










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.56 n.2 San José Jun. 2008
Stem anatomical characteristics of the climbing palm Desmoncus orthacanthos (Arecaceae) under two natural growth conditions in a tropical forest
Joaquín Quiroz1, Roger Orellana1, Gabriel Canto1, Silvia Rebollar2 & Pedro Herrera-Franco1
1. Centro de Investigación Científica de Yucatán, Calle 43, No. 130, Col. Chuburna de Hidalgo, C.P. 97200, Mérida, Yucatán, México; cenciart@yahoo.com
2. Departamento de Biología, Universidad Autónoma Metropolitana, México D.F, México.
Abstract: Desmoncus orthacanthos is a Neotropical climbing palm that resembles rattan and therefore has similar potential applications. The genus Desmoncus (subfamily Arecoideae, subtribe Bactridinae) is distributed throughout the Americas, from veracruz, Mexico, to Brazil and Bolivia. The anatomical characteristics of its support tissue have not been thoroughly studied, although some observations from Central American artisans suggest that the stems collected from undisturbed sites possess better characteristics; these include a good capacity to withstand bending without breaking (i.e. higher fracture strength than plants from disturbed sites). Stem samples were collected from individuals from disturbed and undisturbed sites, at three points along the length of the stem (basal, medium and apical). Collections were made of one ramet from five individuals (n=5) at both sites. Each ramet was divided into three sections: basal, from soil surface to a height of 0.5 m; medium, from a height of 0.5 to 5.0 m; and apical, from a height 5.0 to 10.0 m. An anatomical analysis including vascular bundles, parenchyma elements and fibers was performed in the radial direction and also along the longitudinal direction of the stems. The amount of vascular bundles was greater for samples from undisturbed site stems; the amount of parenchyma cells differ between samples from both sites and the amount of fibers was greater for samples from disturbed site stems. The anatomical structural dimensions were smaller for samples from the undisturbed site stems. These findings partially confirm the artisans belief and supports the conclusion that microclimatic conditions affect plant anatomical structure. Rev. Biol. Trop. 56 (2): 937-949. Epub 2008 June 30.
Key words: Desmoncus orthacanthos, stem anatomy, climbing palms, growth conditions.
Desmoncus orthacanthos Martius is a Neotropical climbing palm (Uhl and Dransfield 1987, Henderson et al. 1995) with clonal growth, whose stems have potential applications for furniture and handcraft similar to those of Asian rattans (Phillips 1991, Quero 1992, Henderson and Chávez 1993, Orellana et al. 1999, Siebert 2000, Escalante et al. 2004). The genus Desmoncus (subfamily Arecoideae, subtribe Bactridinae) is distributed throughout the Americas, from veracruz, Mexico, to Brazil and Bolivia (Uhl and Dransfield 1987, Henderson et al. 1995). It consists of climbing species with sympodial habits in which every ramet develops from a rhizome (Hallé et al. 1978, Uhl and Dransfield 1987). The populations of D. orthacanthos grow along the edges of forests and gaps, both in disturbed and undisturbed sites, as well as in areas near the coast (Henderson et al. 1995); it tolerates fire and/or seasonal floodings (Orellana 1992).
In the literature, only a few studies about the anatomical characteristics of D. orthacanthos stem tissues can be found (Orellana et al. 1999, Tomlinson and Zimmermann 2003, Rowe et al. 2004, Isnard et al. 2005). Many reports about the general behavior of climbing species can be found in the literature and it is known that they find a mechanical support on another plant or external object but are rooted in the soil (Ewers and Fisher 1991, Putz and Mooney 1991, Tyree and Ewers 1996). There are also several studies about the effect of changes in the microenvironment or the operational environment (sensu Salisbury and Ross 1992) on the plant growth and form (Hutchinson and Matt 1977, Begon et al. 1989), and that plants adjust to variations of their environment (Rich 1986, Tilman 1988, Bazzaz 1999, Jaffe et al. 2002). However, little is known about the effect of the operational environment on the dimensions and density of the anatomical structure (Arnold and Mauseth 1999), which determines the stems mechanical properties (Escalante 1996, Orellana et al. 1999). Changes in the microenvironment generate phenotypical responses at the anatomical level that could determine the mechanical characteristics of the plant organs. Rowe et al. (2004) described the mechanical properties as a response through changes in the anatomical structure of the stem. Therefore, differences in the mechanical properties of the stem of D. orthacanthos are readily observed by differences in anatomical organization. They confirmed that highly dense bundles with thick-walled fibers are distributed at the periphery of the stem, and the number of bundles and their density decrease toward the center of the stem cross-section.
In this study the degree of variation in anatomical characteristics in D. orthacanthos stems, growing at two different environmental conditions will be presented. It is assumed that stems growing in the different conditions exhibit significant differences in their anatomical structures. This assumption arises from direct observations from artisans in various Neotropical regions who state that differences on the rigidity of the stems and their strength depends on the original place where they grew up (Schultes 1940, Balick 1979, Chinchilla 1993, Chinchilla 1994, Belsky and Siebert 1998, Marmillod and Gálvez 1998, Orellana et al. 1999).
This study compares the anatomical structure, the diameter of vascular bundles (metaxylem, protoxylem, and the sheath of sclerenchyma), the diameter and number of fibers of the sheath, and the classification type sensu Weiner and Liese (1990, 1993) as well as the number of cells in each field of view for the fundamental parenchyma taken from the central cylinder of stems of D. orthacanthos. These stems grew at undisturbed and disturbed sites within evergreen and semi-evergreen tropical forests sensu Olmsted et al. (1999). The a priori expectation was that significant differences would exist in the stem anatomical structural sizes, as a response from the environmental characteristics of each growth site.
Materials and methods
Samples were collected from the original and secondary forests at Ejido Noh Bec in the state of Quintana Roo (20°22 N, 87°26 W; 19°03 N, 88°43 W), located on the Yucatan Peninsula, Mexico. The reference copies were deposited in the herbarium at CICY with the following registrations: Orellana, Castillo y Quiroz, # 960 and 961; Orellana, # 965 bis; Orellana y Quiroz, # 986.
The undisturbed site is a local reserve of evergreen and semi-evergreen tropical forest with a canopy height from 25 to 35 m that had not been cleared as a result of anthropogenic activities for approximately 80 years, based on local information from the "elder ejidatarios". The disturbed site was a patch of the same original vegetation that had been used for temporary agriculture and had been in recovery for approximately 8 years at the time of sample collection; the canopy in this area was approximately 12 to 15 m high. Table 1 summarizes some environmental average characteristics of both sites.
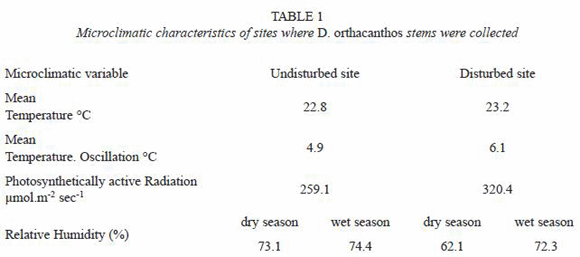
One ramet from five different individuals (n=5) were collected at both sites. Each ramet was divided into three sections: basal, from soil surface to a height of 0.5 m; medium, from a height of 0.5 to 5.0 m; and apical, from a height of 5.0 to 10.0 m (Fig. 1). Three segments from sections measuring 3 cm long were cut from the medium area of the internode and were processed as follows: i) fixing in glycerin-alcohol-water solution 3:2:1; ii) removal of silica deposits with 10 % fluorhydric acid; iii) washing in running water for 48 h; iv) elimination of the wax layer and the outer layer from the samples; v) softening of the stem with 10 % ethylendiamine; vi) washing three times with distilled water under reflux until boiling to remove the softener; vii) dehydration with an alcohol series (30 %, 50 %, 70 %, 85 %, 96 % and absolute anhydrous-alcohol); viii) embedding with glycol methacrylate; ix) sectioning of 7µ thickness samples with a rotating microtome (MICROM mod. 325) for microscopic observations; x) mounting of the samples on slides ; xi) contrast dying with rapid green-safranin at three time periods of 45 seconds each, and washing in distilled water between dying times; xii) mounting with xylol 3:1 (Johansen 1974, Barajas and Echenique-Manrique 1976, Berlyn and Miksche 1976, Gaviño et al. 1979, Rich 1987, Carson 1997, Ruzin 1999).
Optical viewing and measurement of the anatomical structures was performed with a projector (6x) using transversal cuts. The counting was done on a projection for each section (i.e. basal, medium, and apical) using five fields of view each of 2.54x10-4 µm2. A 100:1 micrometric ruler was used as a reference for the measurements. Measurements of the dimensions of 500 parenchyma cells, 500 fibers and 150 vascular bundle units were performed in the radial and longitudinal orientations.
The anatomical descriptions were done using the criteria from Klotz (1978), Tomlinson (1990), and Tomlinson and Zimmermann (2003) for the classification of the vascular bundles according to the phloem position. For the parenchyma type, the classifications of Liese and Weiner (1989, 1990) and Weiner and Liese (1990, 1993) for vascular bundles of rattans species were used.
Microscopic observations were done using a scanning electron microscope (JEOL JSM-6360 LV). The samples were prepared by first dehydrating the stem tissue and then cutting it into cubes. These cubes were placed in a vacuum chamber and covered with a gold layer. Then, each cube was mounted on adhesive tape and placed on a 10 mm aluminum sampleholder plate.
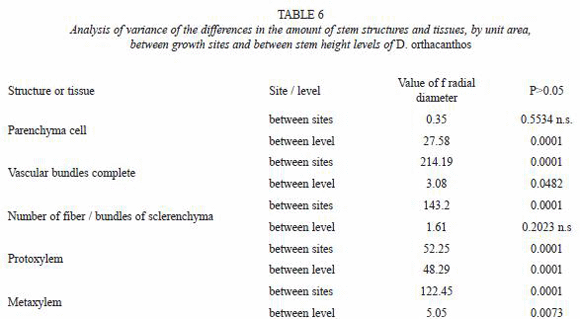
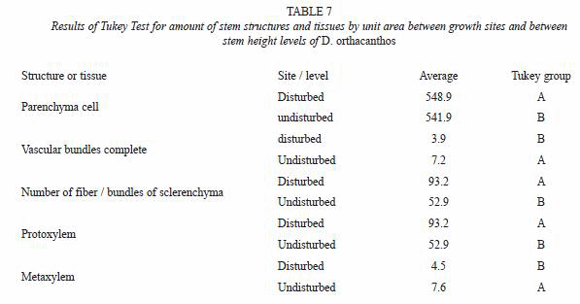
The results from the measurements of the stem anatomical structure were codified by origin site (disturbed vs. undisturbed) and section (basal, medium, apical). The data were analyzed with commercial statistical analysis software called Statistical Analysis System (SAS) (SAS Institute Inc., 1998). An analysis of variance (ANOVA) was performed to verify the existence of statistically significant differences under the assumption of equality. Tukey mean tests were also used to identify differences between means and grouping formation, assigning letters (A, B) to each group to indicate differences between them (Sokal and Rohlf 1980).
Results
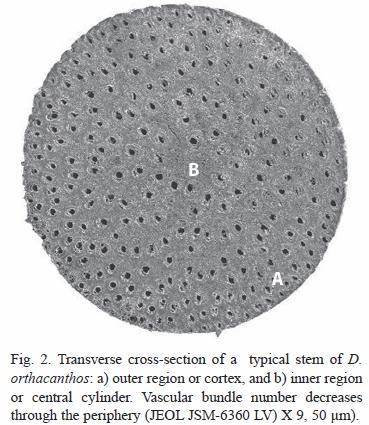
Palms generally have a cylindrical stem, and the area of the transversal cross-sections of the stem, cut from the epidermis to the central cylinder, reveal three distinct zones. The first is a peripheral zone consisting of the epidermis and sub epidermis. The second is an internal zone described by Tomlinson (1990) as the "cortex" or "cortical area", which consists of reduced, dense and highly lignified vascular tissue with fibers. The cortex occupies a small section of the stem. The third section is the central cylinder, which is divided in turn into four regions: periphery, subperiphery, pericentral, and central regions (Tomlinson 1990). The central region consists of a complex of disperse vascular bundles, and each of them is formed by the xylem, phloem and a sheath of fibrous tissue or sclerenchyma, all imbedded in a matrix of ground parenchyma (Tomlinson and Zimmermann 2003).
The cross-sections of all samples at each level of the D. orthacanthos specimens analyzed
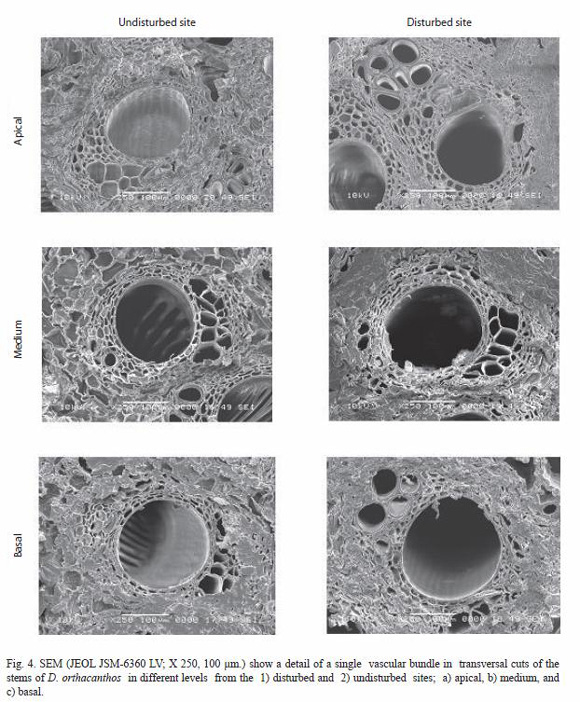
here, from both, the undisturbed and disturbed sites showed a good agreement of the vascular bundle structures with the description given above. A typical vascular bundle consisted of xylem, phloem, and a fiber sheath or sclerenchyma embedded in a matrix of groundparenchyma. The amount of vascular bundles per unit area decreased from the periphery (A) towards the central cylinder (B) (Fig. 2). The vascular bundles have large metaxylem vessels with diameters from 189.7 to 238.5 µm. Protoxylem vessels are found below, at the opposite end of the sclerenchyma. The phloem occurred in a single cluster per vascular bundle, as stated by Weiner and Liese (1990, 1993), and was immersed in the sclerenchyma sheath with the shape of an irregular bunch. The ground parenchyma with diameter from 42.3 to 57.7 µm was formed of tabular cells without intercellular spaces in an elongated, round shape that could be classified as Type C sensu Weiner and Liese (1990, 1993). The sclerenchyma fibers with diameters ranging from 272.4 to 365.2 µm, were arranged as a semicircular sheath over the phloem.
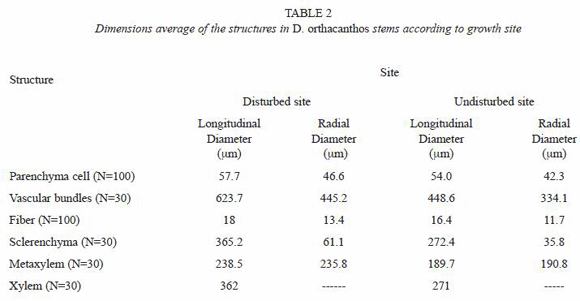
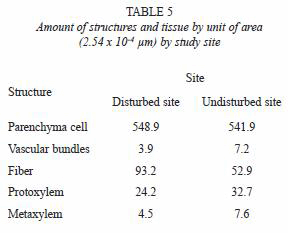
The dimensions of the vascular bundle (i.e. metaxylem and protoxylem, sclerenchyma sheath, parenchyma and fibers) were bigger for the disturbed site stem samples than for those from the undisturbed site (Tables 2 and 3). The amount of vascular bundles per field (2.54x10-4 µm2) was greater in the undisturbed site stems, as shown in Fig. 3 and table 4, while the number of fibers in the sclerenchyma sheath was higher of fibers in the sclerenchyma sheath was higher for the disturbed site stems. The number of parenchyma cells per field (2.54x10-4 µm2) exhibited no significant differences between stems from the two sites (Table 5).
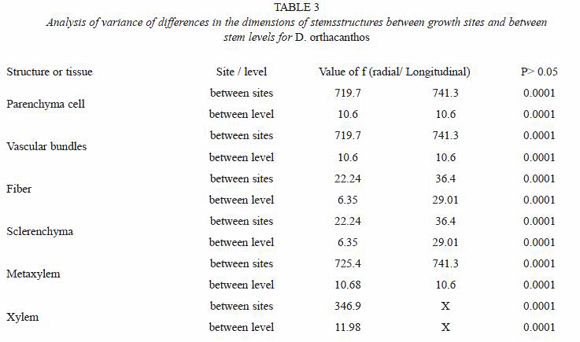
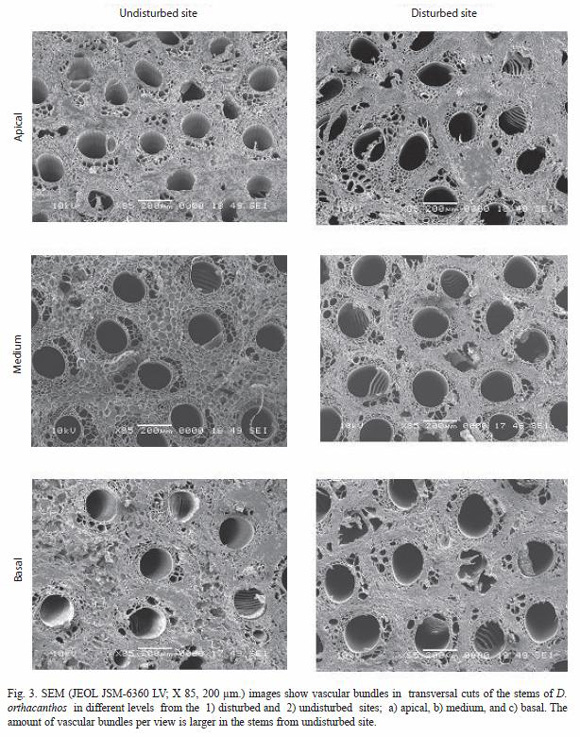
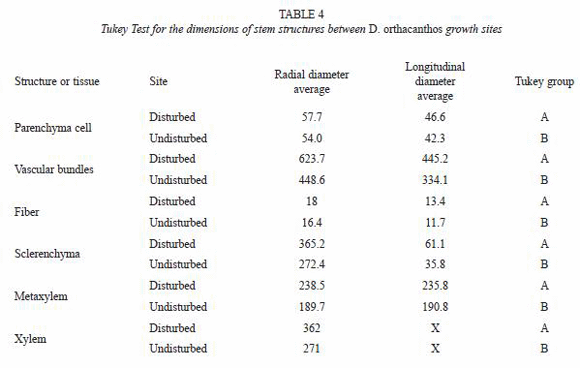
The ANOVA (Table 6) and Tukey test from (Table 7) showed no significant differences in the amount of parenchyma cells between the two sites. However, the vascular bundles and their components were significantly more abundant, although smaller in size, for the undisturbed site stems than for those from the disturbed site stems.
Discussion
The tissue distribution in the D. orthacanthos stem cross-sections was similar to that described for other palm species (Rich 1986, Tomlinson 1990, Liese and Weiner 1989, Weiner and Liese 1990, 1993, Tomlinson and Zimmermann 2003). There is a peripheral zone formed by the epidermis and a sub epidermis (Tomlinson 1990) followed by an interior zone consisting of a peripheral area or "cortex" in which the vascular tissue was densely concentrated, smaller in size and highly lignified. The central cylinder consisted of vascular bundles embedded in parenchymatous tissue, and these bundles gradually increased in size but their number per unit area decreased towards the center of the cylinder.
The vascular bundle structure of stems from both collection sites was similar, and their components had the same structural pattern: the location of the protoxylem and metaxylem, the same arrangement of the phloem like a bunch surrounded by the semicircular sclerenchyma sheath, formed of fibers. The parenchyma in stems from both sites was Type C in the Weiner and Liese (1990) classification scheme.
There were a number of differences between stems samples from the undisturbed and disturbed site. The vascular bundle and fundamental parenchyma sizes were larger for the disturbed site stems than those from the undisturbed site stems suggesting an environmental effect of a parameters such as the light regime on the D. orthacanthos ramet development; measurements of the mean PAR for the undisturbed site resulted in an average value of 259.1 µMol m-2s-1 while for the disturbed site is 320.4 µMol m-2s-1. The vascular bundle size was also smaller and the bundles more abundant in undisturbed site stems (7.2 bundles per view field) than in disturbed site stems (3.9 bundles per view field) as shown in Fig. 3 and 4. As a result, stems from the undisturbed site had a more homogeneous distribution, which are more resistant to biomechanical flexion (live stems) and breaks less easily of the tree branches the D. orthacanthos (Table 8).

To corroborate this assumption, the aver age of the flexural and compressive strength for both mechanical (harvested and processed stems) and biomechanical (with alive stems) were performed, showing a significant difference between the samples from both sites.
The biomechanical flexural strength was higher for the stems from the undisturbed place, attributed to a larger density of vascular bundles, (Table 6). These results explain the empirical observations from the artisans that were mentioned above. However after some treatments of drying and boiling of the stems, the flexural strength (mechanical) is higher for the stems from the disturbed place. This can be explained partially by the structural changes during the treatments carried out on the materials (Quiroz et al., in prep.).
The compressive strength (biomechanical test) was higher for the stems from the disturbed place, and this is considered a result of a higher density of parenchyma cells. As shown in table 6, the parenchyma possesses high water content, which contributes to the biomechanical properties of organisms (Niklas 1992). The results of compression tests (mechanical tests) were similar to the results obtained for the compression tests (biomechanical tests). During the drying process and the thermal treatment, the water of the cells is substituted by oil, which contributes partially with the mechanical properties (Silitonga 1989).
The dimensions of the vascular bundles were significantly different between the stems from the two sites, with the bundles generally being larger for those from the disturbed site stems (Tables 2, 3, 5 and 6). This highlights the close relationship between microclimatic conditions such as temperature, humidity, and PAR (Table 1) at growth sites and the anatomical characteristics. The former determine the size and number of structures and/or tissues per unit area, and consequently define the stem mechanical properties.
The vascular bundles have diverse functions: the xylem transports water and nutrients as well as contributing to the mechanical support; the parenchyma mitigates external pressures with mechanical properties that depend on its dimensions and water content (Ewer et al. 1989, Niklas 1992, Zimmerman et al. 1995); and the sclerenchyma is vital to resist twisting, high pressure and stretching. This suggests that the more abundant vascular bundles in the undisturbed site stems allow for greater flexibility and strength. These functions depend on the amount of bundles per unit area and their distribution, as well as wall thickness and lumen diameter of the tissues (xylem, protoxylem and sclerenchyma) (Rich 1986, Orellana et al. 1999).
The metaxylem diameters observed in this study (238.5 µm for disturbed site stems; 189.7 µm for undisturbed site stems) differed from those reported by Tomlinson and Zimmermann (2003). These authors reported metaxylem diameters of 39 µm in the periphery, 98 µm in the subperiphery, 127 µm in the pericentral and 155 µm in the central all in botanical garden specimens: D. orthacanthos from Cayenne, French Guyana; D. chinantlensis from Fairchild Tropical Garden, Florida; and D. prunifera from Hawaii (the latter two are D. orthacanthos synonyms, as mentioned in Henderson et al. 1995). However, the diameters reported here are similar to the 280 µm diameters described by Klotz (1978) for climbing palms (including D. orthacanthos). The differences between D. orthacanthos and the analogous palms may be due to differences in the growth conditions, as it occurred in the present study, but the size of radial diameter and the longitudinal direction, as well as the number of anatomical structures per field view in D. orthacanthos may also be explained by differences among site variations in the microclimatic conditions.
Although, as stated by Tomlinson and Zimmerman (2003), D. orthacanthos stems cannot be used commercially in the same way as Asian rattans because the parenchyma in the stems maintains a thin, unlignified wall and this may be closely linked to growth environment. Therefore, it is quite possible that D. orthacanthos specimens exist with a thicker parenchyma cell walls and that lignification increases with age, as Rich (1986) mentioned for other palm species.
The anatomical structures of the D. orthacanthos stems studied here exhibited significant variation in response to the growth conditions. Plants from the undisturbed site, a shady environment, must assign more resources to the production of support and elongation structures to reach the higher canopy and access the light they need to produce reproduction structures. At the disturbed site, in contrast, the plants assign their resources to increasing stem diameter, as indicated by the size of the parenchyma cells. Therefore, selection of D. orthacanthos plants for different applications is limited by their environmental conditions since these determine anatomical structure tissue dimensions.
Based on our results we can conclude the following:
1. The central cylinders of the disturbed site plants were generally larger in diameter as a result of their larger vascular bundles (metaxylem, protoxylem and fiber sheath) and parenchyma. However, these plants had fewer vascular bundle units per unit area than the undisturbed site plants.
2. The higher vascular bundle density and quantity per unit area in the undisturbed site plants made them more flexible and resistant than those from the disturbed site, which were more rigid and brittle.
3. The quantity of vascular bundles per unit area in the stem either increases or decreases its strength. This affects an individual plants probability of survival because if the branch breaks it would lose the opportunity to reach the canopy and eventually reproduce. The results confirm that changes in the operational environment (sensu Salisbury and Ross 1994) generate anatomical phenotypical responses that determine the mechanical characteristics of vegetal tissue.
Acknowledgments
The authors thank Felipe Barredo for his help with histological techniques, Ernesto Reyes for support during the statistical analysis, Lilia Can for her assistance in generating the scanning electron microscopic images and Silvia vergara for the drawing of Fig 1. This research was partially supported by the Sistema Regional CONACYT-SISIERRA through the project "Propiedades fisicomecánicas y biomecánicas de cuatro especies de palmeras con potencial como sustitutas del ratán" and the undergraduate thesis of Gabriel Canto, and by the CONACYT through a Ph.D. scholarship (#96144) granted to Joaquin Quiroz.
Resumen
Desmoncus orthacanthos es una palmera trepadora neotropical que puede, potencialmente, utilizarse en usos similares a los del ratán. El género Desmoncus (subfamilia Arecoideae, subtribu Bactridinae) se distribuye en América desde Veracruz, Mexico, hasta Brasil y Bolivia. Esta especie posee características anatómicas que no han sido ampliamente estudiadas pero observaciones hechas por artesanos centroamericanos sugieren que los tallos de sitios no perturbados por actividades humanas presentan mayor resistencia al doblado y menos tallos fracturados en el manejo. Las muestras de tallos fueron recolectadas de individuos que crecen en un sitio conservado y uno perturbado por actividades humanas, a tres alturas de la longitud del tallo (basal, media y apical). Se recolectó una rama de cada uno de cinco individuos (n=5) en ambos sitios. Cada rama fue dividida en tres secciones: basal, desde el nivel del suelo hasta los 0.5 m; medio, desde 0.5 a 5.0 m; y apical, desde 5.0 a 10.0 m. Se midió la cantidad por unidad de área y las dimensiones o tamaño (i.e. diámetro radial y diámetro perpendicular) de paquetes vasculares, células de parénquima y fibras. La cantidad de paquetes vasculares fue mayor en los tallos del sitio conservado, la cantidad de células de parénquima fue diferente entre sitios y la cantidad de fibras fue mayor en los tallos del sitio perturbado por actividades humanas. Estas características muestran cierta relación con los resultados mecánicos en otro estudio de los autores, los cuales confirman parcialmente las observaciones de los artesanos, lo que puede estar estrechamente relacionado con las características microclimáticas de los sitios de crecimiento.
Palabras clave: Desmoncus orthacanthos, anatomía del tallo, palmas trepadoras, condiciones de crecimiento.
References
Arnold, D.H. & J.D. Mauseth. 1999. Effects of environmental factors on development of wood. Am. J. Bot. 86: 67-371. [ Links ]
Balick, M.J. 1979. Economic botany of the Guahibo. 1. Palmae. Econ. Bot. 33: 361-376. [ Links ]
Barajas, M.J. & R. Echenique-Manrique. 1976. Anatomía de maderas de México. No. 1: 12 especies de Jalisco & Veracruz. Instituto de Investigaciones de Recursos Bióticos 1: 29-70. [ Links ]
Bazzaz, F.A. 1999. Plants in changing environments: linking physiological, population and community ecology. Cambridge University, Cambridge, England. [ Links ]
Begon, M., J.L. Harper & C.R. Towsend. 1989. Ecología: individuos, poblaciones y comunidades. Omega, Barcelona, Spain. [ Links ]
Belsky, J.M. & S.F. Siebert. 1998. Non-timber forest products in community development and conservation: Desmoncus sp. (tie-tie) in Gales Point, Manatee, Belize, p. 224-244. In R.B. Primack, D. Bray, H. Galleti & I. Ponciano (eds.). Timber, Tourist and Temples: Conservation and Development in the Maya Forest of Belize, Guatemala, and Mexico. Island, Washington DC, USA. [ Links ]
Berlyn, G.P. & J.P. Miksche. 1976. Botanical microtechnique and cytochemistry. The Iowa State University, Ames, Iowa, USA. [ Links ]
Carson, F.L. 1997. Histotechnology, a self-instructional text. American Society of Clinical Pathologist, Chicago. USA. [ Links ]
Chinchilla A., M.R. 1993. Diagnóstico de la producción y comercialización de artículos elaborados con fibra de bayal (Desmoncus sp.) en los municipios de Santa Ana, San Francisco, San José, San Andrés y Flores, del Departamento del Petén. Facultad de Agronomía, Universidad de San Carlos de Guatemala, Guatemala. [ Links ]
Chinchilla A., M.R. 1994. Caracterización de las poblaciones de bayal (Desmoncus sp.), con fines de aprovechamiento artesanal, en la unidad de manejo forestal de San Miguel, San Andrés, Petén. Tesis para obtener el título de Ingeniero Agrónomo, Facultad de Agronomía,Universidad de San Carlos de Guatemala, Guatemala. [ Links ]
Escalante, S., C. Montaña & R. Orellana. 2004. Demography and potential extractive use of the liana palm Desmoncus orthacanthos Martius (Arecaceae), in southern Quintana Roo, Mexico. Forest Ecol. Manag. 187: 13-18 [ Links ]
Escalante, J. 1996. Caracterización del ratán y de tres especies posibles sustitutas. Tesis de Licenciatura, Instituto Tecnológico de Mérida, Mérida, Mexico. [ Links ]
Ewers, F.W. & J.B. Fisher. 1991. Why vines have narrow stems: histological trends in Bauhinia (Fabaceae). Oecología 88: 233-237. [ Links ]
Ewers, F.W., J.B. Fisher & S.T. Chiu. 1989. Water transport in the liana Bauhinia fassoglensis (Fabaceae). Plant Physiol. 91: 1625-1631. [ Links ]
Gaviño, G., J.C. Juárez & H.H. Figueroa. 1979. Técnicas biológicas selectas de laboratorio y de campo, Limusa, Mexico DF, Mexico. [ Links ]
Hallé, F., R.A.A. Oldeman & P.B. Tomlinson. 1978. Tropical trees and forests. Spriger, New York, USA. [ Links ]
Henderson, A. & F. Chávez. 1993. Desmoncus as useful palm in the western amazon basin. Principes 37: 184-186. [ Links ]
Henderson, A., G. Galeano & R. Bernal. 1995. Field guide to the Palms of the Americas. Princeton University, Princeton, New Jersey, USA. [ Links ]
Hutchison, B.A. & D.R. Matt. 1977. The distribution of solar radiation within a deciduous forest. Ecol. Monogr. 47: 185-207. [ Links ]
Isnard, S., T. Speck & N.P. Rowe. 2005. Biomechanics and development of the climbing habit in two species of the South American palm genus Desmoncus (Arecaceae). Am. J. Bot. 92: 11-23. [ Links ]
Jaffe, J.J., A.C. Leopold & R.C. Staples. 2002. Thigmo responses in plants and fungi. Am. J. Bot. 89: 375–382. [ Links ]
Johansen, D.A. 1974. Plant microtechnique. McGraw Hill, New York, USA. [ Links ]
Klotz, L.H. 1978. Observations on diameters of vessel in stems of palms. Principes 22: 99-106. [ Links ]
Liese, W. & G. Weiner. 1989. Anatomical structures for the identification of rattan. In A.N. Rao, I. Vongkaluang, J. Dransfield, N. Manokaran, C.B. Sastry & G. Dhanaranjan (eds.). Recent Research on Rattan. Faculty of Forestry, Kasetsart University, Thailand and International Development Research Centre, Canada. [ Links ]
Marmillod, D. & J. Galvez. 1998. Efecto del aprovechamiento maderero sobre la población de bayal, especie no maderable del bosque petenero. In BOLFOR, CIFOR, IUFRO. Memorias del Simposio Internacional sobre posibilidades de manejo forestal sostenible en América Tropical, julio 1997, Santa Cruz, Bolivia. [ Links ]
Niklas, K. 1992. Plant biomechanics: an Engineering approach to plant form and function. The University Chicago, Chicago. USA. [ Links ]
Olmsted, I.C., R. Durán, J.A. González-Iturbe, J. Granados & F. Tun. 1999. Vegetación. In Atlas de Procesos Territoriales de Yucatán. UADY, Facultad de Arquitectura, Mérida, Mexico. [ Links ]
Orellana, R. 1992. Síndromes morfológicos y funcionales de las palmeras de la Península de Yucatán. Bull. Inst. Fr. Etudes Andines 21: 651-667. [ Links ]
Orellana, R., P. Herrera, S. Rebollar, J. Escalante, G. López, S. Escalante & L. Gus. 1999. Studies on the potential uses of some native palms of the Yucatan peninsula (Mexico) as substitutes of rattan. Proc. of the 2nd Int. Symp. on ornamental palms and other monocots from the tropics. Acta Hort 486, s.p. [ Links ]
Phillips, M. 1991. The etnobotany and economic botany of tropical vines. In F.E. Putz & H.A. Mooney (eds.). The biology of vines. Cambridge, Cambridge, Massachussetts, USA. [ Links ]
Putz, F.E. & H.A. Mooney. 1991. The biology of vines. Cambridge, Cambridge, Massachussetts, USA. [ Links ]
Quero, H.J. 1992. Current status of Mexican palms. Principes 36: 203-216. [ Links ]
Rich, P.M. 1986. Mechanical architecture of arborescent rain forest palm. Principes 30:117-131. [ Links ]
Rich, P.M. 1987. Developmental anatomy of the stem of Welfa georgii, Iriartea gigantea, and other arborecent palms: implications for mechanical support. Am. J. Bot. 74: 792-802. [ Links ]
Rowe, N., S. Isnard & T. Speck. 2004. Diversity of Mechanical Architectures in Climbing Plants: An Evolutionary Perspective. J. Plant Growth Regul. 23: 108-128. [ Links ]
Ruzin, S. 1999. Plant microtechnique and microscopy. Oxford University, New York, USA. [ Links ]
Salisbury, F.B. & C.W. Ross. 1992. Plant Physiology. Wadsworth, Belmont, California, USA. [ Links ]
SAS Institute. 1998. SAS language guide for personal computers. SAS Institute, Cary, North Carolina, USA. [ Links ]
Schultes, R. 1940. Plantae Mexicana V. Desmoncus chinantlensis and its utilization in native basketry. Bot. Mus. Leafl 8: 134-140. [ Links ]
Siebert, S.F. 2000. Abundance and growth of Desmoncus orthacanthos Mart (Palmeae) in response to light and ramet harvesting in five forest sites in Belize. Forest Ecol. Manag.137: 83-90. [ Links ]
Silitonga, T. 1989. The effect of several cooking oil compositions on manau (Calamus manan Miq.) canes. In Recent research on rattans, p. 187-181. Proceedings of the International Rattan Seminar, Chiang Mai, Thailand. [ Links ]
Sokal, R.R. & F.J. Rohlf. 1980. Introducción a la bioestadística. Reverté, Barcelona, Spain. [ Links ]
Tilman, D. 1988. Plant strategies and the Dynamics and structure of plant communities. Princenton, New Jersey, USA. [ Links ]
Tomlinson, P.B. 1990. The structural biology of palms. Clarendon, Oxford University, New York, USA. [ Links ]
Tomlinson, P.B. & M.H. Zimmermann. 2003. Stem vascular architecture in the American climbing palm Desmoncus (Arecaceae-Arecoideae-Bactridinae). Bot. J. Linn. Soc. 142: 243-254. [ Links ]
Tyree, M.T. & F.W. Ewers. 1996. Hydraulic architecture of woody tropical plants, p. 217-243. In S.S. Mulkey, R.L. Chazdon & A.P. Smith (eds.). Tropical forest plant ecophysiology. Chapman, New York, USA. [ Links ]
Uhl, N.W. & J. Dransfield. 1987. Genera Palmarum. A classification of palms based on the work of Harold E. Moore, Jr. Allen, Lawrence, Kansas, USA. [ Links ]
Weiner, G. & W. Liese. 1990. Rattans-stem anatomy and taxonomic implications. IAWA J. 11: 61-70. [ Links ]
Weiner, G. & W. Liese. 1993. Generic identification key to rattan palms based on stem anatomical characters. IAWA J. 14: 55-61. [ Links ]
Zimmermann, U., F. Meinzer & F.W. Bentrup. 1995. How does water ascend in tall trees and other vascular plants. Ann. Bot. 76: 545-551. [ Links ]

