










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.56 n.1 San José Mar. 2008
Potential role of frugivorous birds (Passeriformes) on seed dispersal of six plant species in a restinga habitat, southeastern Brazil
Verônica Souza da Mota Gomes1,3*, Maria Célia Rodrigues Correia2, Heloisa Alves de Lima2 & Maria Alice S. Alves3*
1 Programa de Pós-Graduação em Ecologia, Universidade Federal do Rio de Janeiro, CCS, IB, Sala A1 008, CP 68020, 21941-590, Rio de Janeiro, RJ, Brazil.
2 Laboratório de Biologia da Reprodução, Museu Nacional do Rio de Janeiro, UFRJ, Quinta da Boa Vista s/n0, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brazil; halcar@acd.ufrj.br; mcorreia@acd.ufrj.br
3 Departamento de Ecologia, IBRAG, Universidade do Estado do Rio de Janeiro, Rua São Francisco Xavier, 524. Maracanã, 20550-011, Rio de Janeiro, RJ, Brazil. fax: 55 21 2587 7614; masa@uerj.br; smgomes@yahoo.com.br
* Corresponding authors
Abstract: Restingas are considered stressful habitats associated with the Brazilian Atlantic forest, and their ecological interactions are poorly known. The goal of the present study was to determine the potential role of frugivorous birds as seed dispersers in a restinga habitat. Data were collected in Parque Nacional da Restinga de Jurubatiba, southeastern Brazil, where the main physiognomy (Open Clusia Formation) is characterized by the presence of patches of vegetation covering 20 to 48 % of the sandy soil and reaching a height of 5 m. Birds were captured with mist nets (12 x 2.5 m; 36 mm mesh; 1 680 net-hrs) and had their fecal and regurgitate samples inspected for seeds. Six plant species found in these bird samples were studied. The germination of seeds obtained from plants was compared to those from the birds. Both groups of seeds were set on Petri dishes at room temperature and washed when infected with fungi. In general, there was no effect on germination rate, and the effect on germination speed was negative. Germination of seeds from Pilosocereus arrabidae treated by the birds seemed to be influenced by storage of defecated seeds, while few Miconia cinnamomifolia seeds both from plants and from birds germinated. Ocotea notata presented a great variation in time to the onset of germination, perhaps an advantage against dissecation. Aechmea nudicaulis, Clusia hilariana and Erythroxylum subsessile probably take advantage of the arrival to favorable microhabitats, not by the gut effect on the seeds. All plant species studied are numerically important for the community and some of them are main actors in the succession of vegetation patches. Among the birds, Mimus gilvus is an important resident species, endemic to restingas in Brazil, while Turdus amaurochalinus is a visitor and may be important for plants that fructify during its passage by the study site. Although the effect of pulp removal was only tested for one species (Achmea nudicaulis) in the present study, we confirmed that the tested effect of restinga frugivorous birds on seed germination was generally null. Although there is a need for more detailed studies on specific animal-plant interactions on this habitat, the overall effect of the birds on seed dispersal in restinga is probably positive. Rev. Biol. Trop. 56 (1): 205-216. Epub 2008 March 31.
Key words: Atlantic forest, Brazil, dispersal, Mimus gilvus, restinga, seed germination, Turdus amaurochalinus.
Restingas are coastal Brazilian ecosystems, geologically recent and poor in endemisms, being most of the flora and fauna typical of the Atlantic forest (Rizzini 1979, Lacerda et al. 1993, Cerqueira 2000). Plants exhibit many different physiological adaptations (Mattos et al. 2004) to the harsh conditions of the environment in restinga (low nutrient levels of the soil, ephemeral water availability, high temperatures, soil and air salinity) (Araujo et al. 1998). Then, asexual reproduction is probably important, as it occurs in other extreme habitats (Richards 1986). Indeed, many plant species present clonal development, although sexual reproduction is also present (Ormond et al. 1991). Therefore, studies concerning the investments and predominance of each reproductive strategy are lacking (Scarano et al. 2004).
The success of sexual reproduction depends on many factors acting in the production of flowers, presence and effectiveness of pollinators and fruit production. Furthermore, seed dispersal may be essential to final production of seedlings and their survival, as the disperser may alter seeds percentage of germination and germination rate (Traveset 1998), and also deposit them in specific favorable microhabitats (Calviño-Cancela 2004). However, there are few works that investigate the effect of frugivorous animals on seed dispersal in restingas (Castiglioni et al. 1995, Passos and Oliveira 2003).
Birds are potential seed dispersers in the restingas, as 67.5 % of woody plant species of the predominant physiognomy is zoochorous (Pimentel 2002) and 15 % to 25 % of bird species in the same environment may be considered frugivorous (Sick 1997, Reis and Gonzaga 2000, Alves et al. 2004). In general, the effect of the birds digestive system on the passage of the ingested seeds is positive or null, both on the percentage of germination and germination rate (Traveset 1998). However, the effects may vary with bird species (Traveset et al. 2001), plant species or habitat (Baskin and Baskin 1998), and also differences on population level (Traveset 1998).
Therefore, the goal of the present study was to evaluate the viability and speed of germination of seeds consumed by frugivorous birds in a restinga habitat, investigating the potential role of birds as seed dispersers in this environment. The specific questions were: 1) Are the eliminated seeds viable? 2) Do birds have any effect on seeds percentage of germination? 3) Do birds interfere in speed of germination?
From the plant species under investigation in the present study, those belonging to the families Clusiaceae, Erythroxylaceae, Lauraceae, Melastomataceae and Bromeliaceae are likely to present fully grown embryos and permeable seed cover structures in ripe fruits, as generally occurs for those families. Consequently, they probably present a lack of organic dormancy, being physical or chemical, which would need to be broken by the passage through the digestive system of some animal. Only Cactaceae tend to present physiological dormancy (Baskin and Baskin 1998). Therefore, in the present study, it was essentially tested the possibility of negative effects of the birds on the germination of ingested seeds.
Material and methods
Birds were captured and seeds collected in the Parque Nacional da Restinga de Jurubatiba (22º17S, 41º41W), a park created in 1998 in Macaé, on the north shore of Rio de Janeiro State. This region is a holocenic sandy plain covered by a mosaic of plant communities called restinga. The landscape is also marked by the presence of many lakes with various salinity levels. Climate is characterized by a wet season between October and April and a drier season between May and September. Mean annual pluviosity is 1 200 mm and temperature 22.6 ºC. The dominant plant formation is the Open Clusia Formation, formed by patches of vegetation that cover 20 % to 48 % of the soil and reach 5 m high, with few small plants in between. The park also includes forest formations, Ericaceae Shrub Formation and other formations with smaller plants, totaling ten plant communities (Henriques et al. 1986, Araujo et al. 1998, Pimentel 2002). The present study was developed in the Open Clusia Formation in two sites: next to "Lagoa Comprida" (22º1641S, 41º3941W) and approximately 2 km to Northeast from that (22º1613.2S, 41º3850.3W).
Between August 2002 and August 2004, field trips were carried out bimonthly in the park to capture birds with mist nets (12 x 2.5 m; 36 mm mesh; 1 680 net-hours). Birds were identified according to Dunning (1989), except for Elaenia spp., from which specimens were collected and compared to the collections of Museu de Zoologia da Universidade de São Paulo (MUZUSP) and Museu Nacional do Rio de Janeiro (MNRJ). The birds were released after sampled, except for the specimens collected for confirming identification. The frugivorous birds studied here were: Coryphospingus pileatus (Wied) 1821 (Pileated Finch), Cyanerpes cyaneus (Linnaeus) 1766 (Red-legged Honeycreeper), Elaenia chiriquensis Lawrence 1865 (Lesser Elaenia), Elaenia flavogaster (Thunberg) 1822 (Yellow-bellied Elaenia), Euphonia chlorotica (Linnaeus) 1766 (Purple-throated Euphonia), Mimus gilvus (Vieillot) 1808 (Tropical Mockingbird), Tachyphonus coronatus (Vieillot) 1822 (Rubycrowned Tanager), Thraupis sayaca (Linnaeus) 1766 (Sayaca Tanager), Tyrannus melancholicus Vieillot 1819 (Tropical Kingbird), Turdus albicollis Vieillot 1818 (White-necked Thrush), Turdus amaurochalinus Cabanis 1851 (Creamybellied Thrush), Zonotrichia capensis (Müller) 1776 (Rufous-collared Sparrow) (common names as in Dunning 1989 and authors and years from Sibley and Monroe Jr. 1990).
During the field trips, seeds apparently intact were eliminated by frugivorous birds, through defecation and natural regurgitation. Most of the times, it was not possible to know if seeds were defecated or regurgitated, as netted birds were kept in dark cloth bags while they waited to be manipulated. From the plant species consumed, it was chosen for the present study the most frequently recorded in the birds samples. The identification of the seeds was possible by comparison with a reference collection previously established. Then, seeds were also collected from fruits obtained directly from the plants, from three individuals of each studied plant species. The seeds were taken from the fruits, washed and stored in paper envelops. Animals and seeds collected are deposited at Depto de Ecologia/UERJ, under the custody of M. A. S. Alves.
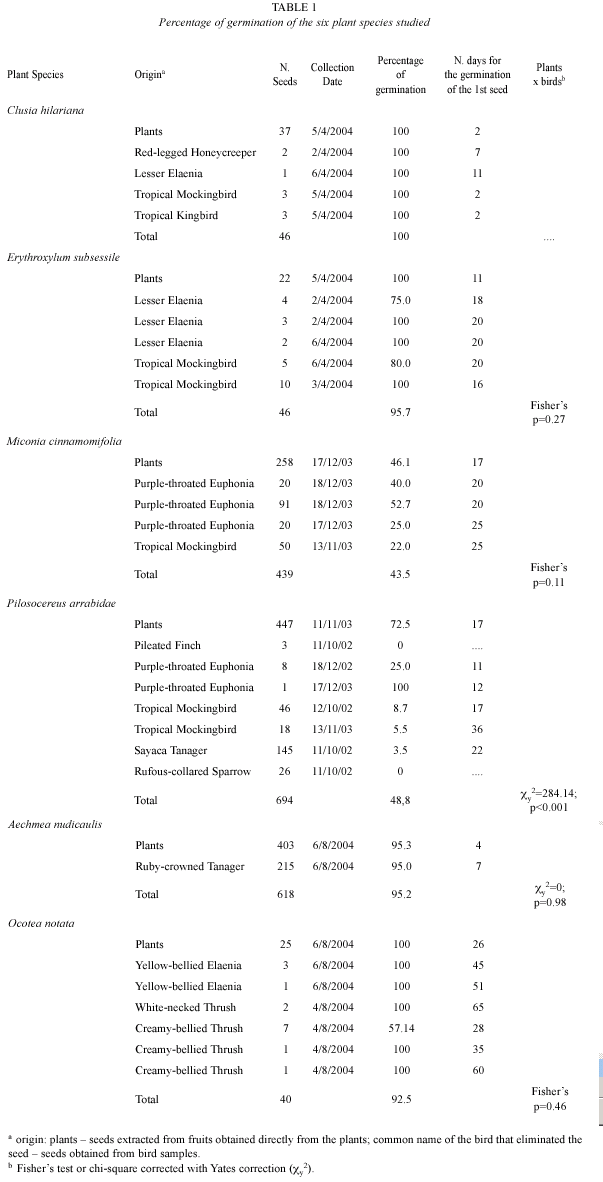
Collected seeds were set on Petri dishes with humid filter paper and left to room temperature and humidity. Seeds collected directly from the plants were set on one dish per species, while seeds from bird samples were grouped by origin (individual bird -see Table 1 to sample numbers). Dishes were examined regularly and the number of germinated seeds recorded. We considered germination when the radicle emerged. Experiments with Clusia hilariana Schltdl. (Clusiaceae), Erythroxylum subsessile (Mart.) O. E. Schulz (Erythroxylaceae), Miconia cinnamomifolia (DC.) Naudin (Melastomataceae) and Pilosocereus arrabidae (Lem.) Byles and G. D. Rowley (Cactaceae) have been conducted between April 13th and August 3rd 2004, while those with Aechmea nudicaulis (L.) Griseb. (Bromeliaceae) and Ocotea notata (Nees) Mez (Lauraceae) between August 9th and December 13th 2004. Seeds presenting fungi were washed with current water and reset in their dishes. That procedure was made because experiment conditions were different from what seeds face in the environment, so it is not possible to know if they would be infested in natural conditions. A possible effect of the presence of the pulp or the whole fruit was tested only for A. nudicaulis in September 2005: in one treatment, five whole fruits were set in Petri dishes on humid absorbent paper; in another treatment, 82 seeds were removed from the fruits along with the pulp and set to germinate in the same conditions.
To statistical analysis, seeds were treated as two groups (eliminated by the birds vs. collected from the plants) and the differences in percentage of seeds germinated and not germinated were tested by chi-square corrected with the Yates correction or by the Fishers test, when frequencies were small (Zar 1996; 2 x 2 contingency tables). Differences in speed of germination between seeds obtained from the plants and from the birds were tested by Linear Regression Analysis (as in Ellison et al. 1993): if the slope of the regression line (forced to the origin) was significantly greater than one, we concluded that birds enhanced the speed and if lower than one, that birds delayed germination.
Results
From the six plant species studied, only the seeds from Ocotea notata and Pilosocereus arrabidae germinated less when treated by the birds (Table 1). Comparing samples from P. arrabidae stored for five months with those stored for more than one year, there was no difference in the percentage of germination, both for those treated by Purple-throated Euphonia (X2=2.25, df=1, 0.25>p>0.10) and by Tropical Mockingbird (X2=0.196, df=1, 0.75>p>0.50). Particularly those seeds treated by Purplethroated Euphonia began to germinate earlier than the seeds taken directly from the plants. On the other hand, seeds treated by Pileated Finch and Rufous-collared Sparrow did not germinate, although the experiment was made only with intact seeds found, among intact and mashed seeds present in the samples of those two bird species. To O. notata, there was a marked variation in the time to the beginning of the germination.
Concerning speed of germination, there was a great variation among the plant species on the values of the slopes of the regression lines (Fig. 1), although none had slope equal or greater than one. This indicates that birds delayed seed germination.
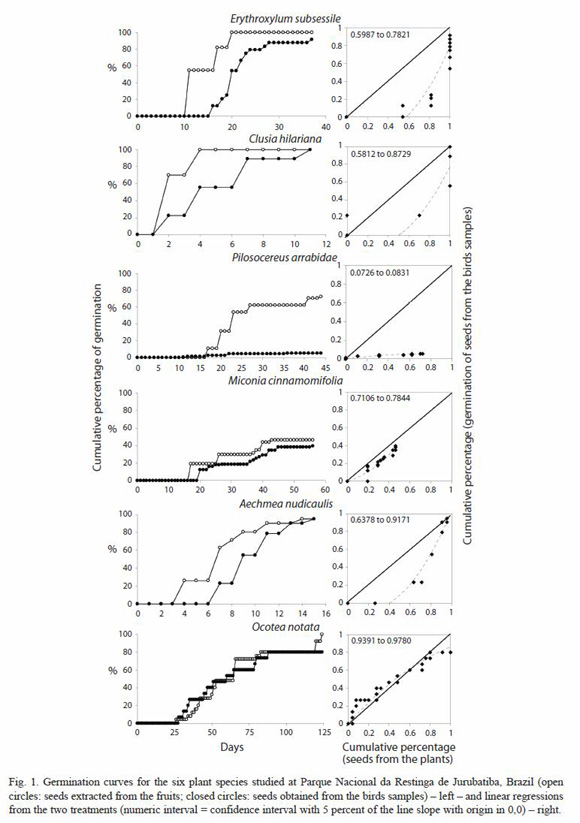
Fruits from A. nudicaulis tested were infested with fungi seven days after the beginning of the experiment and even being washed twice, they have rotten completely in 20 days. Seeds with pulp started to be infested also in seven days, but after washed, they started to germinate in more seven days (14 days after the beginning of the experiment). At the end of the experiment, 84 % of the seeds that had pulp and were washed germinated. When the seeds were washed, practically all the pulp was removed, as it is a liquid substance. Those results demonstrate that the presence of the pulp may delay the germination, even when compared to the seeds treated by the birds, which started to germinate in seven days of experiment (ten days after being collected, Fig. 1).
Discussion
Comparing bird effect on the percentage and speed of germination, the null effect is more frequent in germination studies, followed by the positive effects (Traveset 1998), what is in accordance with the present study only concerning percentage of germination and not speed of germination, which was reduced by the birds.
In the present study, from the seeds of P. arrabidae treated by the birds, only 5.6 % germinated (varying from zero to 100 % between bird species, see Table 1). Probably the germination of seeds that had contact with birds feces was influenced by storage, as the same plant species may reach 54.7 % of germination when treated by Ramphocelus bresilius (Linnaeus) 1766 (Brazilian Tanager) and set immediately to germinate (Castiglioni et al. 1995). Those authors have also observed that seeds from that species maintain their viability for many months when kept dry. Furthermore, Meyer and Witmer (1998) have found a negative effect of the presence of bird fecal material on seed germination for two plant species, one of Lauraceae and one of Caprifoliaceae families. Rufous-collared Sparrow and Pileated Finch revealed to be seed predators for P. arrabidae, what was in accordance with Silva (1988), who observed seed predation of Cereus peruvianus (another Cactaceae) by Rufouscollared Sparrow and Coryphospingus cucullatus (Müller) 1776 (Red-crested Finch). In fact, those two bird genus belong to a subfamily (Emberizinae) whose species are primarily seed predators (Sick 1997). In the Venezuelan semi-arid, seeds of two columnar cacti species, whose fruit pulp has germination inhibitors, germinated better when manually washed than when treated by birds or bats. Among seed dispersers, bats were responsible for the greatest enhance in percentage and delay in the beginning of germination and from the birds tested, Tropical Mockingbird was the best disperser (Naranjo et al. 2003). Many cacti species depend on nurse plants to the initial survivorship of seedlings and many of those nurse plants are perennial shrubs, which are in some cases replaced by the columnar cactus (Rojas-Aréchiga and Vázquez-Yanes 2000). In Restinga de Jurubatiba, P. arrabidae is positively associated to Allagoptera arenaria (Arecaceae), which probably facilitates its germination and is probably replaced by other species during a patch succession (Zaluar and Scarano 2000). Pilosocereus arrabidae has an additional role in this process by attracting seed dispersers that once arrive in the patch to consume its fruits, potentially bring seeds from other plant species (Zaluar 1997).
Miconia cinnamomifolia had seeds treated and non-treated by the birds germinating in the same proportions. Some Melastomataceae present specific light requirements to germinate (Miconia argentea, Pearson et al. 2003), while others present low germination percentage (Clidemia densiflora, Ellison et al. 1993), what suggests that the low percentage presented by M. cinnamomifolia may have been caused by its seeds specific requirements and not by storage. Manhães et al. (2003) also did not find any increase in percentage of germination or speed due to treatment by birds in M. urophylla, in an Atlantic Forest site.
The obtained effect of bird treatment on seed germination of O. notata was negative. However, pulp removal in Ocotea spp. by ants (Leal and Oliveira 1998) or birds (Wenny 2000) may increase percentage of germination, what is also true for other species of Lauraceae (Moraes and Paoli 1995). Such effect was not tested here for that plant species, what indicates that the overall effect of birds on that species seed germination is not totally solved. Furthermore, although O. notata had its percentage of germination reduced by the birds, it presented, among birds samples, a great variation in the time to initiate germination (28 to 65 days, see Table 1). This great variation may mean different dormancy patterns, what occurs even for seeds produced by the same individual plant (Scarano 1998). Under an evolutive perspective, the greater the diversity of seed dispersers, the greater the diversity of gut treatments over the seeds and consequently the greater the variation of times for seedling emergence, diluting dissecation risk in a mesic habitat with an unpredictable rainfall pattern (Izhaki and Safriel 1990). In Restinga de Jurubatiba, during the fructification period of O. notata (July to October, Gomes 2006), rainfall is lower or less regular than the rest of the year (Henriques et al. 1986) and an advantage related to this variation in germination may coincide with what was found by Izhaki and Safriel (1990).
Concerning the remaining species studied, neither A. nudicaulis nor C. hilariana germinate on bare sand (Scarano 2002) and a congeneric species of E. subsessile (E. ovalifolium) germinates very well in the interior of bromeliads tanks (Fialho and Furtado 1993), what also occurs with C. hilariana (Scarano 2002). Aechmea nudicaulis also presents limitations in germinating under high temperatures or in absence of light (Pinheiro and Borghetti 2003). Considering that birds frequently defecate and regurgitate seeds while perched on some vegetation patch (V. S. M. Gomes pers. obs.), they increase the chances of seeds find a favorable microhabitat.
Some studies have demonstrated that pulp removal by birds may have positive effects on seed germination of some plant species (Barnea et al. 1991, Meyer and Witmer 1998, Yagihashi et al. 1999), including restinga species (Castiglioni et al. 1995). In the study of Castiglioni et al. (1995) on the effect of Brazilian Tanager on the germination of restinga species, the effect of pulp removal on percentage of germination was null for seven species and positive for four, and the latter species had also the speed of germination increased. Although the effect of pulp removal was only tested for one species in the present study, we confirmed that the tested effect of restinga frugivorous birds on seed germination was generally null. At the end, the overall effect of the birds is probably positive, counting for pulp removal and microhabitat arrival. Calviño-Cancela (2004) have found that some frugivores were essential to seed germination of Corema album (Empetraceae) in the Mediterranean, not by the direct effects on the seeds, but for their ability of carrying them to favorable microhabitats. Even the plant species that lost some seeds predated by the birds had seeds germinating after gut treatment, another indication that some seeds may survive and some species benefit from that relationship, which conceivably results in a positive balance, or a mutualistic relationship.
The bird species studied are considered generalists in habitat and diet, except for Tropical Mockingbird, species restricted to restingas in Brazil (Sick 1997, Reis and Gonzaga 2000). Such species deserves more detailed studies on their role as a resident seed disperser. On the other hand, Creamy-bellied Thrush is a visitant frugivorous species whose role as seed disperser should also be better investigated, as some plant species may depend on seed dispersal by migratory birds (Poulin et al. 1999). Migratory thrushes (Turdus species) consume fruits and disperse seeds of many plants in arid shrublands in the Mediterranean (Barnea et al. 1992, Obeso and Fernández-Calvo 2002), as well as in urban habitats in the tropics (Mallet-Rodrigues 2001). The effects of species of this genus on seeds of many plant species may be null or positive (Barnea et al. 1991, Traveset and Willson 1997) not counting for pulp removal. In Restinga de Jurubatiba, Creamy-bellied Thrush may be especially important for the dispersal of O. notata, whose fructification coincides with the presence of this bird in the restinga, making those two species mutually important, considering the high frequency of those seeds in fecal samples (Gomes 2006).
From the plant species studied, C. hilariana, E. subsessile and O. notata are among the six species with greatest importance value index (IVI) in the shrub stratum and A. nudicaulis and P. arrabidae are among the eight species with greatest IVI in the herbaceous stratum in the Open Clusia Formation of Restinga de Jurubatiba (Araujo et al. 2004). Particularly Clusia spp. and Aechmea spp. have a special role in the community by facilitating the germination and establishment of other species in restinga (Zaluar and Scarano 2000). Miconia cinnamomifolia is mainly abundant on the surroundings of lagoons (D.S.D. Araujo pers. obs.). Given the great importance of such plant species to community structure and as food sources for birds in restinga, we suggest that future studies should focus on relative investments on sexual and asexual reproduction. Restinga environments may be interesting scenarios for those studies because plants are not endemic and are subject to extreme environmental conditions, differently from their distribution centers (Scarano 2002). Speciesspecific studies should also be carried on, to determine the dispersers effectiveness (Schupp 1993), mainly because the effect of secondary dispersers (Passos and Oliveira 2002, 2004) may be great in restinga habitats and alter the seed rain patterns.
Acknowledgments
To the staff of Laboratório de Ecologia de Aves/Universidade do Estado do Rio de Janeiro (UERJ) (especially Alline Storni, Adriano Lagos, Vanessa Tomás), Charles Ozanick, Francisco Mallet-Rodrigues, Karina Amaral, Davor Vrcibradic, Carlos Humberto Oliveira, Mariana Janiszewski and Ricardo Freitas for helping in the field work. To Dorothy S. D. Araujo and Alexandre Quinet (Ocotea) for the identification of the plants. To Luiz Fábio Silveira and Érica Hasui for helping with birds identification at Museu de Zoologia da Universidade de São Paulo (MUZUSP) and Marcos Raposo for helping at Museu Nacional do Rio de Janeiro (MNRJ). To Paulo Teixeira de Souza for helping in the germination experiments. To F. R. Scarano and R. Monteiro for reviewing the manuscript. To Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Fundação de Apoio à Pesquisa do Estado do Rio de Janeiro (FAPERJ) for scholarships to V. S. M. Gomes. To Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), for the research grant awarded to M. A. S. Alves (process number 302718/03-6). To Idea Wild for the mist nets and to Instituto Biomas and UERJ for the vehicles used in the field. This work is a subproject of Grupo de Vertebrados/ Peld - site 5/CNPq. Birds were captured under the license number P029/03 (Cemave/IBAMA) and collection of specimens under the licenses number 106/2003 and 116/2004 (IBAMA/DIREC -Parque Nacional da Restinga de Jurubatiba), 056/2002 and 093/2004 (Fauna).
Resumen
Las restingas se consideran hábitats estresantes asociados al bosque del Atlántico brasileño y sus ecológicas se conocen poco. El objetivo principal del presente trabajo fue determinar el papel potencial de las aves frugívoras como dispersores de plantas en un hábitat de restinga. La información se recolectó en el Parque Nacional da Restinga de Jurubatiba, Brasil. Las aves se capturaron con redes de niebla y sus muestras fecales y de regurgitación se inspeccionaron en busca de semillas. Estudiamos seis especies de plantas de estas muestras. En general, el efecto del paso por el sistema digestivo de las aves sobre el porcentaje de germinación fue nulo (y negativo en la velocidad de germinación). Todas las especies de plantas estudiadas son numéricamente importantes a la comunidad y algunas de ellas tienen papeles principales en la sucesión de parches de vegetación. Entre las aves, Mimus gilvus es una especie residente importante, endémica de tales hábitats en Brasil, mientras que Turdus amaurochalinus es una especie visitante y puede ser importante para las plantas que fructifican durante su paso por el sitio del estudio. El efecto general de los pájaros en la dispersión de semillas en la restinga es probablemente positivo.
Palabras clave: Dispersión de semillas, bosque atlántico brasileño, germinación, Mimus gilvus, restinga, Turdus amaurochalinus.
Received 03-X-2006. Corrected 27-IV-2007. Accepted 27-VII-2007.
References
Alves, M.A.S., A. Storni, E.M. Almeida, V.S.M. Gomes, C.H.P. Oliveira, R.V. Marques & M.B. Vecchi. 2004. A comunidade de aves na Restinga de Jurubatiba, pp. 199-214. In C.F.D. Rocha, F.A. Esteves & F.R. Scarano (eds.). Pesquisas ecológicas de longa duração na Restinga de Jurubatiba: Ecologia, História Natural e Conservação. RiMa, São Carlos, São Paulo, Brazil. [ Links ]
Araujo, D.S.D., M.C. A. Pereira, & M.C.P. Pimentel. 2004. Flora e estrutura de comunidades na Restinga de Jurubatiba - Síntese dos conhecimentos com enfoque especial para a Formação Aberta de Clusia, pp. 59-76. In C.F.D. Rocha, F.A. Esteves & F.R. Scarano (eds.). Pesquisas ecológicas de longa duração na Restinga de Jurubatiba: Ecologia, História Natural e Conservação. RiMa, São Carlos, São Paulo, Brazil. [ Links ]
Araujo, D.S.D., F.R. Scarano, C.F.C. Sá, B.C. Kurtz, H.L.T. Zaluar, R.C.M. Montezuma & R.C. Oliveira. 1998. Comunidades Vegetais do Parque Nacional da Restinga de Jurubatiba, pp. 39-62. In F.A. Esteves (ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Município de Macaé. UFRJ, Rio de Janeiro, Rio de Janeiro, Brazil. [ Links ]
Barnea, A., Y. Yom-Tov & J. Friedman. 1991. Does ingestion by birds affect seed germination? Funct. Ecol. 5: 394-402. [ Links ]
Barnea, A., Y. Yom-Tov & J. Friedman. 1992. Effect of frugivorous birds on seed dispersal and germination of multi-seeded fruits. Acta Oecol. 13: 209-219. [ Links ]
Baskin, C.C. & J.M. Baskin. 1998. Seed: ecology, biogeography, and evolution of dormancy and germination. Academic, San Diego, California, USA. [ Links ]
Calviño-Cancela, M. 2004. Ingestion and dispersal: direct and indirect effects of frugivores on seed viability and germination of Corema album (Empetraceae). Acta Oecol. 26: 55-64. [ Links ]
Castiglioni, G.D. A., L.S.T. Cunha & L.P. Gonzaga. 1995. Ramphocelus bresilius como dispersor das sementes de plantas da restinga de Barra de Maricá, Estado do Rio de Janeiro (Passeriformes: Emberizidae). Ararajuba 3: 94-99. [ Links ]
Cerqueira, R. 2000. Biogeografia das restingas, pp. 65-75. In F.A. Esteves & L.D. Lacerda (eds.). Ecologia de restingas e lagoas costeiras. NUPEM\UFRJ, Rio de Janeiro, Brazil. [ Links ]
Dunning, J.S. 1989. South American Birds. A photographic aid to identification. Harrowood Books, Newton Square, Pennsylvania, USA. [ Links ]
Ellison, A.M., J.S. Denslow, B.A. Loiselle & M.D. Brenes. 1993. Seed and seedling ecology of neotropical Melastomataceae. Ecology 74: 1733-1749. [ Links ]
Fialho, R.F. & A.L.S. Furtado. 1993. Germination of Erythroxylum ovalifolium (Erythroxylaceae) seeds within the terrestrial bromeliad Neoregelia cruenta. Biotropica 25: 359-362. [ Links ]
Gomes, V.S.M. 2006. Variação espacial e dieta de aves terrestres na Restinga de Jurubatiba, RJ. PhD Thesis. Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil. [ Links ]
Henriques, R.P.B., D.S.D. Araujo & J.D. Hay. 1986. Descrição e classificação dos tipos de vegetação da restinga de Carapebus, Rio de Janeiro. Rev. bras. Bot. 9: 173-189. [ Links ]
Izhaki, I. & U.N. Safriel. 1990. The effects of some Mediterranean scrubland frugivores upon germination patterns. J. Ecol. 78: 56-65. [ Links ]
Lacerda, L.D., D.S.D. Araujo & N.C. Maciel. 1993. Chapter 32: Dry coastal ecosystems of the tropical Brazilian coast, pp. 477-493. In E. van der Maarel (ed.). Dry Coastal Ecosystems: Africa, America and Oceania. Elsevier, Amsterdam, Noord Holland, The Netherlands. [ Links ]
Leal, I.R. & P.S. Oliveira. 1998. Interactions between fungus-growing ants (Attini), fruits and seeds in Cerrado vegetation in southeast Brazil. Biotropica 30: 170-178. [ Links ]
Mallet-Rodrigues, F. 2001. Potencial de germinação de sementes de Ficus microcarpa tendo o sabiá-laranjeira, Turdus rufiventris, como dispersor. Tangara 1: 30-33. [ Links ]
Manhães, M.A., L.C.S. Assis & R.M. Castro. 2003. Frugivoria e dispersão de sementes de Miconia urophylla (Melastomataceae) por aves em um fragmento de Mata Atlântica secundária em Juiz de Fora, Minas Gerais, Brazil. Ararajuba 11: 173-180. [ Links ]
Mattos, E.A., M.I.G. Braz, P.O. Cavalin, B.H.P. Rosado, J.M. Gomes, L.S.T. Martins & R.C.O. Arruda. 2004. Variação espacial e temporal em parâmetros fisio-ecológicos de plantas, pp. 100-116. In C.F.D. Rocha, F.A. Esteves & F.R. Scarano (eds.). Pesquisas ecológicas de longa duração na Restinga de Jurubatiba: Ecologia, História Natural e Conservação. RiMa, São Carlos, São Paulo, Brazil. [ Links ]
Meyer, G.A. & M.C. Witmer. 1998. Influence of seed processing by frugivorous birds on germination success of three North American shrubs. Am. Midl. Nat. 140: 129-139. [ Links ]
Moraes, P.L.R. & A.A.S. Paoli. 1995. Dispersão e germinação de sementes de Cryptocarya moschata Nees and Martius Ex Nees, Ocotea catharinensis Mez e Endlicheria paniculata (Sprengel) Macbride (Lauraceae). Arq. Biol. Tecnol. 38: 1119-1129. [ Links ]
Naranjo, M.E., C. Rengifo & P.J. Soriano. 2003. Effect of ingestion by bats and birds on seed germination of Stenocereus griseus and Subpilocereus repandus (Cactaceae). J. Trop. Ecol. 19: 19-25. [ Links ]
Obeso, J.R. & J.C. Fernández-Calvo. 2002. Fruit removal, pyrene dispersal, post-dispersal predation and seedling establishment of a bird-dispersed tree. Plant Ecol. 165: 223-233. [ Links ]
Ormond, W.T., M.C.B. Pinheiro, H.A. Lima, M.C.R. Correia & A.C. Castro. 1991. Sexualidade das plantas da Restinga de Maricá, RJ. Bol. Mus. Nac. l 87: 1-24. [ Links ]
Passos, L. & P.S. Oliveira. 2002. Ants affect the distribution and performance of seedlings of Clusia criuva, a primarily bird-dispersed rain forest tree. J. Ecol. 90: 517-528. [ Links ]
Passos, L. & P.S. Oliveira. 2003. Interactions between ants, fruits and seeds in a restinga forest in south-eastern Brazil. J. Trop. Ecol. 19: 261-270. [ Links ]
Passos, L. & P.S. Oliveira. 2004. Interaction between ants and fruits of Guapira opposita (Nyctaginaceae) in a Brazilian sandy plain rain forest: ant effects on seeds and seedling. Oecologia 139: 376-382. [ Links ]
Pearson, T.R.H., D.F.R.P. Burslem, C.E. Mullins & J.W. Dalling. 2003. Functional significance of photoblastic germination in neotropical pioneer trees: a seeds eye view. Funct. Ecol. 17: 394-402. [ Links ]
Pimentel, M. 2002. Variação espacial na estrutura de comunidades vegetais da Formação Aberta de Clusia no Parque Nacional da Restinga de Jurubatiba, RJ. PhD Thesis. Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil. [ Links ]
Pinheiro, F. & F. Borghetti. 2003. Light and temperature requirements for germination of seeds of Aechmea nudicaulis (L.) Griesebach and Streptocalyx floribundus (Martius ex Schultes F.) Mez (Bromeliaceae). Acta Bot. Bras. 17: 27-35. [ Links ]
Poulin, B., S.J. Wright, G. Lefebvre & O. Calderon. 1999. Interespecific synchrony and asynchrony in the fruiting phenologies of congeneric bird-dispersed plants in Panama. J. Trop. Ecol. 15: 213-227. [ Links ]
Reis, H.B.R. & L.P. Gonzaga. 2000. Análise da distribuição geográfica das aves das restingas do Estado do Rio de Janeiro, pp. 165-178. In F.A. Esteves & L.D. Lacerda (eds.). Ecologia de restingas e lagoas costeiras. NUPEM\UFRJ, Rio de Janeiro, Brazil. [ Links ]
Richards, A.J. 1986. Plant breeding systems. George Allen and Unwin, London, United Kingdom. [ Links ]
Rizzini, C.T. 1979. Tratado de fitogeografia do Brazil. Aspectos sociológicos e florísticos. Vol. 2. Universidade de São Paulo, São Paulo, Brazil. [ Links ]
Rojas-Aréchiga, M. & C. Vázquez-Yanes. 2000. Cactus seed germination: a review. J. Arid Environ. 44: 85-104. [ Links ]
Scarano, F.R. 1998. A comparison of dispersal, germination and establishment of woody plants subjected to distinct flooding regimes in Brazilian flood-prone forests and estuarine vegetation. Oecologia Bras. 4: 177-193. [ Links ]
Scarano, F.R. 2002. Structure, function and floristic relationships of plant communities in stressful habitats marginal to the Brazilian Atlantic Forest. Ann. Bot. 90: 517-524. [ Links ]
Scarano, F.R., P. Cirne, M.T. Nascimento, M.C. Sampaio, D.M. Villela, T. Wendt & H.L.T. Zaluar. 2004. Ecologia vegetal: integrando ecossistema, comunidade, populações e organismos, pp. 77-97. In
C.F.D. Rocha, F.A. Esteves & F.R. Scarano (eds.). Pesquisas ecológicas de longa duração na Restinga de Jurubatiba: Ecologia, História Natural e Conservação. RiMa, São Carlos, Brazil.
Schupp, E.W. 1993. Quantity, quality and the effectiveness of seed dispersal by animals. Vegetatio 107/108: 15-29. [ Links ]
Sibley, C.G. & B.L. Monroe Jr. 1990. Distribution and taxonomy of birds of the world. Yale University, New Haven, Connecticut, USA. [ Links ]
Sick, H. 1997. Ornitologia Brasileira. Nova Fronteira, Rio de Janeiro, Brazil. [ Links ]
Silva, W.R. 1988. Ornitocoria em Cereus peruvianus (Cactaceae) na Serra do Japi, Estado de São Paulo. Rev. bras. Biol. 48: 381-389. [ Links ]
Traveset, A. 1998. Effect of seed passage through vertebrate frugivores guts on germination: a review. Persp. Plant Ecol. Evol. Syst. 12: 151-190. [ Links ]
Traveset, A., N. Riera & R.E. Mas. 2001. Passage through bird guts causes interspecific differences in seed germination characteristics. Funct. Ecol. 15: 669-675. [ Links ]
Traveset, A. & M.F. Willson. 1997. Effect of birds and bears on seed germination of fleshy-fruited plants in temperate rainforests of southeast Alaska. Oikos 80: 89-95. [ Links ]
Wenny, D.G. 2000. Seed dispersal, seed predation, and seedling recruitment of a neotropical montane tree. Ecol. Monogr. 70: 331-351. [ Links ]
Yagihashi, T., M. Hayashida & T. Miyamoto. 1999. Effects of bird ingestion on seed germination of two Prunus species with different fruit-ripening seasons. Ecol. Res. 14: 71-76. [ Links ]
Zaluar, H.L.T. 1997. Espécies focais e a formação de moitas na restinga aberta de Clusia, Carapebus, RJ. Master Thesis. Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil. [ Links ]
Zaluar, H.L.T. & F.R. Scarano. 2000. Facilitação em restingas de moitas: um século de buscas por espécies focais, pp. 3-23. In F. A. Esteves & L. D. Lacerda (eds.). Ecologia de restingas e lagoas costeiras. NUPEM\UFRJ, Rio de Janeiro, Brazil. [ Links ]
Zar, J.H. 1996. Biostatistical Analysis. Prentice-Hall, London, London, United Kingdom. [ Links ]

