










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.56 n.3 San José Sep. 2008
Mycorrhizal-like interaction between gametophytes and young sporophytes of the fern Dryopteris muenchii (Filicales) and its fungal endophyte
Irma Reyes-Jaramillo1*, Sara Lucía Camargo-Ricalde1 & Ma. de los Ángeles Aquiahuatl-Ramos2
1. Depto. de Biología, 2Depto. de Biotecnología, División de Ciencias Biológicas y de la Salud, Universidad Autónoma Metropolitana-Iztapalapa, México D.F, México. Fax: (+55)-58 04 46 88; irj@xanum.uam.mx
Abstract The morphology of a Glomus-like fungus-host interaction in chlorophyllous gametophytes and young apogamic sporophytes of Dryopteris muenchii A.R. Sm. was studied from ferns cultivated in laboratory, using soil as substrate. An aseptate fungus colonized the gametophytes tissue through the rhizoids, developing vesicles. The fungus penetrated the young sporophytes primary roots by developing appressoria. It spread forming inter- and intra-cellular hyphae through the epidermis and the outermost cortical cell layers, where it formed vesicles, hyphal coils-like and arbuscules. The fungus hyphae never colonized the gametophyte-sporophyte cellular junction. The fungal structures observed on D. muenchii during this study, are rather similar to those reported for the plant host-arbuscular mycorrhizal fungus (AMF) interaction, where the AMF described belonged to Phylum Glomeromycota. Therefore, this study is a contribution to the scarce knowledgement of the association between AMF and chlorophyllous gametophytes and young apogamic sporophytes of ferns. Rev. Biol. Trop. 56 (3): 1101-1107. Epub 2008 September 30.
Key words: Arbuscular mycorrhiza (AM), endophyte, chlorophyllous gametophyte, Dryopteridaceae, endemic fern, Mexico.
Arbuscular mycorrhizal fungi (AMF) are associated with ca. 80-85% of living terrestrial plants (Pirozynski 1981, Dickson 2004), and are considered to be ecologically important for most vascular plants because they improve the uptake of specific nutrients by the host, other functions attributed to the fungi include the promotion of plant growth hormones production, protection of host roots from pathogens, and favors the increase of solubility of soil minerals, among other benefits (Remy et al. 1994, Newsham et al. 1995, Merryweather and Fitter 1996, 1998a, 1998b).
Blackwell (2000) suggested that symbiotic relationships of AMF and the roots of higher plants were essential in moving plants to land. The fossil record shows that some Devonian fossil plants (genera Aglaophyton, Rhynia and Asteroxylon, 400 Myr) already had vesicle-like structures similar to those of Glomus (Pirozynski 1981, Remy et al. 1994), which is consistent with the hypothesis that arbuscular mycorrhizas (AM) were instrumental in the colonization of land by ancient plants (Taylor et al. 1995).
Mycorrhyzal associations are common in the roots of numerous fern species (sporophytic phase) and are documented in several light- and electron microscopic studies (Rayner 1927, Burgeff 1938, Boullard 1957, Fontana 1959, Hepden 1960, Cooper 1976, Mishra et al. 1980, Iqbal et al. 1981, Laferière and Koske 1981, Berch and Kendrick 1982, Gemma and Koske 1990, Ragupathy and Mahadevan 1993, Raja et al. 1995, Schmid and Oberwinkler 1995, Zhi-wei 2000, Muthukumar and Udaiyan 2000, Zhang et al. 2004).
Fern gametophytes (haploid phase) may also be associated with endophytic fungi; in particular, the subterranean achlorophyllous gametophytes within the Pteridophytes, which are frequently infected by fungal endophytes; hence few studies on this matter are found (Bruchmann 1904, 1906, 1908, Campbell 1907, Whittier 1977, Boullard 1979, Peterson et al. 1981, Schmid and Oberwinkler 1993, 1994, 1995, Kovacs et al. 2003, Duckett and Ligrone 2005). In contrast, there is little knowledge about fern chlorophyllous gametophytes associated with endophytic fungi. Campbell (1908) described the AM of some species of Marattiaceae and Gleicheniaceae, and of Osmunda cinnamomea L. Bower (1923) studied the symbiotic interaction in gametophytes of Marattiaceae and Schizaceae. Schmid and Oberwinkler (1995) studied the AM in gametophytes and young sporophytes of Gleichenia bifida (Willd.) Spreng. (=Sticherus bifidus (Willd.) Ching.), and other Gleicheniaceae, and Turnau et al. (2005) worked with Pellea viridis.
Therefore, the aim of this study is to contribute to the knowledge of the AMF-host association by describing the morphology of a Glomus-like fungus-host interaction in chlorophyllous gametophytes and young apogamic sporophytes of Dryopteris muenchii A. R. Sm. (Dryopteridaceae), a narrow endemic Mexican fern species.
Although there are studies of this type in other regions of the world, as far as we know, this is the first one done in Mexico and it is related to the knowledge of the state of D. muenchii wild populations.
Dryopteris muenchii in Mexico
Family Dryopteridaceae comprises 16 genera and ca. 400 species. In Mexico, this family is represented by eight genera: Arachniodes, Didymochlaena, Dryopteris, Olfersia, Phanerophlebia, Polybotrya, Polystichum and Stigmatopteris (Moran 1995). Genus Dryopteris Adans. ("shield fern") occurs in temperate and tropical regions of the world, 300-3050 m.o.s.l., with ca. 225 species. Its highest diversity occurs in Eastern Asia, while only 30 species are found in the New World, 13 of them occurring in Mexico (Mickel and Smith 2004).
According to Mickel and Smith (2004), D. muenchii A.R. Sm. is endemic to Mexico; it has only been collected in the states of Hidalgo and Chiapas; this species is found in cloud forest (2000-2850 m.o.s.l).
Materials and methods
Dryopteris muenchii spores were collected from fertile sporophytes, using paper bags, from the Paraje Bonabil, state of Chiapas, Mexico (5 km on Matzab deviation, San Cristóbal de las Casas-Tenejapa road); voucher specimens are deposited at the Herbario Metropolitano-UAMIZ (A. Mendoza, R-182, R-244). Fern spores were sown, without sterilization, in 16 pots (5 cm in diam.), filled with 30 g of soil collected at 0-30 cm depth. Gametophyte cultures were maintained during nine months under laboratory conditions, inside transparent polyethylene bags for avoiding dehydratation and contamination, and using a 12 h light-12 h darkness photoperiod (artificial light, 75 watts), with a daily mean temperature of 25 °C. Pots were watered every ten days with distilled water. Gametophytes, with and without young sporophytes, were carefully washed with distilled water, and fixed with FAA (formol 5%, glacial acetic acid 5%, ethylic alcohol 45%, and distilled water).
For light microscopy, 200 gametophytes and 250 gametophytes with young sporophytes were cleared with 5% KOH for 15 min and stained with acidic glycerol/trypan-blue 0.08% (Phillips and Hayman 1970, Koske and Gemma 1989). Microphotographs were taken using a light microscope.
Results
D. muenchii gametophytes are chlorophyllous with a cordate-spatulate shape. The midrib, with 3-4 cells thick, projects numerous rhizoids (Figs. 1 and 2). This species is considered apogamic because, in vitro, it does not form archegonia and the sporophytes developed have an apogamic origin. The antheridia are produced in the middle region or in the margins of the prothallia (Fig. 3). From 450 gametophytes examined, 51.6% of the prothallia developed apogamic sporophytes (Fig. 1), 43% were asexual (without antheridia and archegonia), and 5% developed only antheridia without sporophytes. Only 4% of the gametophytes, with or without sporophytes, were mycorrhizal.
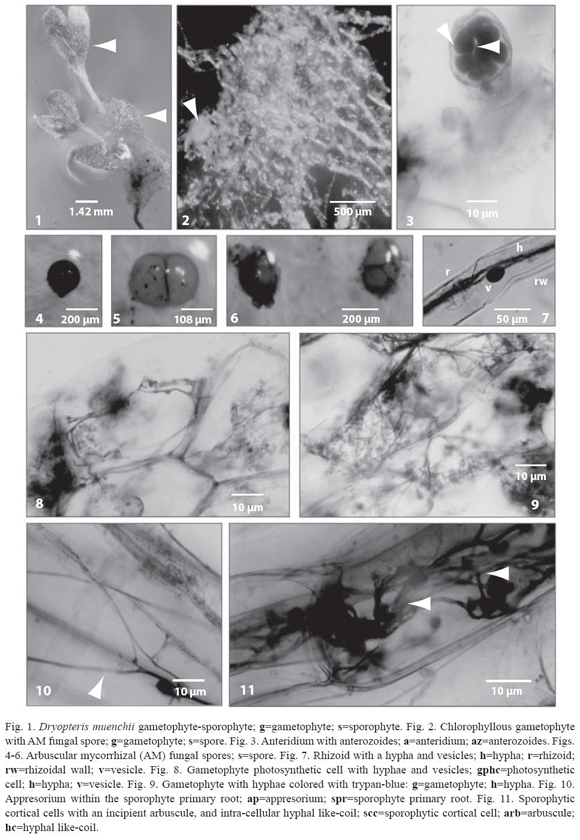
Very few AMF spores were reported amongst the gametophyte rhizoids, the spores were organized in groups of 5, pairs or singles, adhering to each other by common peridial hyphae. AMF spores are globose to subglobose, varying in color from orange- to red-brown (Figs. 4-6).
Gametophyte rhizoids are unicellular and hyaline with 23 µm to 50 µm in diameter. The endophytic fungus enters the host tissue through the rhizoids developing appresoria in the rhizoid wall at the penetration point. Hyphae, 2.5 to 5 µm in diameter, are found inside the rhizoids; these hyphae occur near the rhizoid apex up to the prothallic cells of the gametophyte, where they branch out. Rhizoidal hyphae are aseptate; frequently, these hyphae give rise to oval-shaped vesicles of ca. 15 x 20 µm in diameter, which are densely colored (Fig. 7).
Arbuscular mycorrhizal fungi (AMF) produced inter- and intracellular hyphae on the gametophyte midrib and wings prothallic cells; hyphae branch dispersing over a bigger surface (Figs. 8 and 9).
The meristematic region, placed in the midrib of the gametophyte, and the antheridia are regularly free of infection. The fungus never colonizes the sporophyte foot cells. The primary roots of the young sporophyte are frequently found colonized by a fungal endophyte, morphologically similar to the one described within the gametophyte. The fungus enters the host tissue through the roots developing appessoria in the rhizodermis surface at the penetration point (Fig. 10).
The fungus penetrates the primary roots and spreads forming intra-cellular linear grouped hyphae in the rhizodermis and in the outermost cortical cell layers of the roots, developing hyphal enlargements at the distal hyphal tip, increasing the AMF surface; these structures look like incipient arbuscles (Fig. 11), which colored intensively with trypanblue. Structures similar to hyphal coils were observed, as well, inside the inner root cortex (Fig. 11).
Finally, it is important to mention that the endophitic fungi associated to the young sporophytes, did not infected the gametophyte talus.
Discussion
Under laboratory conditions, D. muenchii is cordiforme, no archegonia are produced, only antheridia were reported; nevertheless, the sporophyte talus has an apogamic origin (Pérez-García et al. 1999, Reyes-Jaramillo and Mendoza 2004).
Apomictic plants are usually adapted to microhabitats, which could explain, in part, the fact that D. muenchii is a narrow endemic fern, and apomixis is also associated with polyploidy, phenomenon rather common in ferns. However, there are no studies relating both AMF and apogamic ferns, in functional and evolutionary terms.
From 450 gametophytes, only 4% of them, with or without sporophytes, were mycorrhizal. The scarce AMF spores, obtained from the pots, closely resemble to those of Glomus globiferum: the spores were organized in groups of 5, pairs or singles, adhering to each other by common peridial hyphae, having a globose to subglobose shape and varying in color from orange- to red-brown (Koske and Walker 1986).
We also found similar fungal structures than the ones found in Gleichenia bifida (Willd.) Spreng by Schmid and Oberwinkler (1995): the endophytic fungal hyphae were aseptate and spread inter- and intra-cellularly, they penetrated the host tissue through the rhizoids, developed vesicles in both the rhizoidal and the prothallic cells.
Within the gametophyte chlorophillic cells, intracellular fungal hyphae were reported; the hyphae branched covering most of the prothallic cells and, in some cases, the hyphae managed to trespass the cell wall, establishing inter-cellular hyphae; however, no arbuscles were registered.
In spite of this, the fungus penetrated the sporophyte primary roots and spread forming intra- and inter-cellular linear grouped hyphae in the rhizodermis and in the outermost cortical cell layers of the roots, developing "arbusculeshaped structures" and hyphal like-coils. The fungus continued to spread by linear or coiled hyphae; this might be considered a fungal adaptation to the limited growth of both the gametophytes and the sporophytes in comparison to the root system of the vascular plants root system.
The fungal structures observed on D. muenchii during this study, are rather similar to those reported for the plant host-AMF interaction, where the AMF described belonged to Glomeromycota. Therefore, this study is a contribution to the scarce knowledgement acquired of the association between AMF and chlorophyllous gametophytes and young apogamic sporophytes of ferns.
Acknowledgments
We thank Rosaura Grether González and Armida Leticia Pacheco Mota, from UAM-Iztapalapa, for the critical review of this manuscript.
Resumen
Se describe la morfología de un hongo endófito afín al género Glomus, como colonizador de gametofitos clorofílicos y de esporofitos apogámicos jóvenes del helecho Dryopteris muenchii A.R. Sm.; el estudio se llevó a cabo con helechos cultivados en el laboratorio y utilizando tierra como substrato. El tejido del gametofito fue colonizado, a través de los rizoides, por un hongo miceliar aseptado, el cual formó vesículas. El hongo logró penetrar las raíces primarias de los esporofitos jóvenes desarrollando apresorios. El hongo se dispersó formando hifas inter- e intra-celulares a través de la epidermis y de la capa de células corticales más externas, donde produjo vesículas, estructuras similares a ovillos y arbúsculos. Las hifas del hongo nunca colonizaron las células de la unión entre el gametofito y el esporofito. Las estructuras observadas durante este estudio en D. muenchii, son muy similares a las de la interacción planta hospedera-hongo micorrícico arbuscular (HMA), en donde el HMA descrito corresponde al Phylum Glomeromycota. Por lo anterior, este estudio es una contribución al escaso conocimiento que se tiene sobre la asociación entre los HMA y gametofitos clorofílicos y esporofitos apogámicos de helechos jóvenes.
Palabras clave: Micorriza arbuscular (MA), endófito, gametofito clorofílico, Dryopteridaceae, helecho endémico, México.
Received 23-II-2007. Corrected 30-XI-2007. Accepted 31-VII-2008.
References
Berch, M. & B. Kendrick. 1982. vesicular-arbuscular mycorrhizae of Soutthern Ontario ferns and fern-allies. Micologia. 74: 769-776. [ Links ]
Blackwell, M. 2000. Terrestrial life-fungal from the start? Science. 289: 1884-1885. [ Links ]
Boullard, B. 1957. La micotrophie chez les ptéridophytes. Sa fréquence, ses caractères, sa signification. Botanist. 41: 5-185. [ Links ]
Boullard, B. 1979. Consideration sur la symbiose fongique chez les Pteridophytes. Syllogeus 19: 1-59 [ Links ]
Bower, F. O. 1923. The ferns. Vol. 1. Cambridge, London, England. [ Links ]
Bruchmann, H. 1904. Úber das Prothallium und die Keimpflanze von Ophioglossum vulgare L. Bot. Zeit. 62: 227-248. [ Links ]
Bruchmann, H. 1906. Über das Prothallium und die Sporenpflanze von Botrychium lunaria Sw. Flora 96: 203-230. [ Links ]
Bruchmann, H. 1908. Das Prothalliun von Lycopodium complanatum L. Bot. Zeit. 66: 169-181. [ Links ]
Burgeff, H. 1938. Mycorriza, p. 159-191 in F. Verdoon. Manual of Pteridology. The Hague, Holland. [ Links ]
Campbell, D.H. 1907. Studies on some Javanese Anthocerothaceae. I. Ann. Bot. 21: 467-486. [ Links ]
Campbell, D. H. 1908. Symbiosis in fern prothallia. Amer. Nat. 42: 154-165. [ Links ]
Cooper, K. M.1976. A field survery of mycorrhizas in New Zeland ferns. New Zealand J. Bot. 14: 169-181. [ Links ]
Dickson, S. 2004. The Arum-Paris continuum of mycorrhizal symbioses. New Phytol. 163: 187-200. [ Links ]
Duckett, J.G. & R. Ligrone. 2005. A comparative cytological analysis of fungal endophytes in the sporophyte rhizomes and vascularized gametophytes of Tmesipteris and Psilotum. Can. J. Bot. 83: 14431456. [ Links ]
Fontana, A. 1959. Ricerche sulla simbiosi micorrizica nelle pteridofite e sui microrganismi normalamente prenti nelle loro radici. Allionia 5: 27-66. [ Links ]
Gemma, J.N. & R.E. Koske. 1990. Mycorrhizae in recent volcanic substrates in Hawaii. Am. J. Bot. 77: 1193-1200. [ Links ]
Hepden, P.M. 1960. Studies in vesicular-arbuscular endophytes. II. Endophytes in the Pteridophyta with special reference to leptosporangiate ferns. Trans.Br. Mycol. Soc. 43: 559-570. [ Links ]
Iqbal, S.H., M. Yousaf & M. Younus. 1981. A field survey of mycorrhizal associations in ferns of Pakistan. New Phytol. 87: 69-79. [ Links ]
Koske, R.E. & J. Gemma. 1989. A modified procedure for staining roots to detect vA mycorrhizas. Mycol. Res. 92: 486-505. [ Links ]
Koske, R.E. & C. Walker. 1986. Glomus globiferum: a new species of Endogonaceae with a hyphal peridium. Mycotaxon. 26: 133-142. [ Links ]
Kovacs, G.M., I. Kottke & F. Oberwinkler. 2003. Light and electron microscopic study on the mycorrhizae of sporophytes of Botrichium virginianum arbuscular structure resembling fossil forms. Plant Biology. 5: 574-580. [ Links ]
Laferière, J. L. & R.E. Koske. 1981. Ocurrence of VA-mycorrhizas in some Rhodes Island Perodophytes. Trans. Br. Mycol. Soc. 76: 331-332. [ Links ]
Merryweather, J. & A.H. Fitter. 1996. Phosphorus nutrition of an obligate mycorrhizal plant treated with the fungicide benomyl in the field. New Phytol. 132: 307-311. [ Links ]
Merryweather. J. & A.H. Fitter. 1998a. The arbuscular mycorrhizal fungi of Hyacinthoides non-scripta. I. Diversity of fungal taxa. New Phytol. 138: 117-129. [ Links ]
Merryweather, J. & A.H. Fitter. 1998b. The arbuscular mycorrhizal fungi of Hyacinthoides non-scripta. II. Seasonal and spatial patterns of fungal populations. New Phytol. 138: 131-142. [ Links ]
Mickel, J.T. & A.R. Smith. 2004. The Pteridophytes of Mexico. Memoirs of the New York Botanical Garden. USA. 88: 1-277. [ Links ]
Mishra, R.R., G.D. Shrama. & A.R. Gatphoh. 1980. Mycorrhizas in the ferns of North Eastern India. Proc. Indian Acad. Sci. 46: 546-551. [ Links ]
Moran, R.C. 1995. Dryopteridaceae, p. 210-226 In R.C. Moran & R. Riba. Flora Mesoamericana: Psilotaceae to Salviniaceae. Instituto de Biología (UNAM), Missouri Botanical Garden & the Natural History Museum, London, England. [ Links ]
Muthukumar, T. & K. Udaiyan. 2000. Vesicular arbuscular mycorrhizae in pteridophytes of western ghats, South India. Phytomorphology. 50: 132-142. [ Links ]
Newsham, K.K., A.H. Fitter & A.R. Watkinson. 1995. Multifunctionality and biodiversity in arbuscular mycorrhizas. TREE. 10: 407-411. [ Links ]
Pérez-García, B., A. Mendoza, I. Reyes-Jaramillo & R. Riba. 1999. Morfogénesis de la fase sexual de seis especies mexicanas de helechos del género Dryopteris (Dryopteridaceae). Rev. Biol. Trop. 47: 69-81. [ Links ]
Peterson, R.L., M.J. Howarth & D.P. Whittier.1981. Interactions between a fungal endophyte and gametophyte cells in Psilotum nudum. Can. J. Bot. 59: 711-720. [ Links ]
Phillips, J.M. & D.S. Hayman. 1970. Improved procedures for clearing roots and staining parasitic and vesiculararbuscular mycorrhizas fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 55: 158-161. [ Links ]
Pirozynski, K.A. 1981. Interactions between fungi and plants through the ages. Can. J. Bot. 59: 1824-1827. [ Links ]
Ragupathy, S. & A. Mahadevan. 1993. Distribution of vesicular-arbuscular mycorrhizae in the plants and rhizosphere soils of the tropical plants. Tamil Nadu, India. Mycorrhiza 3: 123-136. [ Links ]
Raja, P., S. Ragupathy & A. Mahadevan. 1995. Arbuscular mycorrhizal association of pteridophytes of Nilgiris and Kodaikanal hills, South India. Acta Botanica India 23: 181-186. [ Links ]
Rayner, M.C. 1927. Mycorriza. New Phytol. 26: 22-45. [ Links ]
Remy, W., T.N. Taylor., H. Hass & H. Kerp. 1994. Four hundred-million-year-old vesicular arbuscular mycorrhizae. Proc. Natl. Acad. Sci. USA 91: 11841-11843. [ Links ]
Reyes-Jaramillo, I. & A. Mendoza. 2004. Apogamia en Dryopteris muenchii (Dryopteridaceae). Polibotánica 18: 99-110. [ Links ]
Schmid, E. & F. Oberwinkler. 1993. Mycorriza-like interaction between the achlorophyllous gametophyte of Lycopodium clavatum L. and its fungal endophyte studied by light an electron microscopy. New Phytol. 124: 69-81. [ Links ]
Schmid, E. & F. Oberwinkler.1994. Light- and electromicroscopy of the host-fungus interaction in the achorophyllous gametophyte of Botrichum lunaria. Can. J. Bot. 72: 182-188. [ Links ]
Schmid, E. & F. Oberwinkler. 1995. A light- and electron-microscopic a vesicular-arbuscular host-fungus interaction in gametophytes and young sporophytes of the Gleicheniaceae (Filicales). New Phytol. 129: 317-324. [ Links ]
Taylor, T.N., W. Remy, H. Hass & H. Kerp. 1995. Fossil arbuscular mycorrhizae from the early Devonian. Mycologia. 87: 560-573. [ Links ]
Turnau. K., T. Anielska & A. Jurkiewicz. 2005. Mycothallic/ mycorrhizal symbiosis of chlorophyllous gametophytes and sporophytes of a fern, Pellaea viridis (Forsk.) Prantl (Pellaeaceae, Pteridales). Mycorrhiza. 15: 121-128. [ Links ]
Whittier, D.P. 1977. Gametophytes of Lycopodium obscurum grown in axenic culture. Can. J. Bot. 55: 563-567. [ Links ]
Zhang, Y., L.-D. Guo & R.-J. Liu. 2004. Arbuscular mycorrhizal fungi associated with common pteridophytes in Dujiangyan soutwest China. Mycorrhiza. 14: 25-30. [ Links ]
Zhi-wei, Z. 2000. The arbuscular mycorrhizas of pteridophytes in Yunnan, southwest China: evolutionary interpretations. Mycorrhiza. 10: 145-149. [ Links ]

