










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.56 n.4 San José Dec. 2008
Estratificación vertical y preferencia de hospedero de las epífitas vasculares de un bosque nublado de Chiapas, México
Nayely Martínez-Meléndez1, Miguel A. Pérez-Farrera1 & Alejandro Flores-Palacios2
1. Herbario Eizi Matuda (HEM), Escuela de Biología, Universidad de Ciencias y Artes de Chiapas, Libramiento Norte-Poniente s/n, Tuxtla Gutiérrez, 29000, Chiapas, México; nayeluci@yahoo.com.mx; perezfarreram@yahoo.com.mx
2.CEAMISH, Universidad Autónoma del Estado de Morelos, Av. Universidad 1001, Col. Chamilpa, 62210, Cuernavaca, Morelos, México; alejandro.florez@uaem.mx
Abstract: Vertical stratification and host preference by vascular epiphytes in a Chiapas, Mexico, cloud forest. The high diversity of vascular epiphytes in neotropical montane forest has been explained as the result of vertical stratification of the forest and specific relationships between epiphytes and their hosts trees at local scales. In a lower montane cloud forest, we studied the vertical stratification and host preferences of vascular epiphytes in a 0.0625 ha plot where 41 trees ≥ of 10 cm DBH were sampled during 12 months in 2001 and 2002. We found 43 epiphyte species growing on 15 tree genera. We tested for vertical strata and host preferences using 19 epiphyte taxa. We found strong evidence that epiphytes divided the canopy, but those epiphyte species dispersed by animals were generalists with respect to hosts and vertical strata. Wind dispersed epiphytes were vertically stratified, with a higher richness in the lowest tree strata. On average the epiphytes preferred 3.5 host species, suggesting low host preference. Two host species, Ardisia and Quercus, were avoided by the majority of the epiphyte species. Our results show that epiphytes divided the canopy horizontally and were stratified vertically, suggesting that host identity could be important determining the abundance of colonizing sites for epiphytes. Rev. Biol. Trop. 56 (4): 2069-2086. Epub 2008 December 12.
Key words: El Triunfo, epiphytes, Chiapas, host tree preference, vertical distribution.
La alta diversidad de epífitas vasculares en bosques neotropicales de montaña ha sido explicada por la existencia de clados que han podido partir los espacios disponibles tanto horizontal como verticalmente (Gentry y Dodson 1987). En una escala local, la partición horizontal de un bosque por las epífitas, se da a través de la diversidad de microambientes y las características estructurales de diferentes hospederos que brindan diferentes oportunidades para el establecimiento, es decir un mosaico complejo de microhábitats o un mosaico físico que provee el medio para su crecimiento. Estos microambientes pueden ser causados por diferencias en la disponibilidad de humedad, de radiación o por la presencia de briofitas (Benzing 1995, Krömer y Gradstein 2003, Krömer et al. 2007a, b). Por ejemplo, en un mismo bosque la comunidad de epí fitas cambia entre las caras de una montaña (Sudgen y Robins 1979) o con la cercanía de ríos (Leimbeck y Balslev 2001). Además en algunos bosques existen hospederos particu lares que hospedan composiciones de epífitas diferentes o sólo a algunas especies, mientras que otros hospederos casi carecen de epífitas (p. ej. Hietz y Hietz-Seifert 1995, Talley et al. 1996, Mehltreter et al. 2005). Aunque se espera que las epífitas puedan establecerse en cualquier hospedero (Callaway et al. 2002), ha habido tres formas de asociación epífita hos pedero que se han documentado. Cuando una epífita está en un sólo hospedero se interpreta que existe especificad de hospedero (ter Steege y Cornelissen 1989), cuando una epífita habita varios hospederos pero está mayoritariamente en uno, se interpreta como preferencia de hospedero (ter Steege y Cornelissen 1989), y cuando una epífita no está en un hospedero se le llama abstinencia de hospedero (host avoidance) y se ha hipotetizado que es el resultado de la expulsión (axeny) de esta epífita por el hospedero (Benzing 1990, 1995). Los tres fenómenos pueden ocurrir en un mismo bosque y le confieren a la comunidad de epífitas oportunidades para aumentar su diversidad, si existen muchas asociaciones específicas y muchos hospederos amigables, o le imponen restricciones si el bosque está compuesto por hospederos hostiles.
La estratificación vertical es el resultado de la variación microclimática desde la parte alta del dosel hasta el interior del bosque y debe ser pronunciada en bosques altos y cerrados, donde el cambio microclimático es más acentuado (Shaw 2004). Esta variación está determinada por la estructura y altura del dosel que regula la intensidad y dirección de luz recibida, la captación de humedad del aire y la temperatura del aire que llega a las plantas; por la disponibilidad de nutrientes, que está asociada a la presencia de materia orgánica muerta o briofitas en los hospederos (Nadkarni 1984, Parker 1995, Walsh 1996, Freiberg 1997, Freiberg y Freiberg 2000, Gradstein et al. 2003, Krömer y Gradstein 2003, Cardelus y Chazdon 2005). En bosques altos esta variación afecta la distribución de la biota del dosel, ya sean plantas no vasculares (Wolf 1995), vasculares (Hietz y Briones 1998, Nieder et al. 1999, Krömer et al. 2007a), o insectos (Nadkarni 1994, Kitching et al. 1997, Stuntz et al. 1999). Aunque se han usado varias aproximaciones para el estudio de la estratificación vertical (e.g. Jarman y Kantvilas, 1995), la zonificación del dosel por Johansson (1974), aún un poco modificada, ha sido la más usada (ter Steege y Cornelissen 1989, Bøgh 1992, Hietz y Hietz-Seifert 1995, Freiberg 1996, 1999, Rudolph et al. 1998, Nieder et al. 1999, Acebey and Krömer 2001, Nkongmeneck et al. 2002, Krömer y Kessler 2006, Krömer et al. 2007a). Johansson (1974) propuso dividir a los árboles en cinco zonas que representan distintos estratos del dosel, en ellas se ha encontrado que la zona I (base del tronco) es la que contiene la mayor parte de las especies exclusivas y que la mayor riqueza de especies se encuentra en las zonas III y Iv que forman la parte interior y central del dosel (Gentry 1982, Nieder et al. 1999). En este trabajo probamos si existe preferencia de hospederos y estratificación vertical en la comunidad de epífitas vasculares del bosque mesófilo de montaña de la Reserva de la Biosfera El Triunfo, Chiapas, México.
Materiales y métodos
Area de estudio: El trabajo fue llevado a cabo en el Cerro del Quetzal (15°4364" N y 92°5563" W, 2 500 msnm) (Fig. 1) que está en el Polígono III de la Reserva de la Biosfera El Triunfo, en la parte central de la Sierra Madre de Chiapas, México. En este cerro la precipitación media anual es de 2 152 mm y la temperatura media anual es de 21.2 ºC (INEGI 1985, García 1987). La vegetación natural es bosque mesófilo de montaña según la clasificación de Rzedowski (1978) que corresponde con un bosque nublado o "lower montane cloud forest" de acuerdo a la clasificación propuesta por Breedlove (1981). El bosque mesófilo de montaña del área de estudio tiene uno o dos estratos de árboles y un sotobosque denso. El dosel puede alcanzar hasta 30 m de altura, es continuo y denso, existen árboles con contrafuertes, perennifolios y con hojas anchas y coriáceas. Esta integrado por los árboles Amphitecna montana L.O. Williams, Liquidambar styraciflua L., Ulmus mexicana Liebm., Oreopanax capitatus (Jacq.) Decne & Planch, Ocotea bernoulliana Mez., Quercus spp., Gentlea tacanensis (Lundell) Lundell, Saurauia oreophila Hemsl., Drimys granadensis L., Symplocos matudae Lundell, Clusia guatemalensis Hemsl., Cavendishia crassifolia (Benth) Heml., Psychotria skutchii Stadl., Podocarpus matudai Lundell, Matudea trinerva Lundell (Pérez Farrera 2004). El epifitismo es característico del bosque mesófilo de montaña, por ejemplo, se han reportado 120 especies para las 450 has del Polígono I de esta reserva (Long y Heath 1991, Martínez-Meléndez 2008), y un total de 492 especies de epífitas vasculares para toda la reserva Para (Martínez-Meléndez et al., en prep.). De estos, las familias con mayor número de especies son Orchidaceae (170 especies), Polypodiaceae (50 especies), Araceae (48 especies) y Bromeliaceae (47 especies).
Toma de datos: El trabajo de campo fue realizado de enero del 2001 a febrero del 2002. En una zona conservada del área, se estableció un cuadrante de 25x25 m (0.0625 ha), en él se identificaron y marcaron todos los árboles con un diámetro a la altura de pecho (DAP) >10 cm. De cada árbol se midió el DAP y fue trepado con técnicas de ascenso de una sola cuerda (Barker 1997, Barker y Sutton 1997). En cada árbol se recolectaron e identificaron las especies de epífitas que crecían en él y se midió la abundancia de estas desde la base del tronco hasta la parte exterior de la copa en las cinco zonas verticales (ZI-v) propuestas por Johansson (1974). Para determinar la abundancia de las epífitas en cada una de estas zonas se contaron el número de plantas no interconectadas (macollos) (Araceae, Araliaceae, Cactaceae, Clusiaceae, Elleanthus, Ericaceae, Liliaceae, Epidendrum), rosetas (Bromeliaceae), hojas (Pteridofitas) o vástagos (Orchidaceae, Piperaceae) que había de cada especie de epífita. Para el conteo se usaron contadores de mano (marca Lee Tools, modelo estándar).
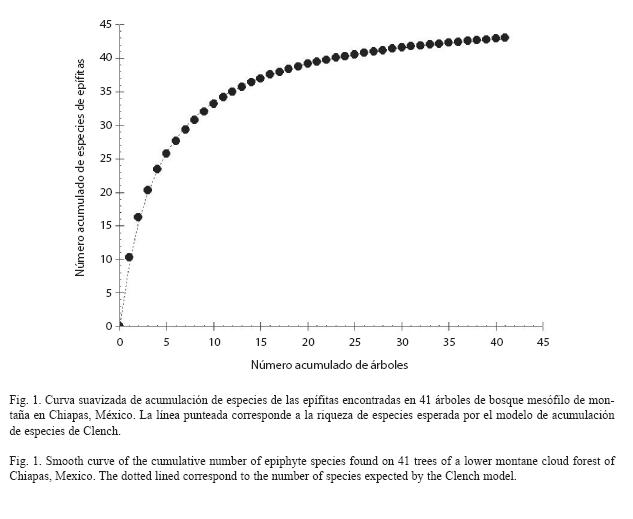
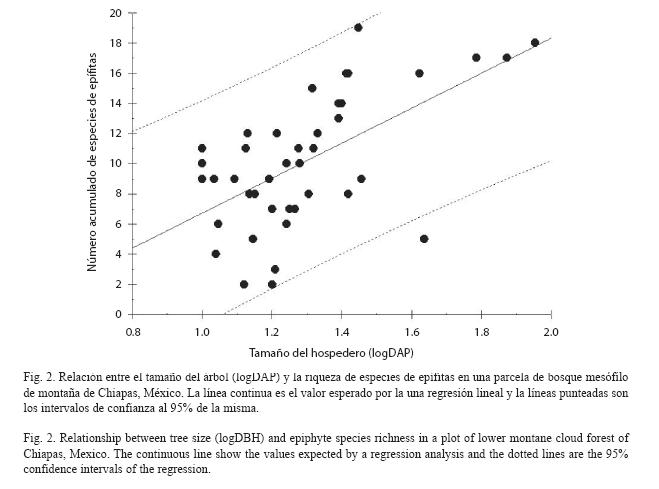
Análisis de datos: Para conocer la eficiencia del muestreo se elaboró una curva suavizada de acumulación de especies con el programa Estimates 8.0 (Coldwell 1997) y se estimó la riqueza de especies de la comunidad utilizando el estimador no paramétrico Chao 1 y el estimador paramétrico Clench que ha sido encontrado útil para predecir la riqueza de especies de epífitas (Flores-Palacios y García-Franco 2001, 2008). Para la elaboración de la curva suavizada se aleatorizó la posición de los árboles iterando en 1 000 ocasiones. Además se analizó si existía una relación entre el tamaño del árbol y la de especies de epífitas como se hace usualmente en trabajos con epífitas (ver Flores-Palacios y García-Franco 2006).
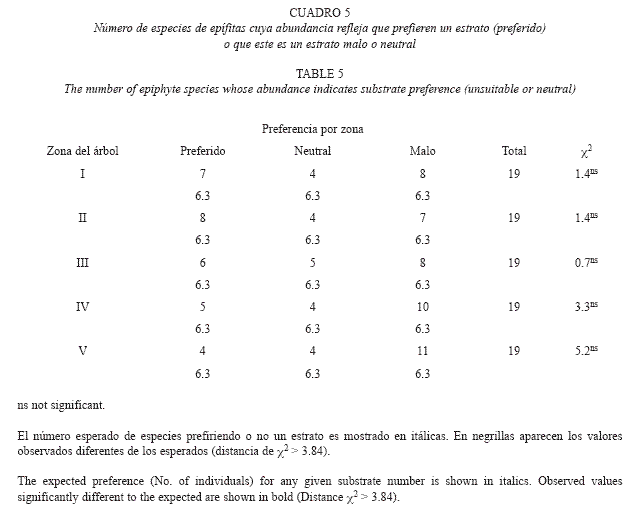
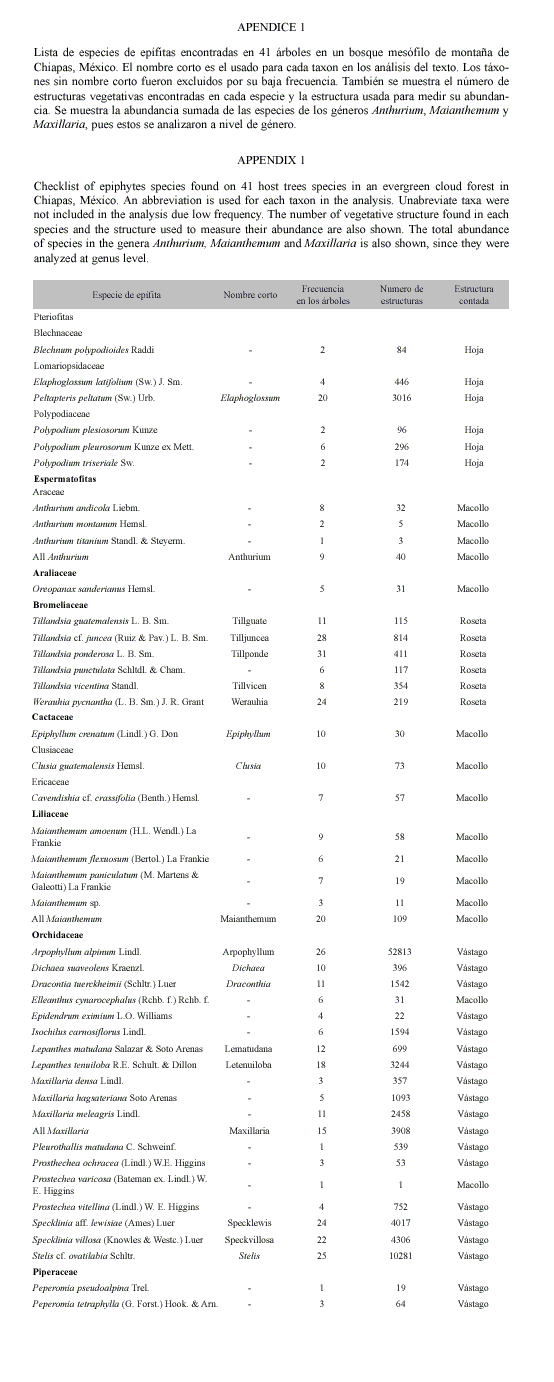
Para determinar la preferencia de hospederos de las especies de epífitas, probamos con una prueba de χ2 si la abundancia observada de 19 táxones de epifitas elegidas en cada especie de hospedero, era similar a la esperada por la abundancia de los hospederos (Zar 1996), como ya ha sido hecho antes (p. ej. Bennett 1987, Mehltreter et al. 2005). Tanto para este análisis como para el de preferencias entre las cinco zonas de Johansson sólo usamos 16 especies de epífitas y tres géneros (Anthurium, Maianthemum y Maxillaria) que fueron encontradas al menos el 20% de los hospederos. Para ponderar la abundancia de las especies de hospederos calculamos el valor de importancia de cada una (Mueller-Dombois y Ellenberg 1974). El valor de importancia fue calculado dividiendo, el número relativo de individuos y el área basal relativa entre dos. El número relativo de individuos y el área basal relativa fueron calculados dividiendo el valor observado de cada especie entre el total de todas las especies. Este valor pondera dos medidas de abundancia de los hospederos, el número de fustes y el volumen de madera que cada especie de árbol ofrece a las epífitas. Aunque encontramos 16 especies de hospederos en nuestra muestra, sólo probamos entre cinco especies y un género (Gentlea, ver resultados), del resto de las especies sólo se encontró un ejemplar y se agruparon en la categoría otras especies. Una vez hecho el análisis de χ2, si las abundancias observadas eran diferentes de las esperadas buscamos que distancias χ2 contribuían a estas diferencia. Consideramos que una especie de epífita tenía preferencia por un hospedero si la abundancia observada era mayor que la esperada y la distancia de χ2 era mayor a 3.84. Consideramos que un hospedero era malo si la abundancia observada era menor que la esperada y la distancia de χ2 era mayor a 3.84. Finalmente concluimos que un hospedero era neutral para una epífita si la abundancia observada era similar a la esperada y la distancia de χ2 era menor o igual a 3.84.
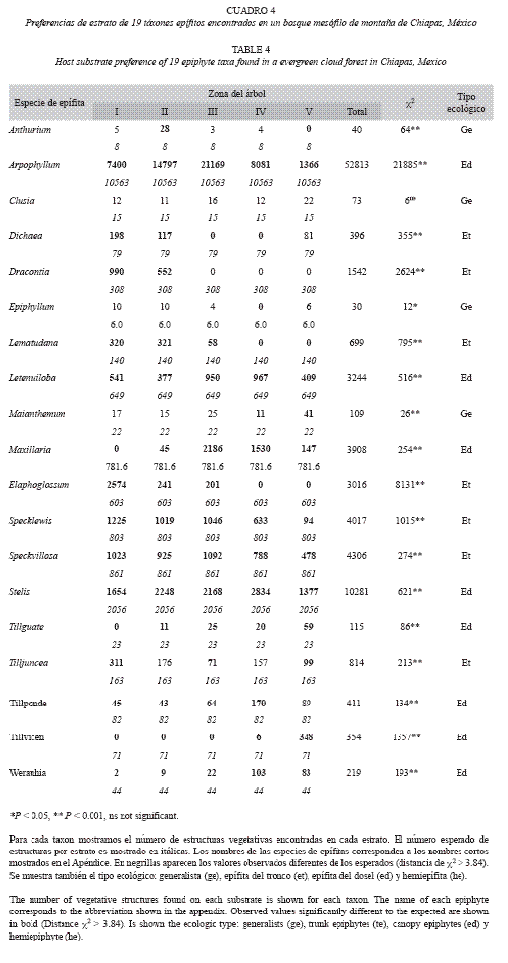
Para determinar si los 19 táxones de epífitas elegidas tenían preferencias entre las cinco zonas de Johansson, con una prueba de χ2 probamos si la abundancia observada de cada especie de epifita en cada zona era similar a la esperada por el azar. Concluimos que una especie de epífita tenía preferencia por un estrato si la abundancia observada era mayor que la esperada y la distancia de χ2 era mayor a 3.84. Consideramos que una epífita evitaba un estrato si la abundancia observada era menor que la esperada y la distancia de χ2 era mayor a 3.84 y que un estrato era neutral para una epífita si la abundancia observada era similar a la esperada y la distancia de χ2 era menor o igual a 3.84. Finalmente en base a este análisis, clasificamos a las epífitas en cuatro tipos ecológicos: de hábito generalistas (que ocurren en tres o más zonas), epífita del dosel superior (que habita en las zonas 3-5), epífita del tronco (que ocurren en las zonas 1-2) tal como se ha hecho por otros autores (Johanson 1974, Acebey et al. 2003, Krömer y Kessler 2006, Krömer 2007a).
Resultados
Preferencias de hospedero: En el cuadrante de 25x25 m encontramos un total de 41 árboles que representan a 16 especies de hospederos de 13 familias (Cuadro 1). Hubo 36 árboles de 10-30 cm, 2 de 31-50 cm, 1 de 51-70 cm y 2 de 71-90 cm, clasificados de acuerdo a su clase diamétrica. De ellos, Gentlea tacanensis fue la especie con mayor valor de importancia y la que tuvo la mayor abundancia de individuos (22%) y de área basal (53%). En el resto del texto nos referimos a las especies de hospederos sólo por el género y se sumaron los valores de importancia de las dos especies de Gentlea para probar por preferencias de hospedero.
Encontramos 43 especies de epífitas (Apéndice). La familia con mayor número de especies fue Orchidaceae (18 especies), seguida de Bromeliaceae (seis especies). Veintisiete especies fueron poco frecuentes para poder probar por preferencias de hospedero o estrato. Para resolver parcialmente este problema se sumo a las especies de los géneros Anthurium, Maianthemum y Maxillaria (Apéndice) y se probó por género. Con esta solución pudimos pro bar por preferencias de hospedero o de estrato entre 16 especies y tres géneros.
El análisis de la curva suavizada de acumulación de especies mostró que se encontraron la mayor parte de las especies de la localidad donde se hizo el muestreo, pues la curva suavizada fue asintótica (Fig. 1) y el número esperado de especies de epífitas por Chao 1 y Clench fue de 43 y 47 especies respectivamente, sugiriendo que el muestreo fue eficiente, pues se muestreó toda la diversidad de especies (Chao 1) o se muestreó el 91.5% de la riqueza de esta localidad (Clench). En promedio encontramos 10.2 ±4.40 especies de epífitas por árbol (media ±desviación estándar, mínimo=2, máximo=19). La riqueza de especies de epífitas se relacionó débilmente con el tamaño del árbol (r2=0.35, p<0.001, Fig. 2).
Los 19 táxones mostraron preferencia por algún hospedero (excepto Epiphyllum) y fueron menos abundantes en al menos una especie de hospedero (Cuadro 2). En promedio cada taxon prefirió 2.1 ± 0.9 de las siete especies de hospederos (media ±desviación estándar, mínimo=0, máximo=4). Los táxones Anthurium, Clusia, Epiphyllum, y Maianthemum son epifitas generalistas, pues entre cuatro y seis especies de los hospederos les resultaron neutrales y prefirieron, o encontraron como un mal hospedero, a 1–2 especies. El resto de las especies se dividió entre aquellas que mayormente tienen hospederos negativos (cuatro hospederos negativos, Arpophyllum alpinum Lindl., Dracontia tuerckheimii (Schltr.) Luer, Lepanthes tenuiloba R.E. Schult. & Dillon, Peltapteris peltata (Sw.) Urb, Specklinia aff. lewisiae (Ames) Luer, Specklinia villosa (Knowles & Westc.) Luer, Stelis cf. ovatilabia Schltr., Tillandsia guatemalensis L.B. Sm., T. ponderosa L.B. Sm. y T. vicentina Standl.), y aquellas que tienen hospederos preferidos o malos (Dichaea suaveolens Kraenzl., Lepanthes matudana Salazar & Soto Arenas, Maxillaria, Tillandsia cf. Juncea (Ruiz & Pav.) L.B. Sm. y Werauhia pycnantha (L.B. Sm.) J.R.) y de ningún a dos hospederos neutrales.
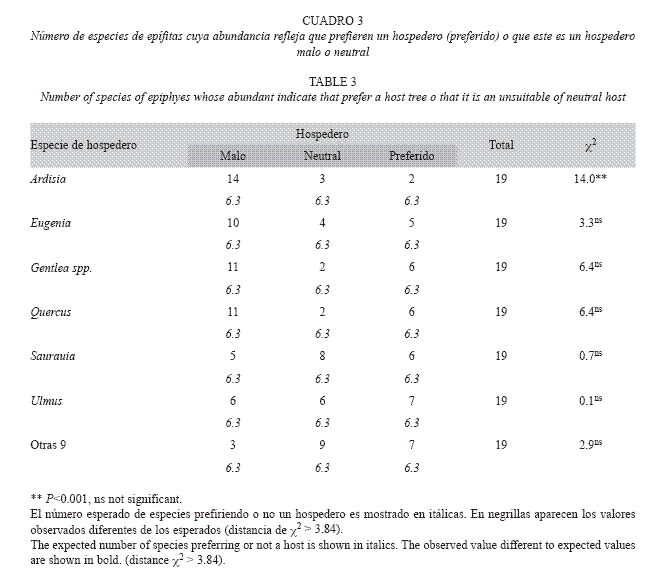
A pesar de que todos los táxones de epífitas mostraron preferencia por algún hospedero, sólo la especie de hospederos Ardisia mostró tener más relaciones negativas con la flora epífita de lo esperado por el azar (Cuadro 3). El resto de los hospederos resultaron ser tan buenos como malos para las epífitas.
Preferencias de estrato vertical: Con la excepción de Clusia guatemalensis, los 19 táxones epífitas mostraron algún tipo de preferencia por algún estrato o alguna zona de Johansson (Cuadro 4). En promedio los táxones prefirieron 1.6±0.9 estratos (mínimo=0, máximo=3), evitaron 2.3±1.1 (mínimo=0, máximo=4) y les resulto neutral 1.1±1.5 (mínimo=0, maximum=5) estratos.
Las epífitas Anthurium, Clusia, Epiphyllum y Maianthemum son generalistas, pues su abundancia mostró que no prefieren ningún estrato o les son neutrales tres o cuatro estratos mientras que uno les es desfavorable o es preferido (Cuadro 4). Dichaea suaveolens, Dracontia tuerckheimii, Lepanthes matuda na, Elaphoglossum peltatum y Tillandsia. cf. juncea prefirieron los estratos inferiores I y II (Cuadro 4). Las especies Specklinia aff. lewisiae y Specklinia villosa, también prefirieron los estratos bajos del dosel pero ampliaron sus preferencias hasta el estrato III (Cuadro 4). Estas siete especies pueden ser conside radas como "especialistas del tronco" ya que ocurren principalmente entre la zona I y II (sensu Krömer et al. 2007a). Cuatro especies prefirieron los estratos intermedios y evitaron los estratos bajos o altos, Arpophyllum alpinum, Lepanthes tenuiloba, Maxillaria y Stelis cf. ovatilabia que fueron más abundantes en los estratos II y III o III y Iv (Cuadro 4). Finalmente, las especies Tillandsia guatemalensis, T. ponderosa, T. vicentina Standl. y W. pycnantha prefirieron el estratos exterior (V) o se distribuyeron del interior del dosel al exterior (III a v) y evitaron los estratos interiores. Estas ocho especies podrían ser consideradas como "especialistas del dosel" ya que pueden preferir hasta tres zonas entre la III y la v (sensu Krömer et al. 2007a)
A pesar de la estratificación vertical de los táxones de epífitas en ningún estrato se concentraron las preferencias (Cuadro 5). Sin embargo, la riqueza acumulada de epífitas os especies, en el III 32 especies, en el IV 21 especies y en el estrato v 24 especies.
Del resultado de esta estratificación surgen dos patrones. El primero es que las epífitas dispersadas por animales tendieron a ser generalistas y las epífitas dispersadas por el viento fueron las que prefirieron hospederos y estratos.
Discusión
El bosque del cuadrante estudiado contiene una riqueza de especies de epífitas similar a la encontrada en bosques neotropicales de la misma altitud (Wolf y Flamenco 2003). Por ejemplo en este estudio se encontraron 43 táxones en 625 m2, similar a lo encontrado en veracruz, México, a una altitud de 1 439 m registraron 53 especies en 625 m2 (Hietz y Hietz-Seifert 1995) y a una altitud de 1 500-1 600 m se ha encontrado entre 37 a 88 especies de epífitas en parcelas de 0.1 ha de bosque mesófilo de montaña (Flores-Palacios y García-Franco 2008). Otros resultados sugieren que los bosque montanos de Bolivia son más ricos y en ellos se han encontrado hasta 175 especies en 0.32 ha (1 600 m de altitud) (Krömer et al. 2007a). Como era esperado, encontramos evidencia de que las epífitas del bosque mesófilo de montaña del Triunfo, se dividen el dosel, es decir, pueden preferir más de un estrato.
Las epífitas de los géneros Anthurium, Clusia, Epiphyllum y Maianthemum tienen frutos carnosos en forma de baya dispersados por animales, principalmente aves y murciélagos (endozoocoria). Este síndrome de dispersión les permite a las semillas germinar envueltas en materia orgánica y hace menos importante la interacción entre la epífita y el árbol. Es posible que esto explique la distribución generalista de estos géneros en el área estudiada. A diferencia de estos géneros, las Pteridofitas, Bromeliaceae y Orchidaceae, epífitas tienen esporas o semillas diminutas, que para establecerse interactúa directamente con la corteza de los árboles, los cojines de criptógamos o el substrato orgánico sobre las ramas horizontales junto con otras epífitas vasculares (Benzing 1990, Krömer y Gradstein 2003). Así que para estas últimas epífitas la interacción con la corteza de los hospederos y otras epífitas podría ser más importante en el establecimiento.
Se espera que las epífitas vasculares tengan patrones específicos de estratificación vertical en un forofito y estos patrones deberían estar relacionados con las diferencias en la tolerancia a las condiciones de luz y humedad o sus adaptaciones ecofisiológicas (Johansson 1974, ter Steege y Cornelissen 1989, Jácome et al. 2004, Krömer et al. 2007a). Por ejemplo, la luz disminuye y la humedad aumenta desde el dosel hasta el piso boscoso (Parker 1995). En el bosque estudiado, la estratificación de las epífitas fue alta, ya que la mayoría de los táxones mostraron algún tipo de preferencia por algunas zonas, y asimismo podrían ser clasificadas en epífitas del tronco o dosel. Sin embargo, sólo Elaphglossum peltatum y Tillandsia vicentina mostraron preferencia por solo una zona mientras se abstenían del resto. El resto de los táxones preferían más de un estrato o preferían uno y tenían estratos neutrales, lo que sugiere traslape de nichos (Nieder et al. 1999). Aún con este traslape, para el bosque estudiado es posible identificar especies de epífitas que corresponden con cada zona del dosel.
En la mayoría de los trabajos se ha reportado que las zonas del dosel III y Iv de Johansson (1974) son los que concentran la mayor riqueza de especies. (e.g. ter Steege y Cornelissen 1989, Nieder et al. 1999, Krömer et al. 2007a). Nosotros no encontramos evidencia que favorezca este patrón, pues en ningún estrato se concentraron las preferencias y fue la zona I la más rica en especies. Este patrón se ocasionó por la abundancia de especies que prefieren el estrato I y II (siete especies), helechos y hemiepífitas y tal vez es una propiedad de este bosque, pero es necesario un estudio más extenso para saber si este es el patrón en este bosque montañoso.
Se ha hipotetizado que las epífitas deberían tener estrategias de establecimiento gene- ralistas, que les permitan colonizar cualquier hospedero, haciendo irrelevante la identidad del mismo (Callaway et al. 2002). Nuestros datos muestran que las epífitas logran esta blecimientos generalistas, pero que hay una tendencia débil a preferir a algunos hospederos. Menos de la mitad de las especies de epífitas (cinco) mostraron mayormente preferencia o neutralidad por al menos cuatro hospederos, lo que sugiere baja preferencia. La abundancia de estas especies debe estar determinada por la abundancia de hospederos preferidos. A pesar de estos patrones ninguna de las especies de epífitas fue específica a un hospedero. Lo cual reitera la rareza de la especificad de hospedero (Benzing 1990) y la tendencia entre las epífitas a habitar en más de una especie de árbol. Es más destacable que para cinco especies de epífitas el bosque es un escenario de buenos o malos hospederos, y que la mayor parte (11 especies) de las especies de epífitas tuvieron mayormente hospederos malos. Esto sugiere que los hospederos del bosque estudiado podrían limitar a la mayor parte de las especies de epífitas, ya sea por que los árboles se saturan con otras especies de epífitas (Flores-Palacios y García-Franco 2006) o por que tienen condiciones en su corteza que limitan el establecimiento de algunas especies, cualquiera de estas hipótesis deben ser probadas con nuevos datos.
El papel principal de los hospederos parece ser simplemente proveer de substrato a las epífitas, sin embargo las epífitas difieren en abundancia de acuerdo a la especie hospedera (Went 1940, Johansson 1974, Benzing 1981, Bennett 1986, ter Steege y Cornelisen 1989, Migenis y Ackerman 1993, Dejean et al. 1995, Kernan y Fowler 1995) es decir, existen varias características de las especies de forofitos que afectan la presencia y abundancia de las epífitas (Frei y Dodson 1972, Schlesinger y Marks 1977, Benzing 1995, Krömer y Gradstein 2003). Ejemplo de estas características son el tamaño del árbol, lo cual puede regular la intensidad de luz captada por las epífitas del dosel o del interior; la estructura del árbol, la presencia del número de ramas gruesas será proporcional a la cantidad de materia orgánica en ellas; la textura de la corteza y características químicas, una corteza rugosa tiene un efecto positivo sobre la colonización de epífitas, aunque algunas sustancias emitidas pueden ocasionar lo contrario; la disponibilidad de agua y nutrientes también son necesarios para el establecimiento de estas (ter Steege y Cornelissen 1989, Hietz y Hietz-Seifert 1995, Callaway et al. 2002, Krömer et al. 2007a, b). Nuestros datos muestran que no hubo un hospedero que concentrara las preferencias de las epífitas, pero sí hubieron dos especies de hospederos que resultaron ser malos hospederos para la mayor parte de las epífitas, Ardisia y Quercus. Es posible que Ardisia resultara un mal hospedero por que posee corteza exterior lisa y suele tener fustes rectos, sin muchas ramificaciones, por lo que podría no ser capaz de proteger a las semillas del escurrimiento de agua. Y Quercus, probablemente por su escasa representación en el cuadrante de estudio, lo que sugiere poca superficie para el establecimiento de las comunidades epífitas.
Nuestros resultados muestran que en un bosque diverso en epífitas vasculares, éstas parten el dosel horizontalmente y se estratifican verticalmente. Además sugieren que la identidad de los hospederos es importante porque puede determinar los sitios colonizables para muchas epífitas, ya sea limitándolas o favoreciéndolas. Nuestros datos sugieren que en este bosque, sólo las epifitas dispersadas por endozoocoria escapan a la estratificación vertical y a las preferencias de hospedero.
Agradecimientos
Agradecemos a los guardaparques Marco Tulio, Luís, Pedro, Ismael. A Rubén Martínez-Camilo y J. Fernando Rodríguez-García su ayuda en el trabajo de campo. Miguel Angel Soto Arenas y Robert L. Dressler ayudaron con la identificación de las especies. A Thorsten Krömer por la revisión de este manuscrito en español. A Andrew vovides por la revisión en inglés. Este proyecto fue financiado por SIBEJ-CONACYT (proyecto 20000506014) y Christopher Davidson y Sharon Christoph. Una versión previa de este trabajo fue presentada por N. Martínez-Meléndez como su tesis de licenciatura en Escuela de Biología de la Universidad de Ciencias y Artes de Chiapas.
Resumen
En escala local, la alta diversidad de epífitas vasculares en bosques montanos neotropicales ha sido explicada por que logran estratificar verticalmente al dosel y tienen relaciones específicas con sus hospederos. En un bosque de montaña estudiamos la estratificación vertical y las preferencias de hospedero de las epífitas vasculares en un cuadrante de 0.0625 ha en donde muestreamos 41 árboles ≥10 DAP, durante 12 meses en los años 2001 y 2002.
Encontramos 43 especies de epífitas en 15 géneros de árboles. Probamos preferencia de estratos verticales o de hospedero con 19 táxones epífitos. Encontramos evidencia de que las epífitas se dividen el dosel; aunque, las epífitas dispersadas por animales tendieron a ser generalistas. Entre las epífitas dispersadas por el viento reconocimos una clara estratificación vertical, concentrando especies en la parte baja de los árboles. En promedio las especies de epífitas prefirieron 3.5 especies de hospederos, sugiriendo una baja preferencia. Dos especies de árboles resultaron ser malos hospederos, Ardisia y Quercus. Nuestros resultados muestran que las epífitas parten el dosel horizontalmente, se estratifican verticalmente y sugieren que la identidad de los hospederos es importante por que puede determinar la abundancia de sitios colonizables para muchas epífitas.
Palabras Clave: El Triunfo, epífitas, Chiapas, forofitos, distribución vertical.
Recibido 17-VIII-2007. Corregido 30-VI-2008. Aceptado 31-VII-2008.
Referencias
Acebey, A. & T. Krömer. 2001. Diversidad y distribución vertical de epífitas en los alrededores del campamento río Eslabón y de la Laguna Chalalán, Parque nacional Madidi, Dpto. La Paz, Bolivia. Rev. Soc. Boliviana Bot 3: 104-123 [ Links ]
Barker, M. 1997. An update on low-tech methods for forest canopy access and on sampling a forest canopy. Selbyana 18: 61–71. [ Links ]
Barker, M.G. & S.L. Sutton. 1997. Low-tech methods for forest canopy access. Biotrópica 29: 243-247 [ Links ]
Beckner, J. 1979. Host trees for cultivated orchids. Amer. Orchid Soc. Bull. 48: 792-795. [ Links ]
Bennett, B.C. 1986. Patchiness, diversity, and abundance relationships of vascular epiphytes. Selbyana 9: 70-75. [ Links ]
Bennett, B.C. 1987. Spatial distribution of Catopsis and Guzmania (Bromeliaceae) in southern Florida. Bull. Torrey Bot. Club. 114: 265–271. [ Links ]
Benzing, D.H. 1981. The population dynamics of Tillandsia circinata (Bromeliaceae): cypress crown colonies in southern Florida. Selbyana 5:256-263. [ Links ]
Benzing, D.H. 1990. Vascular Epiphytes. Cambridge University, Massachusetts, EEUU. [ Links ]
Benzing, D.H. 1995. Vascular Epiphytes, p. 225-254. In M.D. Lowman & n.M. nadkarni (eds.). Forest canopies. Academic, San Diego, California, EEUU. [ Links ]
Breedlove, D. E. 1981. Flora of Chiapas Part 1. Introduction to the Flora of Chiapas. The California Academy of Science, San Francisco, California, USA. [ Links ]
Bogh, A. 1992. Composition and distribution of the vascular epiphyte flora o fan Ecuadorian montane rain forest. Selbyana 13: 25-34. [ Links ]
Callaway, R.M., K.O. Reinhart, G.W. Moore, D.J. Moore & S.C. Pennings. 2002. Epiphyte host preferences and host traits: mechanisms for species specific interactions. Oecología 132: 221-230. [ Links ]
Cardelús, C.L. & R.L. Chazdon (2005). Inner-crown microenviroments of two emergent tree species in a lowland wet forest. Biotropica 37: 238-244. [ Links ]
Castro-Hernández, J.C., J.H.D. Wolf, J.G. García-Franco & M. González-Espinosa. 1997. The influence of humidity, nutrients and light on the establishment of the epiphytic bromeliad Tillandsia guatemalensis in the highlands of Chiapas, Mexico. Rev. Biol. Trop. 47: 763–773. [ Links ]
Dejean, A., I. Olmstead & R.R. Snelling. 1995. Treeepiphyte-ant relationships in the low inundate forest of the Sian Kaan Biosphere Reserve, Quintana Roo, Mexico. Biotropica 27: 57-70 In R. [ Links ]M. Callaway, K.O. Reinhart, G.W. Moore, D.J. Moore & S.C. Pennings. 2002. Epiphyte host preferences and host traits: mechanisms for species specific interactions. Oecología 132: 221-230.
Flores-Palacios, A. & J.G. García-Franco. 2001. Sampling methods for vascular epiphytes: their effectiveness in recording species richness and frequency. Selbyana 22: 181–191. [ Links ]
Flores-Palacios, A. & J.G. García-Franco. 2006. The relationship between tree size and epiphyte richness: testing four different hypotheses. J. Biogeog. 33: 323–330. [ Links ]
Flores-Palacios, A. & J.G. García-Franco. 2008. Habitat isolation changes the beta diversity of the vascular epiphyte community in lower montane forest, veracruz, Mexico. Biodivers. Conserv. 17: 191–207. [ Links ]
Frei, J.K. & C.H. Dodson. 1972. The chemical effect of certain bark substrates on the germination of early growth epiphytic orchids. Bull. Torrey Bot. Club 99: 301-307. [ Links ] In R.M. Callaway, K.O. Reinhart, G.W. Moore, D.J. Moore & S.C. Pennings. 2002. Epiphyte host preferences and host traits: mechanisms for species specific interactions. Oecologia 132: 221-230.
Freiberg, M. 1997. The influence of epiphyte cover on branch temperature in a tropical tree. Plant. Ecol. 153: 241-250. [ Links ]
Freiberg, M. 1999. The vascular epiphytes on a virola michelii tree (Myristicaceae) in French Guiana. Ecotropica 5: 78-81. [ Links ]
Freiberg, M. & E. Freiberg. 2000. Epiphyte diversity and biomass in the canopy of lowland and montane forest in Ecuador. J. Trop. Ecol. 16: 673-688. [ Links ]
García, E. 1987. Modificaciones al sistema de clasificación climática de Köppen (para adaptarlo a las condiciones de la República Mexicana). Univ. Nac. Aut. de Méx. México, México DF, México. [ Links ]
Gentry, A.H. 1982. Patterns of neotropical plant species diversity, p. 1-84. In M.K. Hecht, B. Wallace & G.T. Prance (eds.). Evolutionary biology, Plenum, New York, EEUU. [ Links ]
Gentry, A.H. & C. Dodson. 1987. Diversity and biogeography of neotropical vascular epiphytes. Ann. Missouri Bot. Gard. 74: 205-233. [ Links ]
Gradstein, S.R., N.M. Nadkarni, T. Krömer, I. Holz & N. Nöske. 2003. A protocol for rapid and representative sampling of vascular and non-vascular epiphyte diversity of tropical rain forest. Selbyana 24: 105-111. [ Links ]
Hietz, P. & O. Briones. 1998. Correlation between water relations and within-canopy distribution of epiphytic ferns in a Mexican cloud forest. Oecologia 114: 305-316. [ Links ]
Hietz, P. & U. Hietz-Seifert. 1995. Composition and ecology of vascular epiphyte communities along an altitudinal gradient in central veracruz, Mexico. J. veg. Sci. 6: 487–498. [ Links ]
Jácome, J., G. Galeano, M. Amaya & M. Mora. 2004. vertical distribution of epiphyte and hemiepihpytic Araceae in a tropical rain forest in Chocó, Colombia. Selbyana 23: 118-123. [ Links ]
Jarman, S. & G. Kantillas. 1995. Epiphytes on an old Huon Pine tree Legarastrobos in Tasmanian rainforest. New Zeal. J. Bot. 33:65-78. [ Links ]
Johansson, D. 1974. Ecology of vascular epiphytes in west African rain forest. Acta Phytogeograpica Suecica. 59: 1–136. [ Links ]
INEGI, 1985. Carta de Climas. villahermosa escala 1: 1000000. México. [ Links ]
Kernan, C. & n. Fowler. 1995. Diferencial substrate use by epiphytes in Corcovado Nacional Park, Costa Rica: a source of guiad structure. J. Ecol. 83:65-73. [ Links ]
Kitching, R.L., H. Mitchell, G. Morse & C. Thebaud. 1997. Determinants of species richness in assemblages of canopy leaf litter ants (Hymenoptera: Formicidae) in a neotropical montane forest. Psyche 97: 81-93. [ Links ]
Krömer, T. & S.R. Gradstein. 2003. Species richness of vascular epiphytes in two primary forest and fallows in the Bolivian Andes. Selbyana 24: 190-195. [ Links ]
Krömer, T. & M. Kessler. 2006. Filmy ferns (Hymenophyllaceae) as high-canopy epiphytes. Ecotropica 12: 57-63. [ Links ]
Krömer, T., M. Kessler & S.R. Gradstein. 2007a. vertical stratification of vascular epiphytes in submontane and montane forest of the Bolivian Andes: the importance of the understory. Plant Ecol. 189: 261-278. [ Links ]
Krömer, T., S.R. Gradstein & A. Acebey. 2007b. Diversidad y ecología de epífitas vasculares en bosques montanos primaries y secundarios de Bolivia. Ecol. Bolivia 42: 23-33. [ Links ]
Leimbeck, R.M. & H. Balslev. 2001. Species richness and abundance of epiphyte Araceae on adjacent floodplain and upland forest in Amazonian Ecuador. Biodivers. Conserv. 10: 1579–1593. [ Links ]
Long, A. & M. Heath. 1991. Flora of the El Triunfo Biosphere Reserve, Chiapas, México: a preliminary floristic inventory and the plant communities of polygon I. Anales Inst. Biol. Univ. Autón. Méx. Ser. Bot. 62: 133-172. [ Links ]
Martínez-Meléndez, J., M.A. Pérez-Farrera & O.Farrera-Sarmiento. 2008. Inventario florístico del Cerro El Cebú y zonas adyacentes en la Reserva de la Biosfera El Triunfo (Polígono v), Chiapas, México. Bol. Soc. Bot. Méx. 82: 21-40. [ Links ]
Martínez-Meléndez N., M.A. Pérez-Farrera y R. Martínez-Camilo (en prep.). Las epífitas de la reserva de la biosfera El Triunfo, Chiapas, México. [ Links ]
Mehltreter, K., A. Flores-Palacios & J.G. García-Franco. 2005. Host preferences of low-trunk vascular epiphytes in a cloud forest of veracruz, Mexico. J. Trop. Ecol. 21: 651–660. [ Links ]
Migenis, L.E. & J.D. Ackerman. 1993. Orchid-sporophyte relationships in a forest watershed in Puerto Rico. J. Trop. Ecol. 9: 231-240. [ Links ] In R.M. Callaway, K.O.
Reinhart, G.W. Moore, D.J. Moore & S.C. Pennings. 2002. Epiphyte host preferences and host traits: mechanisms for species specific interactions. Oecología 132: 221-230.
Mueller-Dombois, D. & H. Ellenberg. 1974. Aims and methods of vegetation ecology. John Wiley and Sons, Nueva York, Nueva York, EEUU. [ Links ]
Nadkarni, n.M. 1994. Diversity of species and interactions in the upper tree canopy of forest ecosystems. Am. Zool. 34: 70-78. [ Links ]
Nieder, J., S. Engwald & W. Barthlott. 1999. Patterns of neotropical epiphyte diversity. Selbyana 20: 66-75. [ Links ]
Nkongmeneck Bernard-Aloys, M.D. Lowman & J.T. Atwood. 2002. Epiphyte diversity in primary and fragmented forests of cameroon. Selbyana 23: 121-130. [ Links ]
Parker, G.G. 1995. Structure and microclimate of forest canopies, p. 73-106. In M.D. Lowman & n.M. Nadkarni (eds.). Forest canopies. Academic, San Diego, California, EUUU. [ Links ]
Pérez-Farrera, M.A. 2004. Flora y vegetación de la reserva de la biosfera El Triunfo: diversidad, riqueza y endemismo, p. 77-100. In M.A. Pérez-Farrera, N. Martínez-Meléndez, A. Hernández-Yáñez & v. Arreola-Muñoz (eds.). La Reserva de la Biosfera El Triunfo, Tras una Década de Conservación, Universidad de Ciencias y Artes de Chiapas, México DF, México. [ Links ]
Rudolph, D., G. Rauer, J. nieder & W. Barthlott. 1998. Distributional patterns of epiphytes in the canopy and phorophyte characteristics in a western Andean rain forest in Ecuador. Selbyana 19: 27-33. [ Links ]
Rzedowski, J. 1978. vegetación de México. Limusa, México DF, México. [ Links ]
Schlesinger, W. & P. Marks. 1977. Mineral cycling and the niche of Spanish moss, Tillandsia usneoides L. Amer. J. Bot. 64: 1254-1262. [ Links ]
Shaw, D.C. 2004. Vertical organization of canopy biota, p. 73-101. In M.D. Lowman & H.B. Rinker (eds.). Forest canopies (second edition). Elselvier Academic, San Diego, California, EEUU. [ Links ]
Sugden, A.M. & R.J. Robins. 1979. Aspects of the ecology of vascular epiphytes in Colombian cloud forest, I. The distribution of the epiphytic flora. Biotropica 11: 173-188. [ Links ]
Stuntz, S., U. Simon & G. Zotz. 1999. Assessing the potential influence of vascular epiphytes on arthropod diversity in tropical tree crowns: hypotheses, approaches and preliminary data. Selbyana 20: 276-283. [ Links ]
Talley, S.M., R.O. Lawton & W.N. Setzer. 1996. Host preference of Rhus radicans (Anacardiaceae) in a southern deciduous hardwood forest. Ecology 77: 1271-1276 In R. [ Links ]M. Callaway, K.O. Reinhart, G.W. Moore, D.J. Moore & S.C. Pennings. 2002. Epiphyte host preferences and host traits: mechanisms for species specific interactions. Oecología 132: 221-230.
ter Steege, H. & J.H.C. Cornelissen. 1989. Distribution and ecology of vascular epiphytes in lowland rain forest of Guyana. Biotropica 21: 331–339. [ Links ]
Walsh, R.P.D. 1996. Microclimate and hydrology, p. 206-236. In P.W. Richards (ed.). The tropical rainforest. Cambridge, Cambridge, UK. [ Links ]
Went, F.W. 1940. Soziologie der epiphyten eines tropischen Urwaldes. Ann. Jard. Bot. Buitenz. 50: 1-98. [ Links ]
Wolf, J.H. 1995. Non vascular epiphyte diversity patterns in the canopy of an upper montane rain forest (2 550-3 670 m), Central Cordillera, Colombia. Selbyana 16: 185-195. [ Links ]
Wolf, J.H.D. & A. Flamenco. 2003. Patterns in species richness and distribution of vascular epiphytes in Chiapas, Mexico. J. Biogeog. 30: 1689–1707. [ Links ]
Zar, J.H. 1996. Biostatistical Analysis. Prentice-Hall Inc., Englewood Cliffs, Nueva Jersey, EEUU. [ Links ]
Referencias de internet
Colwell, R.K. 1997. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. version 5 Users guide and Application, University of Connecticut, Storrs. (Consultado: 28 julio 2008; http: viceroy.eeb.uconn.edu/estimates). [ Links ]


{kind=link}
{kind=link}