities (Fig. 1). The highest values of diversity corresponded to Volcán Irazú and Monteverde, and the lowest value to Cerro de la Muerte indicating that the diversity was high and quite similar between volcán irazú and Monteverde. However, the difference in species composition was nearly the most extreme between these two localities (Fig. 2). Monteverde had the second highest number of species but shared less of these species with other localities (Fig. 2). On the contrary, two pairs of localities, Irazú-Barva and Poás-Cerro de la Muerte shared a relatively high number of species (52% in both cases), though the irazú had 56 species and Barva only 35; Poás and Cerro de la Muerte had a similar number of species (Fig. 2). No associations were detected between species richness and: diversity values, area of potential habitat available, and elevation across localities (Spearman correlations, p>0.20 in all cases).

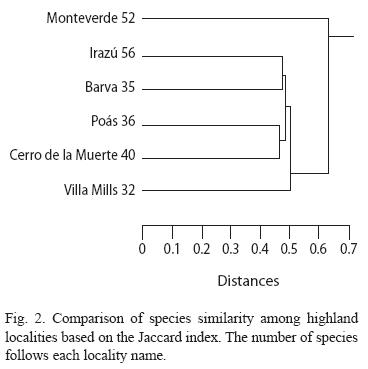
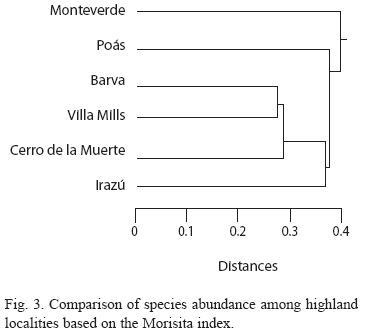
The pattern of abundance (using the Morisita) across localities showed more concordance with the pattern of diversity (Fig. 1 and 3). Monteverde showed the highest difference in species abundance when compared with other localities, while Barva and Villa Mills were the most similar localities (Fig. 3). Other localities shared intermediate number of species and species abundance. The abundance analysis indicated that either a relatively small number of species were common between localities or the abundance of the same species largely differ between localities (if a pair of localities share the same number of species and these species have the same abundance the distance of the branch linking these two localities is 0). In this case a relatively low number of species is shared between localities (Fig. 3) and species shared between localities had different abundances (data not shown).
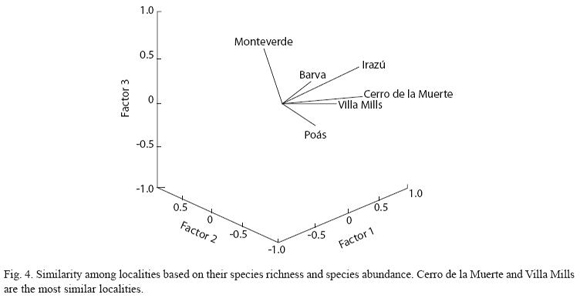
The pattern of similarity in species richness and species abundances across localities is perhaps better visualized in the Fig. 4, based on PCA. it displays the position of the localities into a species richness-species abundance multidimensional space. Thus, localities grouping close together possess similar combination of richness and species abundances. For example, Cerro de la Muerte and villa Mills were the most similar localities based in the combination of composition and abundance of the highland bird species, on the contrary, Monteverde and irazú differ the most with other localities.
Discussion
The Shannon-Wiener diversity index performed poorly in retaining information on species richness and species abundance from a given community, making comparisons of species richness and their abundances across highland localities difficult if not impossible; similar results were also found by Yue et al. (2007) in other study cases. information on species composition and abundance is critical since knowing the species present in a given locality is necessary to understand causes of species geographical distribution, function of species in a community, and even more critical for conservation of species (Primack 1998). Each species is a unique evolutionary entity and there are no a single pair of species that have the same life history traits, the same distribution, or play identical role in a community (Schluter and Ricklefs 1993). However, in some cases as in plant pollination or seed dispersal different bird species may accomplish similar functions (Stiles 1985, Moermond and Denslow 1985). This type of interspecific interactions and the evaluation of similar roles played by different species in different communities only are possible if information on species composition and their abundances is retained by the statistical methods used.
There is often no a single statistical method that retain all information needed to answer even a simple question, such as uncovering the possible causes that determine the pattern of species composition across highland bird communities. However, multivariate analyses could be used instead of diversity indices, since many of these analyses often retain a fairly amount of information on species richness and species abundance across communities (Green 1979, Blondel et al. 1988). Several of these multivariate analyses that are easy to implement, such as cluster analyses or multiple regressions, provide much more information on the study system than any of the diversity indices (Aho and Bush 1993). For example, in the present study Cerro de la Muerte and villa Mills have the most similar pattern of species richness-species abundance, possibly determined by habitat similarity and the short distance that separates both localities. More complex multivariate analyses, such as Canonical Correspondence Analysis, provide very valuable information on environmental variables associated to the presence and abundance of the species in a community (Palmer 1993).
In addition, particular hypotheses associated to changes in species richness across localities, or change in abundance of one, or a group of species can be tested using univariate, bivariate, and/or rarefaction statistical tests (Sanders 1968, Schluter and Ricklefs 1993). For example, the relation between potential habitat available or elevation and species richness across communities could be easily assessed either by using correlation or regression tests, rather than indices of diversity. Rarefaction method, although the species lost their "identity", has proved to be very robust to standardize all samples to a common size (Krebs 1989, Gotelli and Graves 1996, Sackett et al. 2008). Even the simplest method as reporting the number of species per taxonomic category (e.g., family or genus) possibly provides more information than a diversity index value (Sax and Gaines 2003).
In conclusion, many different statistical tests retain more information of patterns of species composition and species abundances across localities than the Shannon-Weiner index, or similar indices. Furthermore, the implementation of different statistical methods will allow testing particular hypotheses on patterns of species richness and species abundance among communities.
Acknowledgments
We thank William G. Eberhard and José
A. Vargas for multiple discussions on the problems inherent to diversity indices, José A. Vargas and Gerardo Avalos for valuable comments on the manuscript and the vicerrectoría de investigación for financial support.
Resumen
Los índices de diversidad y en particular Shannon-Wiener, han sido ampliamente utilizados para el análisis de la diversidad a diferentes escalas geográficas y ecológicas. Estos índices de diversidad poseen serios problemas conceptuales y estadísticos, lo que dificulta o hace imposible la comparación de riqueza o abundancia de especies entre comunidades. Aquí describimos los problemas conceptuales y estadísticos asociados al uso del índice de diversidad Shannon-Wiener, y además presentamos métodos estadísticos alternativos que brindan una mayor información biológica de las especies y permiten una mejor comparación entre comunidades.
Palabras clave: indices de diversidad, riqueza de especies, abundancia de especies, análisis multivariables, avifauna de tierras altas.
Received 31-VIII-2008. Corrected 18-XI-2008. Accepted 16-XII-2008.
References
Adams, J.E. & E.D. McCune. 1979. Application of the generalized jack-knife to Shannons measure of information used as an index of diversity, p. 117-131. In J.F. Grassle, G.P. Patil, W. Smith & C. Taillie (eds.). Ecological diversity in theory and practice. international Cooperative Publishing House, Fairland, Maryland. [ Links ]
Aho, J.M. & A.O. Bush. 1993. Community richness in parasite of some fishes from North America, p. 185-193. In R.E. Ricklefs & D. Schluter (eds.). Species diversity in ecological communities. Historical and geographical perspectives. University of Chicago, Chicago, Illinois, USA. [ Links ]
Andrewartha, H.G. & L.C. Birch. 1954. Selection from the distribution and abundance of animals. University of Chicago, Illinois, USA. [ Links ]
Barrantes, G. in press. The role of historical and local factors in determining species composition of the highland avifauna of Costa Rica and western Panamá. Rev. Biol. Trop. in press. [ Links ]
Barrantes, G. & B.A. Loiselle. 2002. Reproduction, habitat use, and natural history of Phainoptila melanoxantha. Ornitol. Neotrop. 13: 121-136. [ Links ]
Begon, M., C.R. Townsend & J.L. Harper. 2006. Ecology: from individuals to ecosystems. Blackwell, Oxfort, UK. [ Links ]
Bethke, R.W. & T.D. Nudds. 1993. Variation in the diversity of ducks along a gradient of environmental variability. Oecologia 93: 242-250. [ Links ]
Blondel, J., D. Chessel & B. Frochot. 1988. Bird species impoverishment, niche expansion and density inflation in Mediterranean island habitats. Ecology 69: 1899-1917. [ Links ]
Bouton, N., F. Witte, J.J. van Alpen, A. Schenk & O. Seehausen. 1999. Local adaptation in populations of rock-dwelling haplochromines (Pisces: Cichlidae) from southern Lake Victoria. P. Roy. Soc. Lond. B Bio. 266: 355-360. [ Links ]
Cain, M.L., W.D. Bowman & S.D. Hacker. 2008. Ecology. Sinauer, Sunderland, Massachusetts, USA. [ Links ]
Castro-Franco, R., G.G. Vergara García, M.G. Bustos Zagal & W. Mena Arizmendi. 2006. Diversidad y distribución de anfibios del estado de Morelos, México. Acta Zool. Mex. 22: 103-117. [ Links ]
Chao, A. & T.J. Shen. 2003. Nonparametric estimation of Shannons index of diversity when there are unseen species in sample. Environ. Ecol. Stat. 10: 429-443. [ Links ]
Cody, L.M. 1993. Bird diversity components within and between habitats in Australia, p. 147-158. In R.E. Ricklefs & D. Schluter (eds.). Species diversity in ecological communities. Historical and geographical perspectives. University of Chicago, Chicago, Illinois, USA. [ Links ]
Dickman, M. 1968. Some indices of diversity. Ecology 49: 1191-1193. [ Links ]
Forster, R.M., V. Créach, K. Sabbe, W. Vyverman & L.J. Stal. 2006. Biodiversity–ecosystem function relationship in microphytobenthic diatoms of the Westerschelde estuary. Mar. Ecol. Prog. Ser. 311: 191-201. [ Links ]
García Novo, F. 2003. Biodiversidad y conservación de especies. Rev. R. Acad. Cienc. Exact. Fis. Nat. (Esp) 97: 97-120. [ Links ]
Ghent, A.W. 1991. Insights into diversity and niche breadth analyses from exact small-sample tests of the equal abundance hypothesis. Am. Midl. Nat. 126: 213-255. [ Links ]
Grant, P. 1986. Ecology and evolution of Darwins finches. Princeton University, Princeton, New York, USA. [ Links ]
Green, R.H. 1979. Sampling design and statistical methods for environmental biologists. Wiley, New York, USA. [ Links ]
Gotelli, N.J. & G.R. Graves. 1996. Null models in ecology. Smithsonian Institution, Washington D.C., USA. [ Links ]
Haffer, J. 1974. Avian speciation in tropical South America. Nuttall Ornithological Club, Cambridge, Massachusetts, USA. [ Links ]
Hendrickson, J.A., Jr. & P.R. Ehrlich. 1971. An expanded concept of "species diversity". Not. Nat. Acad. Nat. Sci. Philad. 439: 1-6. [ Links ]
Howe, H.F. & L.C. Westley. 1988. Ecological relationships of plants and animals. Oxford University, New York, USA. [ Links ]
Hubbell, S. 2001. The unified neutral theory of biodiversity and biogeography. Princeton University, Princeton. New Jersey, USA. [ Links ]
Humphries, C.J. & L.R. Parenti. 1999. Cladistic Biogeography. Interpreting patterns of plant and animal distributions. Oxford University, New York, USA. [ Links ]
Hurlbert, S.H. 1984. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 54: 187-211. [ Links ]
Hutchinson, G.E. 1959. Homage to Santa Rosalia, or why are there so many kinds of animals? Am. Nat. 93: 145-159. [ Links ]
Krebs, C.J. 1989. Ecological methodology. Harper Collins. New York, New York, USA. [ Links ]
Lloyd, M. & R.J. Ghelardi. 1964. A table for calculating the "equitability" component of species diversity. J. Anim. Ecol. 33: 217-225. [ Links ]
Losos, J. & D.B. Miles. 1994. Adaptation, constraint, and the comparative method: phylogenetic issues and methods, p. 60-98. In P.C. Wainwright & S.M. Reilly (eds.). Ecological morphology. Integrative organismal biology. University of Chicago, Chicago, illinois, USA. [ Links ]
Lourenço, W.R. 2003. Scorpion biogeography: a review, p. 227-240. In J.J. Morrone & J. Llorente Bourquets (eds.). Una perspectiva Latinoamericana de la biogeografía. Las Prensas de Ciencia, Facultad de Ciencias, UNAM, México. [ Links ]
Mac Arthur, R. 1972. Geographical ecology. Patterns in the distribution of species. Princeton University, Princeton, New Jersey, USA. [ Links ]
MacArthur, R.H. & E.O. Wilson. 1967. The theory of island biogeography. Princeton University, Princeton, New Jersey, USA. [ Links ]
Mac Arthur, R. & J.W. Mac Arthur. 1961. On bird species diversity. Ecology 42: 594-598. [ Links ]
Mac Arthur, R.H., J.W. Mac Arthur & J. Preer. 1962. On bird species diversity. II. Prediction of bird census from habitat measurements. Am. Nat. 99: 167-174. [ Links ]
Mac Nally, R., E. Fleishman, L.P. Bulluck & C.J. Betrus. 2004. Comparative influence of spatial scale on beta diversity within regional assemblages of birds and butterflies. J. Biogeogr. 31: 917-929. [ Links ]
Magurran, A.E. 1988. Ecological diversity and its measurement. Princeton University, Princeton, New Jersey, USA. [ Links ]
May, R.M. 1975. Pattern of species abundance and diversity, p. 81-120. In M. Cody & J.M. Diamond (eds.). Ecology and evolution of communities. Harvard University, Cambridge, Massachusetts, USA. [ Links ]
Mayr, E. 1963. Animal species and evolution. Harvard University, Cambridge, Massachusetts, USA. [ Links ]
Moermond, T.C. & J.S. Denslow. 1985. Neotropical avian frugivores: patterns of behavior, morphology, and nutrition, with consequences for fruit selection. Ornithol. Monogr. 36: 865-897. [ Links ]
Monge-Nájera, J. 1994. Ecological biogeography in the Phylum Onychophora. Biogeographica 70: 111-123. [ Links ]
Palmer, M.W. 1993. Putting things in even better order: the advantages of canonical correspondence analysis. Ecology 74: 2215-2230. [ Links ]
Peet, R.K. 1975. Relative diversity indices. Ecology 56: 496-498. [ Links ]
Peterson, G., C.R. Allen & C.S. Holling. 1998. Ecological resilience, biodiversity and scale. Ecosystems 1: 6–18. [ Links ]
Pinkus Rendón, M., G. ibarra-Núñez, v. Parra-Tabla, J.A. García-Ballinas & Y. Hénaut. 2006. Spider diversity in coffee plantations with different management in southeast Mexico. J. Arachnol. 34: 104-112. [ Links ]
Polis, G.A., C.A. Myers & R.D. Holt. 1989. The ecology and evolution of intraguild predation: potential competitors that eat each other. Annu. Rev. Ecol. Syst. 20: 297-330. [ Links ]
Prance, G.T. 1987. Biogeography of neotropical plants, p. 46-65. In T.C. Whitmore & G.T. Prance (eds.). Biogeography and Quaternary history in tropical America. Clarendon, Oxford, England. [ Links ]
Primack, R.B. 1998. Essentials of conservation biology. Sinauer, Sunderland, Massachusetts, USA. [ Links ]
Rawlinson, K.A., J. Davenport & D.K.A Barnes. 2005. Temporal variation in diversity and community structure of a semi-isolated neuston community. P. Roy. Irish. Acad. B. 105B: 107-122. [ Links ]
Ricklefs, R.E. & D. Schluter. 1993. Species diversity in ecological communities. Historical and geographical perspectives. University of Chicago, Chicago, Illinois, USA. [ Links ]
Robertson, D.R. 1996. interspecific competition controls abundance and habitat use of territorial Caribbean damselfishes. Ecology 77: 885-899. [ Links ]
Roig-Juñent, S. & G.E. Flores. 2001. Historia biogeográfica de las áreas áridas de América del Sur austral, p. 257-272. In J.J. Morrone & J. Llorente Bourquets (eds.). Una perspectiva Latinoamericana de la biogeografía. Las Prensas de Ciencia, Facultad de Ciencias, UNAM, México. [ Links ]
Roughgarden, J. & S. Pacala. 1989. Taxon cycle among Anolis lizard populations: review of evidence, p. 403-432. In D. Otte and J.A. Endler (eds.). Speciation and its consequences. Sinauer, Sunderland, Massachusetts, USA. [ Links ]
Roughgarden, J. & J. Diamond. 1986. Overview: the role of species interaction in community ecology, p. 333-343. In J. Diamond & T.J. Case (eds.). Community Ecology. Harper and Row, New York, USA. [ Links ]
Rosenzweig, M.L. 1995. Species diversity in space and time. Cambridge University, Cambridge, New York, USA. [ Links ]
Sackett, T.E., C.M. Buddle & C. Vincent. 2008. Relevance of collected juveniles to the analysis of spider communities. J. Arachnol. 36: 187-190. [ Links ]
Sanders, H.L. 1968. Marine benthic diversity: a comparative study. Am. Nat. 102: 243-282. [ Links ]
Sax, D.F. & S.D. Gaines. 2003. Species diversity: from global decreases to local increases. Trends Ecol. Evol. 18: 561-566. [ Links ]
Schluter, D. 1993. Adaptive radiation in sticklebacks: size, shape, and habitat use efficiency. Ecology 74: 699-709. [ Links ]
Schluter, D. 2000. The ecology of adaptive radiation. Oxford University, New York, USA. [ Links ]
Schoener, T.W. 1974. Resource partitioning in ecological communities. Science 185: 27-39. [ Links ]
Sekercioglu, C.H. 2006. Increasing awareness of avian ecological function. Trends Ecol. Evol. 21: 464–471. [ Links ]
Shapiro, A.M. 1975. The temporal component of butterfly diversity, p. 181-195. In M. Cody & J.M. Diamond (eds.). Ecology and evolution of communities. Harvard University, Cambridge, Massachusetts, USA. [ Links ]
Schluter, D. & R.E. Ricklefs. 1993. Species diversity. An introduction to the problem, p. 1-12. In R.E. Ricklefs & D. Schluter (eds.). Species diversity in ecological communities. Historical and geographical perspectives. University of Chicago, Chicago, illinois, USA. [ Links ]
Simpson, G.G. 1953. The major features of evolution. Columbia University, New York, New York, USA. [ Links ]
Singh, S.P., P. Sah, V. Tyagi & B. Singh Jina. 2005. Species diversity contributes to productivity – Evidence from natural grassland communities of the Himalaya. Curr. Sci. India 89: 548-552. [ Links ]
Slud, P. 1964. The birds of Costa Rica. Distribution and ecology. Bull. Mus. Nat. Hist. 128: 1-430. [ Links ]
Statzner, B. & B. Moss. 2004. Linking ecological function, biodiversity and habitat: a mini-review focusing on older ecological literature. Basic Appl. Ecol. 5: 97-106. [ Links ]
Stiles, F.G. 1985. Seasonal patterns and coevolution in the hummingbird-flower community of a Costa Rican subtropical forest. Ornithol. Monogr. 36: 757-787. [ Links ]
Viloria, A.L. 2003. Historical biogeography and the origins of the satyrine butterflies of the tropical Andes, p. 247-262. In J.J. Morrone & J. Llorente Bourquets (eds.). Una perspectiva Latinoamericana de la biogeografía. Las Prensas de Ciencia, Facultad de Ciencias, UNAM, México. [ Links ]
Washington, H.G. 1984. Diversity, biotic and similarity indices. Water Res. 18: 653-694. [ Links ]
Wiens, J.A. 1989. The ecology of bird communities. Foundations and patterns. Cambridge University, Cambridge, New York, USA. [ Links ]
Whittaker, R.H. 1972. Evolution and measurements of species diversity. Taxon 21: 213-251. [ Links ]
Wolf, L. 1976. Avifauna of the Cerro de la Muerte region Costa Rica. Am. Mus. Novit. No. 2606. [ Links ]
Yue, T.X., S.N. Ma, S.X. Wu & J.Y. Zhan. 2007. Comparative analyses of the scaling diversity index and its applicability. Int. J. Remote Sens. 28: 1611-1623. [ Links ]

