










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.56 n.4 San José Dec. 2008
Indicadores morfológicos y reproductivos del pez Gambusia puncticulata (Poeciliidae) en sitios muy contaminados del río Almendares, Cuba
Yureidy Cabrera Páez, Consuelo Aguilar Betancourt & Gaspar González-Sansón
Centro de Investigaciones Marinas, Universidad de La Habana, Cuba Calle 16e/1ra y 3ra, No.114, CP: 11300, Miramar, Ciudad de La Habana, Cuba; ycabrera@cim.uh.cu
Abstract: Reproductive and morphological indicators of the fish Gambusia puncticulata (Poeciliidae) in very polluted sections of Almendares River, Cuba. The Almendares River watershed covers a large portion of Havana city. Human activities have negative impacts over it. We studied the alteration of biological processes in fish from polluted sites. Stressor-based cumulative effects assessment was applied at individual level in Gambusia puncticulata (Poey, 1854). Three sites of Almendares River were sampled monthly from July 2006 to June 2007. Length and weight of females, and condition factor for both sexes, were higher in the impacted sites. An increase of food supply due to eutrophication can explain why fishes are heavier and longer at polluted sites. Gonadosomatic, specific fecundity index and embryo numbers were significantly higher on the reference site. The Hepatosomatic index was higher in females from the most contaminated sites. Differences in δ13C and δ15N among contaminated and reference sites suggest that the food source is not the same. Rev. Biol. Trop. 56 (4): 1991-2004. Epub 2008 December 12.
Key words: Almendares River, contamination, Gambusia puncticulata, indicators, Cuba.
Los peces de agua dulce se encuentran hoy amenazados por la continua agresión del medio. Esto se debe a los vertimientos de residuales, tanto domésticos como industriales, que contaminan las aguas, provocando la muerte de las poblaciones existentes. Existen varias razones para considerar a los peces como organismos útiles para medir la degradación ambiental, ya que la ictiofauna actúa como integradora de los impactos directos e indirectos sobre el ecosistema acuático (Chapman 2002).
Evaluar y estimar los efectos de la actividad humana sobre el ecosistema acuático es una tarea compleja. Debido a esto, han sido propuestos un gran número de bioindicadores y pruebas a organismos para la evaluación de ecotoxicidad de compuestos antropogénicos en ambientes acuáticos (Nunes et al. 2005). En esta investigación se realiza un análisis de la variación de indicadores biológicos seleccionados, en relación con el impacto humano. Este estudio es uno de los primeros en realizarse en un ambien- te dulceacuícola cubano, utilizando como base una metodología para la detección del impacto ambiental, basada en los efectos observados en organismos indicadores.
El objetivo principal fue evaluar las alteraciones de los procesos biológicos en los peces que viven en sitios muy contaminados, aplican do una metodología basada en la detección de los efectos acumulativos de agentes estresantes múltiples a nivel de individuo.
Materiales y métodos
Zona de estudio: La investigación se realizó sobre poblaciones naturales de peces en tres sitios diferentes a lo largo del río Almendares. Los sitios de muestreo fueron: El Rodeo, considerado como sitio de referencia, y dos sitios altamente contaminados, el Bosque de La Habana y el Puente 23. La zona más alta del río (Rodeo) presenta una contaminación dada fundamentalmente por materia orgánica. La contaminación presente en el Bosque de La Habana es de naturaleza orgánica e industrial. Sus aguas se caracterizan por presentar altos valores de amonio, nitrito, nitrato y coliformes fecales proporcionándole al área una ligera eutrofización. Estudios realizados en los sedimentos mostraron la presencia de metales pesados como el cobre, plomo y zinc, clásicos indicadores de contaminación antropogénica (Olivares-Rieumont et al. 2005). Esto mismo fue encontrado para el Puente 23 (zona más baja del río), pero con niveles mucho más altos que exceden los reportes internacionales para sedimentos y calidad de las agua en ríos no contaminados. Se reportó, además, la presencia de hidrocarburos, grasas y aceites, lo que demuestra el deterioro considerable de la calidad de las agua en esta zona. Por lo que a lo largo del río Almendares se observa claramente un gradiente de contaminación, que va aumentando hacia la parte más baja del río. Esto está en correspondencia con los valores de oxígeno disuelto, los cuales tienden a disminuir hacia la desembocadura (4.6mg/L en el Rodeo, 0.4mg/L en el Bosque de La Habana y 0.4mg/L en el Puente 23), por el contrario, los valores de la demanda bioquímica de oxígeno (DBO5) aumentan (6.3mg/L en el Rodeo, 30.2mg/L en el Bosque de La Habana y 29.6mg/L en el Puente 23). Los tres sitios, además, están afectados por las mismas condiciones climáticas.
Análisis biológico: La investigación se realizó sobre poblaciones naturales de peces en tres sitios diferentes a lo largo del río Almendares. Se capturaron peces mensualmente en el período comprendido entre julio del 2006 y junio del 2007. Los peces fueron capturados empleando jamos de malla plástica tejida, con una abertura de 1mm. Las capturas se realizaron de forma activa desde las márgenes del río. Los individuos fueron trasladados vivos al laboratorio donde fueron sacrificados. Los especímenes fueron medidos (longitud total, LT) utilizando un ictiómetro graduado (mm) y pesados (peso húmedo total, PT) en una balanza analítica de 1 mg de precisión. Posteriormente, se le hizo la disección a cada ejemplar hembra para extraerle las vísceras. Se obtuvo el peso húmedo total sin vísceras (PSv), el peso de la gónada (PG) y el peso del hígado (PH).
Los ejemplares fueron conservados en formol al 4% para su análisis posterior. Seguidamente, se determinó en los peces machos el largo total del gonopodio (LG), utilizando un micrómetro ocular. En el caso de las hembras preñadas se determinó el número de embriones (NE) y el peso de cada uno por separado.
Para la investigación de los isótopos estables se recolectaron peces en mayo del 2007 y fueron conservados en etanol al 70% para su posterior análisis. Se realizó un muestreo preliminar del contenido estomacal. Para ello se escogieron 35 machos de cada sitio distribuidos equitativamente a lo largo del año y se les extrajo el estómago. El contenido estomacal se revisó mediante un microscopio estereoscópico. Cada entidad alimentaria encontrada se clasificó hasta el taxon más bajo posible.
Análisis de datos: Se calcularon los siguientes índices morfométricos y organosomáticos:
Factor de condición:
FC= PT/LT3 (PSv/LT3 en las hembras)
Índice hepatosomático: IHS=(PH/PSv)x100Índice gonadosomático: IGS=(PG/PSv)x100
Índice de fecundidad específica: IFE=NE/PSV
Largo relativo del gonopodio: LRG=(LG/LT) x100
El análisis de los Índices y otras variables se realizó agrupando los individuos según clases de talla (<23 mm; 23-25 mm; 25-27 mm; 27-29 mm; 29-31 mm; 31-33 mm; >33 mm) o peso (<0.3 g; 0.3-0.5 g; 0.5-0.7 g; 0.7-0.9 g; >0.9 g) de acuerdo a cada caso.
Para expresar la composición de la dieta, se utilizó el método de frecuencia de presencia (Fi) definido por Rosecchi y Novaze (1987) como:
Fi=100* fi /n
Donde:
fi: Número de estómagos donde la entidad i fue encontrado.
n: Total de estómagos analizados.
Procesamiento estadístico: Para determinar la significación estadística de las diferencias observadas se seleccionó como método estándar, el análisis de varianza paramétrico de efectos fijos (ANOvA). En los casos en que se encontraron diferencias significativas se aplicó la prueba de comparaciones múltiples de Student-Newman-Keuls (SNK). Previo a la aplicación de los análisis se determinó la no existencia de desviaciones importantes de la normalidad y la homogeneidad de varianza, siguiendo los criterios de Zar (1996) y Underwood (1997). Se transformaron los datos en los casos donde se comprobó que existía una distribución asimétrica o un desvío muy grande de la homogeneidad de varianza de los valores de la variable analizada. En estos casos, los análisis de varianza se realizaron sobre los datos transformados. Tomando en consideración que esta es una especie que se reproduce continuamente, a lo largo de todo el año, no deben esperarse variaciones mensuales marcadas en ninguno de los indicadores analizados. Al finalizar los muestreos y previo a los análisis entre sitios, que es el objetivo central de este trabajo, se realizaron análisis de varianza a las variables consideradas y en ningún caso hubo diferencias entre meses, con lo cual se corroboran los supuestos considerados en el diseño de la investigación. Se utilizó el coeficiente de correlación por rangos de Spearman para el análisis de la similitud en la composición de la dieta entre sitios de muestreo. En todos los casos se fijó un nivel de significación de 0.05. Para el procesamiento de los datos obtenidos se empleó el programa STATISTICA 6.0 para Windows.
Resultados
Pesos y tallas: Se analizaron 2 980 ejemplares, de los cuales 616 fueron hembras, 958 machos y 1 406 indeterminados. Los individuos indeterminados son aquellos cuyo sexo no está definido o desarrollado. Se incluyen en esta categoría, los jóvenes y los animales que por su talla podrían ser hembras pero cuyo sexo no puede ser definido con seguridad.
El análisis de varianza permitió verificar la existencia de interacción significativa entre sitios y sexos con respecto al largo total (F4, 2971=12.177, p<0.001). La SNK (Fig. 1) indica que las hembras de los sitios contaminados son mayores que las hembras del sitio de referencia. Los machos tienen tallas medias similares en todos los sitios y son significativamente menores que las hembras. Los indeterminados tampoco difieren entre sitios y presentan tallas más pequeñas que los individuos con sexo definido.
Al analizar el peso, se encontró la existencia de interacción significativa (F4, 2971=23.830, p<0.001) entre sitios para cada sexo. En el Rodeo predominan las hembras de menor peso con respecto a los restantes sitios de muestreo (Fig. 2). No se encontraron diferencias significativas para los pesos de los machos e indeterminados entre sitios.
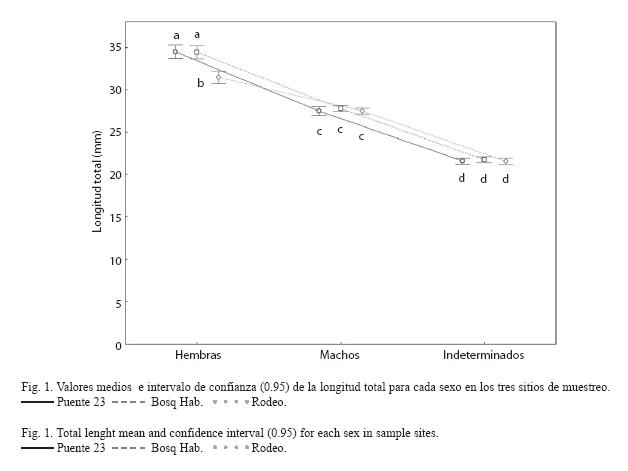
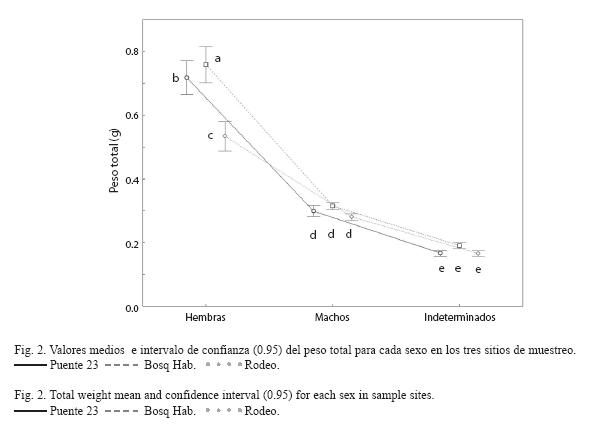
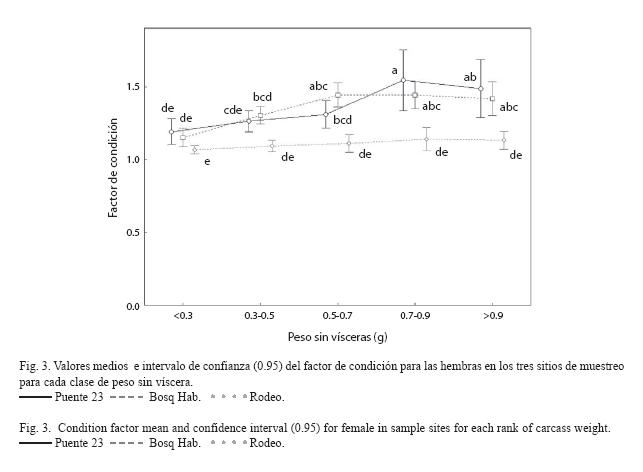
Factor de condición e índice hepatosomático: Al analizar el factor de condición para las hembras se encontró que existe una interacción significativa entre sitios y clases de peso (F8, 599=2.62, p=0.008). Según los resultados de la prueba SNK, los valores medios observados del FC para las hembras mostraron una tendencia a ser mayores en los lugares más contaminados (Fig. 3). Las hembras del Rodeo se caracterizan por presentar un patrón muy similar del FC en todas las clases de peso de los individuos, no siendo así en los otros dos sitios donde se observa un incremento a medida que aumenta el tamaño de los ejemplares.
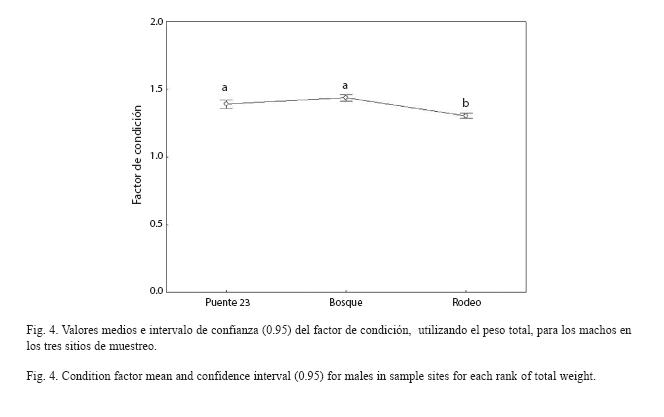
Al analizar el FC para los machos se encontraron diferencias significativas entre sitios (F2,943=17.009, p<0.001). No se encontró interacción significativa entre este factor y las clases de peso (F8,943=0.0019, p=0.910). Según la prueba SNK, el valor medio calculado del FC en los machos resultó significativamente menor en el Rodeo (Fig. 4).
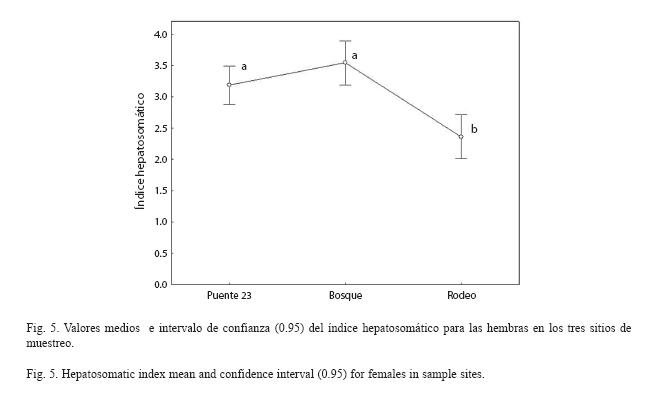
No se encontró interacción significativa entre sitios y clases de peso (F8,447=1.46, p=0.17) al analizar el índice hepatosomático. Se encontraron diferencias significativas entre sitios (F2,447=12.99, p<0.001). Las hembras del Rodeo presentaron valores más bajos que las hembras de los restantes sitios (Fig. 5).
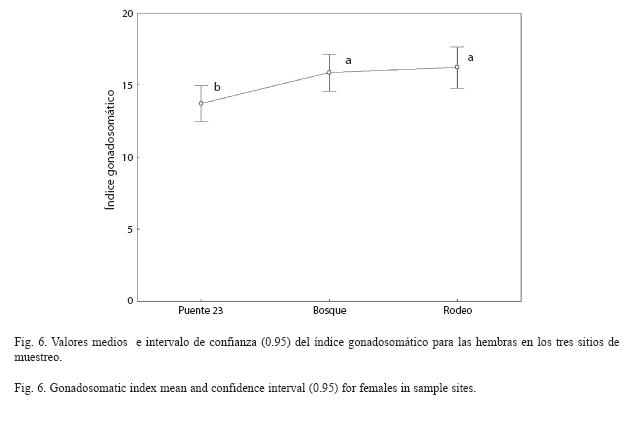
Reproducción: Al analizar el índice gonadosomático, se encontró que no existe una interacción significativa entre sitios y clases de peso (F8,581=0.82, p=0.58). Se encontraron diferencias significativas entre sitios (F2,581=23.62, p<0.001). Al aplicar la SNK obtuvimos que la zona más contaminada, Puente 23, presentó los valores más bajos (Fig. 6).
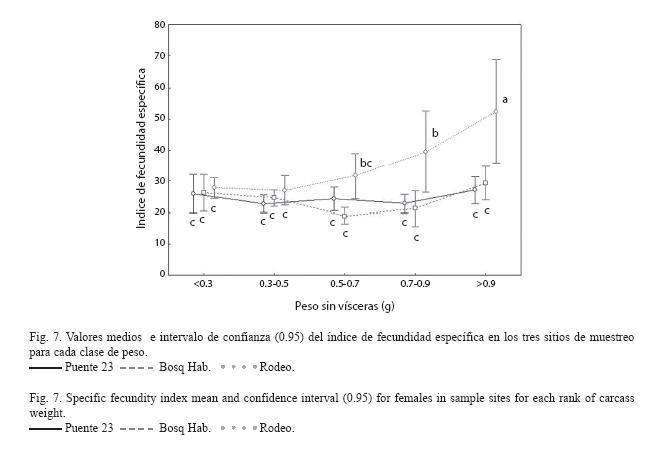
Al analizar el índice de fecundidad específica se encontró que hay interacción significativa (F8,600 =3.33, p<0.001) entre lugares y clases de peso. La prueba SNK (Fig. 7) indica que el valor del IFE es significativamente menor en los sitios más contaminados con respecto al sitio de referencia en las dos clases de peso más altas. Las hembras menores presentan valores medios de IFE muy similares entre sitios.
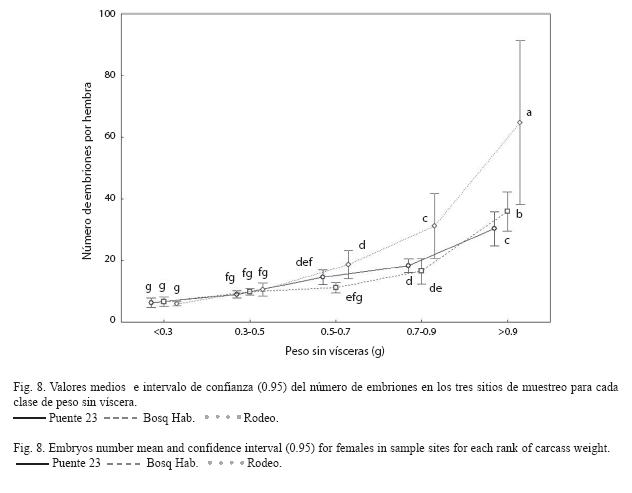
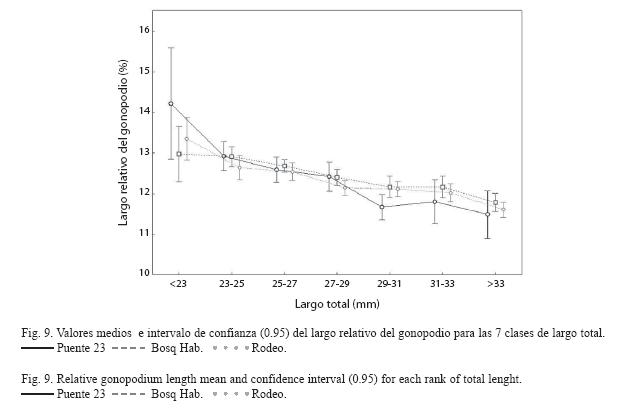
El ANOVA para el número de embriones entre sitios y clases de peso demostró la existencia de una interacción significativa entre los mismos (F8,600 =13.98, p<0.001). Existe una tendencia evidente a ser mayor el número de embriones a medida que el tamaño de la hembra es mayor en los tres sitios. Además se encontró que el número de embriones en el Rodeo es significativamente mayor con respecto a los sitios más contaminados en las clases de peso mayores (Fig. 8). Al analizar el largo relativo del gonopodio se encontró que existe una interacción significativa entre sitios y clases de talla (F12,937=1.94, p=0.03) La prueba SNK definió grupos de medias que se superponen notablemente y no permiten delimitar una tendencia clara que sugiera diferencias entre sitios (Fig. 9).
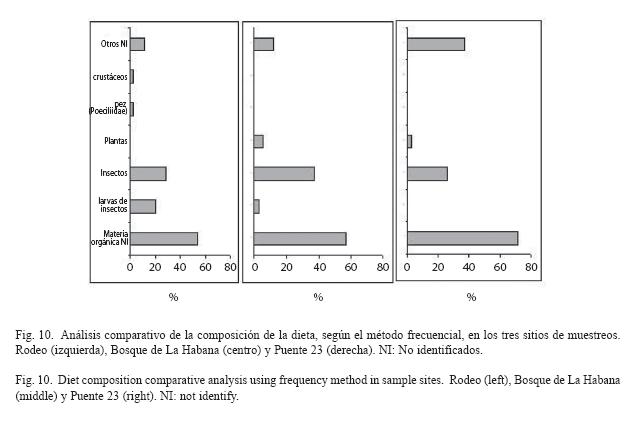
Alimentación: Los componentes de la dieta observados durante el análisis del contenido estomacal fueron agrupados en 7 categorías. En estas se incluyen una amplia gama de organismos, porque su clasificación hasta taxones más bajos es prácticamente imposible debido a la fragmentación o grado alto de digestión en que se encuentran.
La entidad más frecuente en los estómagos analizados corresponde a "insectos" en el sitio Rodeo y Bosque de La Habana. En el Puente 23 se encontró con mayor frecuencia la entidad "otros NI". Se presentaron entidades que sólo fueron observadas en sitios específicos como es el caso de un pez de la Familia Poeciliidae visto en un estómago de un individuo del Rodeo. Lo mismo sucede con la entidad "crustáceos". Se observó una mayor representación de la entidad "larvas de insectos" en el Rodeo. La entidad "plantas" sólo fue observada en los sitios más contaminados (Fig. 10).
Al calcular los coeficientes de correlación de Spearman entre pares de sitios estudiados, se obtuvieron valores no significativos en los pares Rodeo-Puente 23 (Rs=0.56, p=0.190) y Rodeo-Bosque de La Habana (Rs=0.71, p=0.074), mientras que el valor fue alto y significativo entre el Puente 23 y el Bosque de La Habana (Rs=0.93, p=0.002). Esto indica que existe una gran similitud en el orden de importancia de las entidades alimentarias entre el Puente 23 y el Bosque de La Habana, ambos sitios difieren del Rodeo.
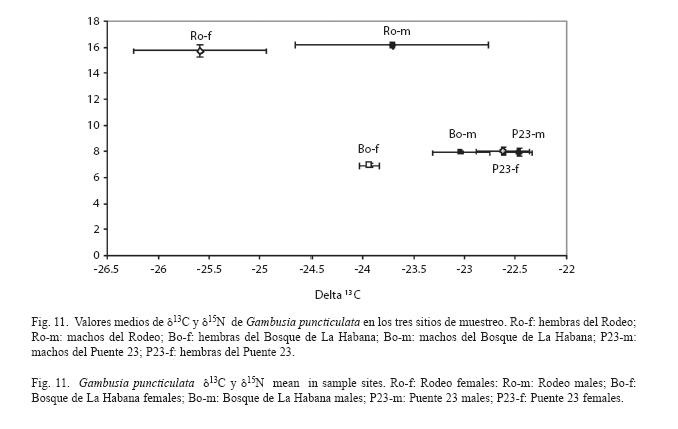
Cociente de isótopos estables: El valor del δ15N está alrededor de 16 para ambos sexos en el Rodeo. En los sitios más contaminados como el Puente 23 y el Bosque de La Habana este valor se encuentra alrededor de 8. En cuanto al δ13C se observaron valores de alrededor de -25.6 para el caso de las hembras de Rodeo, mientras que el resto de los casos presentaron valores entre -24 y -22.5 aproximadamente (Fig. 11).
Al realizar el ANOvA se encontró que el valor δ15N no presentó diferencias significativas entre sexos (F1,26=4.13, p=0.052) y sí con respecto al sitio (F2,26=591.55, p<0.001). La SNK arrojó que esta diferencia es significativamente mayor, para ambos sexos, en el Rodeo en comparación con los otros dos sitios. Los individuos de las zonas contaminadas presentan menor valor de δ15N.
Las diferencias entre sitios =9.85, (F2,26 p<0.001) para los valores medios de δ13C evidenciaron una tendencia bastante clara. Las medias de δ13C fueron significativamente mayores en los sitios de mayor contaminación. El Rodeo presentó un valor relativamente más empobrecido dado fundamentalmente por las hembras de este sitio. Se encontró que existen diferencias en cuanto al sexo (F1,26=6.44, p=0.02). Los machos presentaron los valores mayores.
Discusión
Indicadores morfológicos y reproducción: El resultado más relevante en relación a los indicadores morfológicos es que las hembras colectadas en los sitios contaminados presentan, en general, valores significativamente mayores de talla, peso y factor de condición. Esto pudiera estar relacionado con una mayor disponibilidad de alimento en los sitios más afectados, dada fundamentalmente por la eutrofización. Breitburg (2002) plantea que los efectos del enriquecimiento por nutrientes y eventualmente de la eutrofización dependen en gran medida del balance entre dos aspectos, positivo (más alimento) y negativo (anoxia e incremento de materia orgánica en suspensión) de estos procesos. Estudios de impacto del río Almendares sobre la zona costera mostró que los individuos de la especie Stegastes partitus que habitaban cerca de la desembocadura de este mismo río, presentaron las mayores tallas y pesos (Aguilar y González-Sansón 2000). Toft et al. (2004), por el contrario, observaron una disminución de la talla y del peso en hembras de Gambusia sp en lugares contaminados por descargas de residuales de molinos de pulpa de madera.
Los valores altos del índice hepatosomático en las hembras de los sitios más contaminados pudieran deberse a la naturaleza de la contaminación, la cual es mucho más agresiva en estas partes del río. varios autores (Kime 1998, Porter y Janz 2003, Toft et al. 2003, 2004), han observado un incremento del IHS como consecuencia de la exposición de los organismos a agentes estresantes. Se ha demostrado que el hígado puede aumentar su tamaño en presencia de determinado estrés, particularmente después que el pez ha sido expuesto a la contaminación o a tóxicos específicos (Arcand-Hoy y Metcalfe 1999). El incremento de este órgano puede deberse a la hiperplasia o a la hipertrofia como una respuesta adaptativa para ampliar la capacidad del hígado de desintoxicarse de compuestos extraños (Addison 1984). Tricklebank et al. (2002), por el contrario, han encontrado que algunas fuentes de contaminación pueden conllevar a una reducción de este índice. Se ha demostrado una reducción del IHS en peces expuestos a aguas ácidas (Lee et al. 1983) y a sustancias químicas por ejemplo el cloruro de cadmio (Pereira et al. 1993).
El índice gonadosomático se encuentra influenciado por factores ambientales como son los cambios de fotoperíodo, temperatura, concentración de oxígeno disuelto, niveles de agua, estimulación acústica por la lluvia, disponibilidad de comida y densidad de la población (Burns 1985). En este estudio, el valor más bajo del IGS en el Puente 23, pudiera deberse a que la contaminación de sus aguas es mucho más elevada por recibir la descarga de todo el recorrido del río. Se han reportado disminuciones del IGS en experimentos realizados con distintos contaminantes sobre los peces (Pereira et al. 1993, Kime 1995, Mills et al. 2001). varios autores reportan el efecto negativo de los residuales de molinos de pulpa para papel en el IGS (Munkittrick et al. 1994, Janz et al. 1997, van den Heuvel et al. 2002). Generalmente se asume que, valores del IGS más bajos que lo normal indican un desvío de la energía que habitualmente se usa para el crecimiento de las gónadas. Esta energía es utilizada para combatir un agente estresante de algún tipo (Goede y Barton 1990).
La producción de huevos viables está entre los indicadores más sensibles a la exposición de contaminantes (Suter et al. 1987). Las aguas de los sitios del Puente 23 y el Bosque de La Habana están muy contaminadas, lo cual pudiera contribuir a que la fecundidad específica y el número de embriones por hembra, sea menor en estos sitios con relación al Rodeo. Esto coincide con lo planteado por numerosos autores (McMaster et al. 1991, Adams et al. 1992, Munkittrick et al. 1994, Kime 1995, Janz et al. 1997, Guidetti et al. 2002).
Autores como Toft et al. (2003) utilizan el largo relativo del gonopodio como indicador de la presencia de contaminantes en el agua. Estos autores plantean que una disminución del largo del gonopodio se debe a una alteración de la función endocrina cuando el pez está expuesto a determinados compuestos en el agua. Estos resultados pudieran justificarse a partir de la sensibilidad del gonopodio ante químicos presentes en el agua que actúan como "disruptores endocrinos". En esta investigación no se encontraron diferencias en el tamaño del gonopodio al comparar entre sitios, por lo que no hay evidencia de la presencia de contaminantes hormonalmente activos que afecten de forma directa el sistema endocrino en este sexo.
Alimentación e isótopos estables: Los hábitos alimentarios de la especie estudiada resultaron muy similares a lo descrito por otros autores para otras especies del mismo género en diferentes regiones. Esta especie es básicamente depredadora que se alimenta principal-mente de pequeños invertebrados, aunque en sus estómagos se han encontrado peces y restos de materia vegetal (Fink 1971). Según los resultados, Gambusia puncticulata se pudiera considerar como un depredador oportunistageneralista, debido a la gran variabilidad que presenta en la composición de su dieta. Esto es consistente con lo encontrado por Specziár (2004) para G. holbrooki. La dieta comprende desde artrópodos terrestres (larvas y adultos) como entidad alimentaria fundamental, hasta la fauna y plantas acuáticas como componentes secundarios.
Los tres sitios de estudios se caracterizaron por una vegetación abundante en la ribera del río, lo cual permite el desarrollo de invertebrados terrestres en la zona. Estos fueron frecuentemente encontrados en los estómagos analizados. Bruo y Dozar (1997), plantearon que en zonas como estas existen muchos invertebrados, principalmente insectos, que desarrollan gran parte de su ciclo vital sobre el follaje de los árboles. Esto pudiera ser la causa de un incremento en la abundancia de insectos en los estómagos de los peces analizados en esta investigación.
En este estudio, se investigó solamente la dieta de los machos y este aspecto tuvo un carácter preliminar. No obstante, se encontraron diferencias en la composición del alimento natural entre los sitios más contaminados y el de referencia. Es por ello que este factor debe ser tenido en cuenta a la hora de interpretar la respuesta biológica de los peces.
Las diferencias encontradas para δ13C entre los sitios contaminados y el sitio de referencia sugieren que las fuentes de alimento no son las mismas. Esto está en correspondencia con las diferencias que existen entre las características geomorfológicas y ecológicas de los sitios. Se encontró evidencia de que la fuente de alimentación en los machos puede ser diferente a la de las hembras.
Las diferencias observadas en los valores de δ15N entre sitios indican que las fuentes de nitrógeno en el sitio de referencia están enriquecidas en comparación con los sitios de mayor contaminación. Esto no coincide con lo planteado por numerosos autores que han encontrado que las descargas de aguas albañales tienen mayores valores de δ15N que otras fuentes de nitrógeno (Macko y Ostrom 1994, Heikoop et al. 2000). Cabana y Rasmussen (1996) consignan que los peces en ríos y lagos afectados por descargas de aguas albañales tienen valores más altos de δ15N que los que se desarrollan en aguas prístinas. Por otra parte, algunos estudios realizados en las zonas costeras, reflejan una disminución de los valores de δ15N asociada a la contaminación con aguas de albañal (Rau et al. 1981). Aguilar (2005) plantea la posibilidad de que ciertos tipos de aguas de albañal puedan tener valores relativos de δ15N menores. Esto podría estar asociado con el proceso de tratamiento de las aguas (Gaston y Suthers 2004) o por la influencia de fuentes diversas de nitrógeno.
Lo más importante en este caso, sin embargo, es la existencia de fuentes de nitrógeno diferentes en los sitios contaminados con respecto al sitio de referencia. Es posible que otras actividades humanas, como la agricultura y la ganadería realicen aportes significativos de sustancias nitrogenadas en el sitio del Rodeo y que estas tengan valores de δ15N elevados. Otra causa posible de las diferencias observadas es que los organismos autótrofos tengan marcas isotópicas diferentes para el nitrógeno en los sitios estudiados.
Este trabajo constituye una primera aproximación al conocimiento de los hábitos alimentarios de esta especie y su posible variación por la acción de factores naturales y antrópicos. No obstante, se necesita verificar y ampliar los estudios relacionados con esta temática. Se necesita de un análisis más detallado de la variación natural de las marcas isotópicas (entre años, épocas, y varios sitios) así como, de una investigación más amplia de la dieta natural de esta especie.
Agradecimientos
Agradecemos al Dr. Allen Curry, Director del Canadian Rivers Institute y al laboratorio de isotopos estables de la naturaleza, ambos de la Universidad de New Brunswick, Canada, por su apoyo para el procesamiento de las muestras de isotopos utilizadas en este trabajo.
Resumen
El río Almendares es el más grande de Ciudad de La Habana. La actividad humana adyacente ejerce un impacto significativo. Evaluamos las alteraciones de los procesos biológicos en los peces que viven en sitios muy contaminados de este río. Se aplicó una metodología basada en los efectos acumulativos de agentes estresantes múltiples a nivel de individuo en la especie Gambusia puncticulata (Poey, 1854). Este estudio se efectuó en tres sitios del río Almendares con muestreos mensuales desde julio del 2006 hasta junio del 2007. Se encontró un incremento en el largo y el peso de las hembras, y en el factor de condición para ambos sexos en los sitios más impactados. Este resultado se atribuye a la mayor disponibilidad de alimento debido a la eutrofización. El índice gonadosomático, el índice de fecundidad específica y el número de embriones por hembra, tuvieron valores significativamente más altos en el sitio de referencia. El largo relativo del gonopodio no mostró diferencia significativa entre sitios. El índice hepatosomático presentó valores más altos en las hembras provenientes de los sitios más contaminados. Las diferencias encontradas para δ13C y δ15N entre los sitios contaminados y el de referencia sugieren que las fuentes de alimento no son las mismas.
Palabras clave: río Almendares, contaminación, Gambusia puncticulata, indicadores, Cuba.
Recibido 14-II-2008. Corregido 14-VII-2008. Aceptado 14-VIII-2008.
Referencias
Adams, S.M., W.D. Crumby, M.S. Greeley, Jr., M.G. Ryon & E.M. Schilling. 1992. Relationship between physiological and fish population responses in a contaminated stream. Env. Toxicol. Chem. 11: 1549-1557. [ Links ]
Addison, R.F. 1984. Hepatic mixed functions oxidase induction in fish as possible biological monitoring system, p. 51-60. In P. Hodson & J. Niragu (eds.). Contaminant effects on fisheries. Wiley, Nueva York, EEUU. [ Links ]
Aguilar, C. 2005. La ictiofauna costera de Ciudad de La Habana: Efectos acumulativos de agentes estresantes múltiples en varios niveles de organización biológica. Tesis de Doctorado, Centro de Investigaciones Marinas, Universidad de La Habana, Ciudad de La Habana, Cuba. [ Links ]
Aguilar, C. & G. González. 2000. Influencia de la contaminación de la Bahía de La Habana (Cuba) sobre las asociaciones de peces costeros. 1. Abundancia y diversidad. Rev. Inv. Mar. 21: 60-70. [ Links ]
Arcand, L. & C. Metcalefe. 1999. Biomarkers of exposure of broen bullheads to contaminants in the lower great lakes, North America. Env. Toxicol. Chem. 18: 740-749. [ Links ]
Breitburg, D. 2002. Effects of hypoxia and the balance between hypoxia and enrichment, on coastal fish and fisheries. Estuaries 25: 767-781. [ Links ]
Bruo, V.A. & C.B. Dozar. 1997. Distribution and phonologies of Louisiana Sphingidae. J. Lepidopterists Soc. 51: 156-175. [ Links ]
Burns, J.R. 1985 The effect of low latitude photoperiods on the reproduction of female and male Poecilliopsis gracillis and Poecillia sphenops. Copeia 4: 961-965. [ Links ]
Cabana, G. & J.B. Rasmussen. 1996. Comparison of aquatic food chains using nitrogen isotopes. Proc. Nat. Acad. Sci. USA. 93: 10844-10847. [ Links ]
Chapman, P.M. 2002. Integrating toxicology and ecology: Putting the "eco" into ecotoxicology. Mar. Pol. Bull. 44: 7-15. [ Links ]
Fink, W.L. 1971. A revision of the Gambusia puncticulata complex (Pisces: Poeciliidae). Publ. Gulf. Coast. Res. Lab. Mus. 2: 11- 46. [ Links ]
Gaston, T.F. & I.M. Suthers. 2004. Spatial variation in δ13C and δ15N of liver, muscle and bone in a rocky reef planktivorous fish: the relative contribution of sewage. J. Exp. Mar. Biol. Ecol. 304: 17-33. [ Links ]
Goede, R.W. & B.A. Barton. 1990. Organismic indices and an autopsy based assessment as indicators of health and condition of fish. Americ. Fish. Soc. Sym. 8: 123-144. [ Links ]
Guidetti, P., G. Fanellib, F. Fraschettia, A. Terlizzia & F. Boeroa. 2002. Coastal fish indicate human induced change in the Mediterranean littoral. Mar. Env. Res. 53: 77-94. [ Links ]
Heikoop, J.M., M.J. Risk, A.v. Lazier, E.N. Edinger, J. Jompa, G.v. Limmon, J.J. Dunn, D.R. Browne & H.P. Schwarcz. 2000. Nitrogen-15 signals of anthropogenic nutrients loading in reef corals. Mar. Pol. Bull. 40: 628-636. [ Links ]
Janz, D.M., M.E. McMaster, K.R. Munkittrick & G. Van Der Kraak. 1997. Elevated ovarian follicular apoptosis and heat shock protein-70 expression in the white sucker exposed to bleached kraft pulp mill effluent. Toxicol. Appl. Pharm. 147: 391-398. [ Links ]
Kime, D.E. 1998. Disruption of liver function, p. 201-246. In D. Kime. Endocrine disruption in fish. Kluwer, Boston, Dordrecht, London, Inglaterra. [ Links ]
Kime, D.E. 1995. The effects of pollution on reproduction in fish. Fish Biol. Fish. 5: 52-96. [ Links ]
Lee, R.M., S.B. Gerking & B. Jezierska. 1983. Electrolyte balance and energy mobilization in acid stressed rainbow trout, Salmo gairdneri and their relation to reproductive success. Env. Biol. Fish. 8: 115-123. [ Links ]
Macko, S.A. & N.E. Ostrom. 1994. Pollution studies using stable isotopes, p. 45-62. In K. Lajtha & R.H. Michener (eds.). Stable isotopes in ecology and environmental science. Blackwell Sci., Oxford, Inglaterra. [ Links ]
McMaster, M.E., G.J. van Der Kraak, C.B. Portt, K.R. Munkittrick, P.K. Sibley, I.R. Smith & D.G. Dixon. 1991. Changes in hepatic mixed function oxygenase (MFO) activity, plasma steroid levels and age at maturity of white sucker (Catostomus commersoni) population exposed to bleached kraft pulp mill effluent. Aquat. Toxicol. 21: 199-218. [ Links ]
Mills, L.J., R.E. Gutjarh-Gobell, R.A. Haebler, D.J. Borsay-Horowitz, S. Jayaraman, R.J. Pruell, R.A. McKinney, G.R. Gardner & G.E. Zaroogian. 2001. Effects of estrogenic (o,p´-DDT; octyphenol) and antiandrogenic (p,p´-DDE) chemicals on indicators of endocrine status in juveniles males summer flounder (Paralichthys dentatus). Aquat. Toxicol. 52: 157-176. [ Links ]
Munkittrick, K.R., G.J. Van Der Kraak, M.E. Mc Master, C.B. Prott, M.R. van Den Hewvel & M.R. Serous. 1994. Survey of reciving water environmental impacts associated with discharges from pulp mills. Gonad sixe, liver sixe, hepatic erod activity and plasma six steroid levels in white sucker. Env. Toxicol. Chem. 13: 1089-1101. [ Links ]
Nunes, B., F. Carvalho & L. Guilhermino. 2005. Acute toxicity of widely used pharmaceuticals in aquatic species: Gambusia halbrooki, Artenia parthenogenesis and Tetraselnius chuii. Ecotox. Env. Saf. 61: 413-419. [ Links ]
Olivares, R., S.D. de la Rosa, L. Lima, D. Graham, K. do Alessandro, J. Borroto, F. Martínez & J. Sánchez. 2005. Assessment of heavy metal levels in Almendares River sediments. Havana City, Cuba. Water Research 39: 3945-3953. [ Links ]
Pereira, J., R. Mercalado, C. Kuropat, D. Luedke & G. Senne. 1993. Effects of cadmiun accumulation on serum vitelogenin levels and hepatosomatic and gonadosomatic indices of winter flounder. Env. Toxicol. 24: 427-431. [ Links ]
Porter, C.M & D.M. Janz. 2003. Treated municipal sewage discharge affects multiple levels on biological organization in fish. Ecotox. Env. Saf. 54: 109-206. [ Links ]
Rau, G.H., R.E. Sweeney, I.R. Kaplan, A.J. Mearns & D.R. Young. 1981. Differences in animal 13C, 15N and the abundance between a polluted and an unpolluted coastal site: Likely indicators of sewage uptake by a marine food web. Estuarine, Coastal and Shelf Sci. 13: 701-707. [ Links ]
Rosecchi, E. & Y. Novaze. 1987. Comparaison de cinq indices alimentaires utilisés dans I`analyse des contenus stomacaux. Rev. Trav. Inst. Peches Marit. 49: 11-123. [ Links ]
Specziar, A. 2004. Life history pattern and feeding ecologyof the introduced Gambusia halbrooki in a thermal spa under temperature climate of lake Hévíz, Hungary. Hydrol. 522: 249-260. [ Links ]
Suter, G.W., A.E. Rosen, E. Linder & D.F. Parkhurst. 1987. Endpoints for responses of fish to chronic toxic exposures. Env. Toxicol. Chem. 6: 793-809. [ Links ]
Toft, G., E. Baatrup & L. Guillette, Jr. 2004. Altered social behavior and sexual characteristics in Gambusia halbrooki living downstream of a paper mill. Aquat. Toxicol. 70: 213-222. [ Links ]
Toft, G., T. Edwards, E. Baatrup & L. Guillette, Jr. 2003. Disturbed sexual characteristics in male Gambusia halbrooki from a lake contaminated with endocrine disruptors. Env. Health Persp. 3: 695-701. [ Links ]
Tricklebank, K., M. Kingsford & H. Rose. 2002. Organochloride pesticide and hexachlorobenzene along central coast of New South Wales: multiscale distributions using Parma microlepsis as an indicator. Env. Pol. 116: 319-335. [ Links ]
Underwood, A.J. 1997. Experiments in ecology. Their logical design and interpretation using analysis of variance. Cambrige University, Cambrige, Inglaterra. 504 p. [ Links ]
Van Den Heuvel, M.R., R.J. Ellis, L.A. Tremblay & T.R. Stuthridge. 2002. Exposure of reproductively maturing rainbow trout to a New Zealand pulp and paper mill effluent. Ecotox. Env. Saf. 51: 65-75. [ Links ]
Zar, J.H. 1996. Biostatistical analysis. Prentice Hall, New Jersey, EEUU. 620 p. [ Links ]

