










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.56 n.1 San José Mar. 2008
Growth of the Pacific jack Caranx caninus (Pisces: Carangidae) from the coast of Colima, México
Elaine Espino Barr1, Manuel Gallardo Cabello2, Esther Guadalupe Cabral Solís1, Arturo Garcia Boa1 & Marcos Puente Gómez1
1 CRIP-Manzanillo,INP, Playa Ventanas s/n, Manzanillo, Colima, 28200, México. Tel: 01 (314) 33 32 37 50. Fax: 01(314) 33 32 37 51; elespino@bay.net.mx; escama@webtelmex.net.mx; elespino@gmail.com
2 Instituto de Ciencias del Mar y Limnología, UNAM, Apartado Postal 70-305, C.P 04510, México, D.F., México; gallardo@mar.icmyl.unam.mx
Abstract: The Pacific jack Caranx caninus is a common species fished by artisanal fishermen off the coast of Colima, México. During 2002, monthly samples of morphometric data and otoliths were taken to determine age and growth. Seven age groups were identified. The highest growth, 14.4 cm, takes place during the first year of life. During the second year, C. caninus grows 11.76 cm; the third year 9.61 cm; the fourth 7.85 cm; the fifth 6.41 cm and sixth year 5.24 cm. The constants of von Bertalanffy´s growth equation were: L∞ = 83.26 cm, W∞ = 18.138 g, K = 0.202, to = -0.283 and A0.95 = 15 years. Growth curves of other species of the same genus were calculated in order to compare them with the one obtained in the present work. The gonadosomatic index presented higher values during November and May. The periods of more intensive feeding are from August to February. Rev. Biol. Trop. 56 (1): 171-179. Epub 2008 March 31.
Key words: Caranx caninus, age and growth, length frequency distribution, sagittae, asterisci, von Bertalanffys equation, longevity.
Caranx caninus Günther 1868 (Fig. 1) is a pelagic and coastal species of the continental shelf from California, USA to the north of Perú (Chirichigno et al. 1982). It is fished during the whole year in the coast of Colima, México, and its capture volumes summing about 15 % of total catch annual average make it important (Cruz-Romero et al. 1993).

The Pacific jack is considered a second and third class commercial product; its price does not exceed five pesos on the beach or eight pesos per kg in the market ($0.50 to $0.80 USD). In the State of Guerrero it is considered a second class fish but with a better price (Pérez-Velásquez, pers. com.). Analysis on its taxonomic description have been made by Castro-Aguirre (1978), Chirichigno et al. (1982), van der Heiden (1985), Bussing and López (2004); on general aspects of its fishery by Ramírez-Granados (1977), Cruz-Romero et al. (1989, 1996), Espino-Barr et al. (2003, 2004). Cruz-Romero et al. (1993) analyzed general aspects of its biology. Recently Torres-Aguilar (2002) studied its age and growth determination in Guerrero and Michoacán, with vertebrae and spines. Espino-Barr et al. (2006) determined the average length for each age with the analysis of the sagittae and the asterisci.
Age and growth analysis are necessary to understand the populations dynamics and the fluctuations of its biomass in time and space (Lucano-Ramírez et al. 2006). The use of the ELEFAN method (Gayanilo et al. 1994; Jiménez-Badillo 2006) and the analysis of the otoliths are good instruments to estimate age groups and obtain growth parameters.
The weight-length relation helps to calculate the condition factor that is necessary to understand weight variations through the year (Safran 1992). Likewise, the calculations of the gonadosomatic index and the gastric repletion index help to understand the growth phenomena, the gonadic maturity and the trophic dynamics of the populations (Saucedo-Lozano et al. 1999; Rojas-Herrera et al. 2004; Rojas et al. 2004; Aguirre-León and Díaz-Ruiz 2006).
In this study the average length for each age was estimated by means of the monthly length frequency distributions. The time of the slow and fast growth bands formation in the otoliths sagittae was determined. The growth constants of von Bertalanffy´s (1938) equation were calculated from the polimodal analysis and from the analysis of sagittae and asterisci. The longevity and the values of the condition factor, gonadosomatic and gastric repletion were also calculated.
Materials and methods
From January to December 2002, fifty specimens of the Pacific jack C. caninus were obtained monthly from the commercial catch in Manzanillo, Colima, México, and taken to the lab where standard (Ls, cm) and total length (Lt, cm), total (Wt, g) and eviscerated weight (We, g), gonad (Wg, g) and stomach weight (Ws, g) were recorded for each individual. The total weight (Wt), standard (Ls) and total length (Lt) of over 200 individuals per month were taken in situ.
The length frequency distribution was analyzed with the ELEFAN program of the FISAT package (Gayanilo et al. 1994) to obtain the average length of the polimodal curves corresponding to each age group.
The average length of the age groups determined by the analysis of the otoliths sagittae and asterisci by Espino-Barr et al. (2006) were used to obtain the constants of von Bertalanffy´s (1938) growth equation. The observed values were: for age 0=6.33 cm, age 1=15.09 cm; age 2=32.19 cm; age 3=41.50 cm; age 4=48.17 cm; age 5=55.58 cm and age 6=58.75 cm. The time of the growth ring formation was determined, observing whether the borders had slow or fast growth rings. In every case otoliths were observed by transparency with transmitted light; the hyaline zone corresponds to the slow growth band and the opaque zone to the fast growth band, which is in contrast with reflected light (Blacker 1974).
There were no growth differences between sexes (Espino-Barr et al. 2006).
The constants L∞, K y to of von Bertalanffy´s (1938) equation were obtained with Ford´s (1933), Walford´s (1946) and Gulland´s method (1964) and were adjusted by convergent iterations with Newton´s algorithm with the Solver program in Excel software (Microsoft 1992).
Weight-length relationship was calculated with the function W=a*Lb, and a "t student test" was used to prove allometry (Mendenhall 1987, Zar 1996). The weight for every age was obtained with the growth data in length and the weight-length function. Weight growth was obtained substituting Lt and L∞ for Wt and W∞, in von Bertalanffy´s (1938) equation. Age limit or longevity (95 % of L∞) was determined with Taylor´s equation (1958, 1960):
A0.95=ln(1-0.95)/k+to
Monthly values of the condition factor, equivalent to the "a" parameter of weightlength equation, were obtained for total and eviscerated weight (Safran 1992), and compared with the confidence interval, to explain changes in the gonad, liver and stomach throughout the year.
The gonadosomatic index (GSI) was determined with the equation described by Rodríguez-Gutiérrez (1992): GSI=Wg/Wt*100, where Wg is gonad weight and Wt is the total weight of the individual.
The gastric repletion index (GRI) according to Sierra and Popota (1988) is GRI=Ws/Wt*100, where Ws is the stomach weight and Wt is the total weight of the individual.
Results
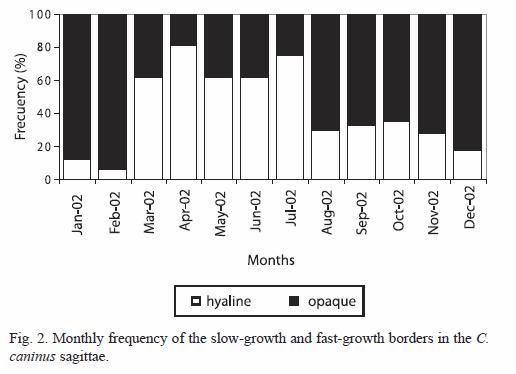
Growth rings time of formation of the slow and fast growth bands: C. caninus shows that a higher percentage of otoliths with fast growth borders occurs during August to February (autumn-winter), while the highest percentage of otoliths with slow growth bands in the border were observed during March to July (spring-summer) (Fig. 2).
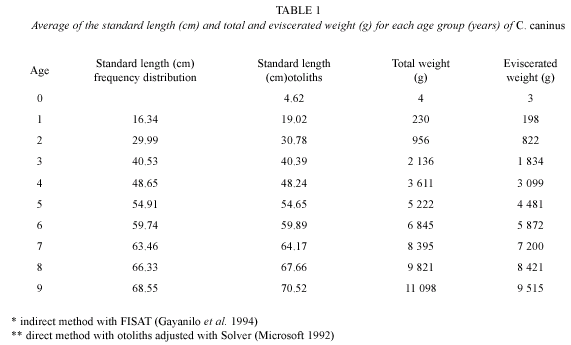
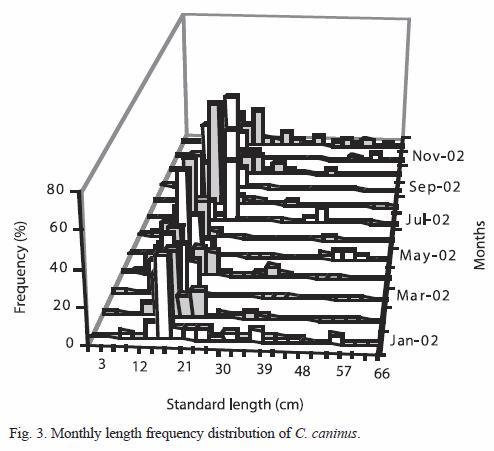
Analysis of length frequency: the values of standard length for each age are shown in Table 1 and Figure 3. The first year the organism grows 16.34 cm, during the second year it has reached 30 cm. Growth parameters for the standard length are: L∞=76.00 cm, K=0.26 years-1 and to=- 0.07. The highest increment occurs during the first year of life, during the next years growth rate decreases notably: 13.66 cm from age one to age two; 10.53 from age two to age three; 8.12 from age three to age four; 6.26 from age four to age five and 4.82 cm from age five to age six.
Analysis of otoliths: analysis of the sagittae and asterisci otoliths permitted to identify seven age groups. Growth parameters for the standard length were: L∞=83.26 cm; K=0.202 years-1; to=-0.283. Instantaneous growth rate was 14.4 cm from age cero to age one; 11.76 from age one to age two; 9.61 from ages two to three; 7.85 from ages three to four; 6.41 from ages four to five and 5.24 cm from ages five to six.
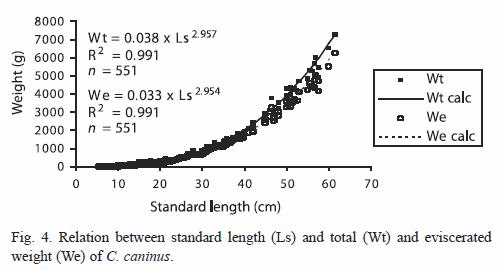
Growth in weight: The allometric growth index value of the weight-length equation was isometric: b=2.957 with total weight data (t=0.561<t0.05(2)426) and b=2.954 with eviscerated specimens (t=0.875<t0.05(2)91); they were statistically equal to 3 (Fig. 4).
Theoretical growth in weight: table 1 shows the values of theoretical growth for total and eviscerated weight at each age. The first year the organism reaches 230 g of total weight and 198 g of eviscerated weight. For the second year the values increase potentially: 956 g of total weight and 822 g of eviscerated weight. The asymptotic total weight was Wt∞ = 18.138 g and the eviscerated asymptotic weight, We = 15.544 g.
Longevity (Age A0.95): C. caninus reached 95 % of its infinite length L∞ at 15 years.
Bioecologic considerations during the growth of C. caninus: analysis of the confidence intervals of the indexes showed that there are no significant changes throughout the year. Figure 5 shows the trends of the monthly average data of the indexes of condition factor (a), gonadosomatic (GSI) and the gastric repletion (GRI). The massive spawning periods occur in November and May (GSI). The most active feeding season represented as condition factor (a tot and a evi) is from August to February. The gastric repletion index (GRI) seems stable during this period, reaching its higher value in December, January and May.
Discussion
According to the results showed in Table 2, each year a band of fast and of slow growth are deposited in the otoliths, validating the use of this structure to estimate age based in the number of bands in Caranx caninus.
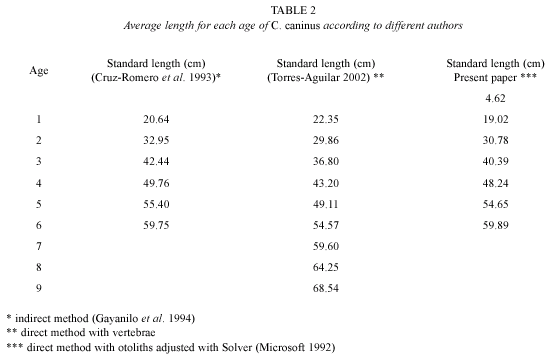
In this paper, the values obtained with the length frequency analysis were very similar to those found by the identification of growth rings in the sagittae and asterisci otoliths, which renders age determination in C. caninus valid with both methods. The otolith analysis allowed the calculation of the average length for age group cero, which was not identified by the polimodal curves, due to the very few organisms under one year of age in the commercial catch because of gear size selectivity. The use of the ELEFAN program (Gayanilo et al. 1994) distinguished similar age group comparable with those found by Cruz-Romero et al. (1993), who also used an indirect method for their analysis. It is important to note that the values of this paper and those of Torres-Aguilar (2002) in relation to the ones found a decade ago by Cruz-Romero et al. (1993) suggest that the population is stable in spite of the fluctuations in the physical, chemical and fishery variables; a reduction in its length which is an over fishing symptom has not been observed (Espino-Barr 2000).
Allometric growth index shows intrannual variations, which depend in great measure on the relation of the organism with its environment and is inversely proportional to the condition factor (Safran 1992). Cruz-Romero et al. (1993) found a negative allometric value, i.e., b=2.28, while in the present work the indexes are isometric b=2.957 with total weight and b=2.954 with eviscerated weight.
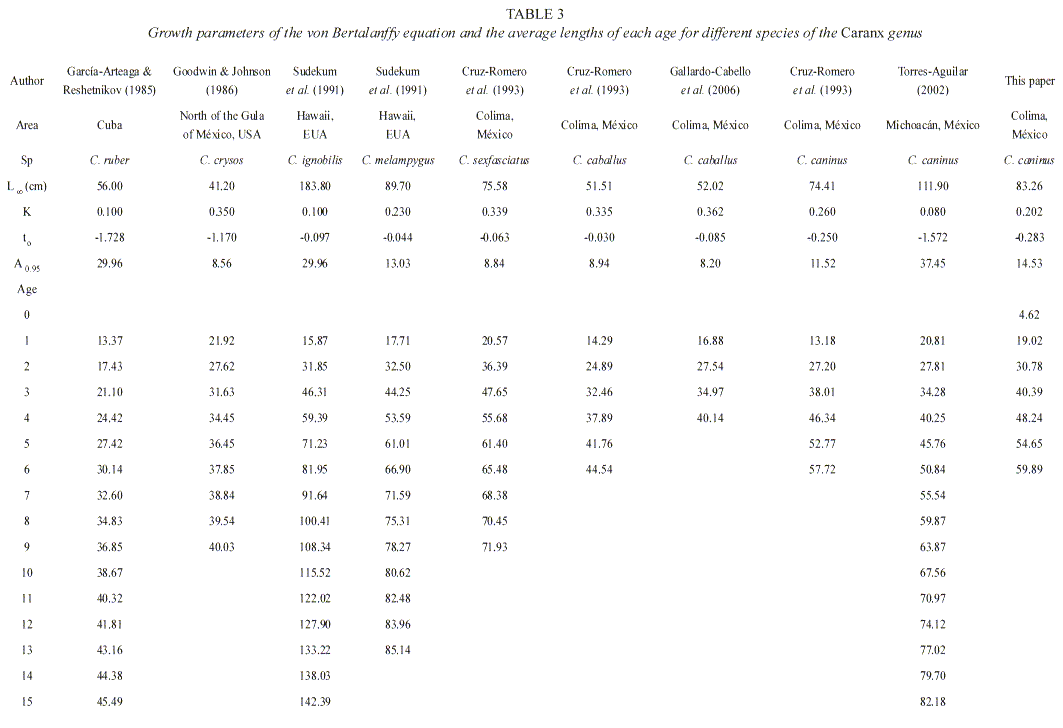
Theoretical value of longevity (Taylor 1958) is based in von Bertalanffy´s formula as 95 % of the asymptotic length (Taylor 1960). As with the growth parameters, longevity is very sensitive to the sample, that is, if the organisms are large, the asymptotic length increases and the growth coefficient decreases, whereas if the organisms present a large "K" value its asymptotic length is reached in a shorter period of time. In the present work, the value of longevity for the different species of carangids is from eight and beyond. Calculations for the Pacific jack in Colima are 12 and 15 years (Cruz-Romero et al. 1993 and this study, respectively), while the values for Guerrero and Michoacán (Torres-Aguilar 2002) reach up to 37 years.
Results on growth analysis of other species of Caranx genus were used, because of the lack of specific data (García-Arteaga and Reshetnikov 1985, Goodwin and Johnson 1986, Sudekum et al. 1991, Cruz-Romero et al. 1993, Torres-Aguilar 2002 and Gallardo-Cabello et al. 2006). For the different species, the value of the K index result in curves of steeper trends for larger specimens (0.26 to 0.35) and the length reaches the asymptotic length L∞ faster or in a shorter time (Table 3). This occurs with C. crysos a species with the highest growth rate which reaches 21.92 cm in its first year. Together with C. caninus (Torres-Aguilar 2002), C. ruber, C. ignobilis and C. melampygus these are the carangids with the highest longevity.
The GSI is an indicator of the gonad development and therefore of the reproductive cycle of the fish (Rodríguez-Gutiérrez 1992). The results indicate higher reproductive activity in May and November, different from C. caballus whose active months are June and October. This helps to avoid interspecific competition and achieve a successful spawning (Ibáñez-Aguirre 1995, Rojas-Herrera et al. 2004). These two species have similar characteristics and behavior. Morphologically the Pacific jack reaches larger sizes, but from the fishery point of view they are both coastal pelagic and are fished in the same way.
The three indexes: condition factor (a), gonadosomatic (IGS) and gastric repletion (IRG), help to assume that the main spawning periods occur in November and May, because during this period the corporal waste of the fish is higher and looses weight. The most active feeding periods coincide with the period of the fast growth rings in the otoliths, that is, during the months of August to February, period that also coincides with the rainy season (Observatorio de Manzanillo, pers. com.; Espino-Barr 2000) responsible for the wash of abundant nutriments to the rivers and to the coastal zones; the same is true for the hurricane season (Observatorio de Manzanillo, pers. com.; Espino-Barr 2000) which remove the marine bottoms of Colima´s narrow continental shelf, and sends nutrients to the surface of the ocean and increases food availability. During this time, the individual weight increases and a recovery of the reproductive population takes place (larvae and juveniles); also the recruitment to the fishing zone occurs.
Conversely, during the spring-summer period (March-August), the availability of the food diminishes the values of the condition factor index decreases and the slow growth rings are formed. However during this season a massive spawning period is also observed during May.
Resumen
El jurel Caranx caninus es una especie común que se captura en la pesca artesanal frente a las costas de Colima, México. Mensualmente, durante 2002, se tomaron datos morfométricos y muestras de otolitos para determinar la edad y el crecimiento. Se encontraron siete grupos de edad; durante el primer año de vida se da el mayor crecimiento, hasta 14.4 cm. Durante el segundo año, C. caninus crece 11.76 cm; el tercero 9.61 cm; el cuarto 7.85 cm; el quinto 6.41 cm y el último año crece 5.24 cm. Las constantes de la ecuación de crecimiento de von Bertalanffy fueron: L∞ = 83.26 cm, W∞ = 18.138 g, K = 0.202, to = -0.283 y la longevidad A0.95 = 15 años. Las curvas de crecimiento de otras especies del mismo género se compararon con las obtenidas en el presente trabajo. El índice gonadosomático presentó valores más altos durante noviembre y mayo. Los periodos de tiempo de alimentación más intensa son de agosto a febrero.
Palabras clave: Caranx caninus, edad y crecimiento, distribución de frecuencia de talla, sagittae, asterisci, ecuación von Bertalanffy, longevidad.
Received 02-IV-2006. Corrected 08-V-2007. Accepted 22-VIII-2007.
References
Aguirre-León, A. & S. Díaz-Ruiz. 2006. Estructura de tallas, madurez gonádica y alimentación del pez Diapterus rhombeus (Gerreidae) en el sistema fluviodeltaico Pom-Atasta, Campeche, México. Rev. Biol. Trop. 54: 599-611. [ Links ]
Blacker, R.W. 1974. Recent advances in otolith studies, p. 67-90. In J. Harden-Jones (ed.). Sea Fisheries Research, London, England. [ Links ]
Bussing, W.A. & M.I. Lopez. 2004. Peces de Isla del Coco y peces arrecifales de la costa pacifica de Centro América Meridional. Rev. Biol. Trop. 52: 1-191. [ Links ]
Castro-Aguirre, J.L. 1978. Catálogo sistemático de los peces marinos que penetran a las aguas continentales de México, con aspectos zoogeográficos y ecológicos. Depto. Pesca, INP, Serie Científica 19, México D.F., México. [ Links ]
Chirichigno, N., W. Fischer & C.W. Nawen . 1982. Catálogo de especies marinas de interés económico actual o potencial para América Latina. Infopesca, Pacífico Centro y Suroriental, Roma fao/pnud, Italy. [ Links ]
Cruz-Romero, M., E. Espino-Barr & A. Garcia-Boa. 1989. Lista de peces del litoral Colimense. INP/SEPESCA Serie: Doc. Trab. 1: 21 p. [ Links ]
Cruz-Romero, M., E. Espino-Barr & A. Garcia-Boa. 1993. Carángidos: aspectos biológico-pesqueros en el litoral colimense. Cuad. Mex. Zool. 1: 81-88. [ Links ]
Cruz-Romero, M., E. Espino-Barr & A. Garcia-Boa. 1996. Potencial de la pesca ribereña en el estado de Colima, México, en 1989. INP/SEMARNAP, Ciencia Pesquera 12: 30-35 [ Links ]
Espino-Barr, E., M. Cruz-Romero & A. Garcia-Boa. 2003. Peces marinos con valor comercial de la costa de Colima, México. CONABIO, INP, CRIP-Manzanillo, ISBN 970-9000-23-3, México D.F., México. [ Links ]
Espino Barr, E., E.G. Cabral Solís, A. Garcia Boa & M. Puente Gómez. 2004. Especies marinas con valor comercial de la costa de Jalisco, México. SAGARPA–INP, ISBN-968-800-570-3, México D.F., México. [ Links ]
Espino-Barr, E. 2000. Criterios Biológicos para la administración de la pesca multiespecífica artesanal en la costa de Colima, México. Tesis de Doctorado, Colima, México. [ Links ]
Espino-Barr, E., M. Gallardo-Cabello, A. Garcia-Boa, E.G. Cabral-Solís & M. Puente-Gómez. 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: sagitta, asteriscus and lapillus of Caranx caninus (Pisces: Carangidae) in the coast of Colima, Mexico. J. Fish. Aquat. Sci. 1: 157-170. [ Links ]
Ford, E. 1933. An account of the herring investigations conducted at Plymouth during the years from 1924 to 1933. J. Mar. Biol. Assoc. U.K. 19: 305-384. [ Links ]
Gallardo-Cabello, M., E. Espino-Barr, A. Garcia-Boa, E.G. Cabral-Solís & M. Puente-Gómez. 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: sagitta, asteriscus and lapillus of Caranx caballus (Pisces: Carangidae) in the coast of Colima, Mexico. Internat. J. Zool. Research 2: 34-47 [ Links ]
García-Arteaga, J.P. & Y.S. Reshetnikov. 1985. Age and growth of the barjack (Caranx ruber) off the coast of Cuba. J. Ichthyol. 25: 120-131. [ Links ]
Gayanilo, F.C., P. Sparre & D. Pauly. 1994. The FAOICLARM Stock Assessment Tools (FiSAT) User´s Guide. FAO Computarized Information Series (Fisheries), Rome, Italy. [ Links ]
Goodwin, J.M. & A.G. Johnson. 1986. Age, growth, and mortality of blue runner, Caranx crysos form the northern Gulf of Mexico. Northeast Gulf Sci. 8: 107-114. [ Links ]
Gulland, J.A. 1964. Manual of methods of fish population analysis. FAO Fish, Tech. Paper., Rome, Italy. [ Links ]
Jiménez-Badillo, L. 2006. Age-growth models for tilapia Oreochromis aureus (Perciformes, Cichlidae) of the Iniernillo reservoir, Mexico and reproductive behaviour. Rev. Biol. Trop. 54: 577-588. [ Links ]
Ibáñez-Aguirre, A.L. 1995. Algunos aspectos de la dinámica de poblaciones de Mugil cephalus (Linneo, 1758) y M. curema (1836) (Pisces: Mugilidae) en la laguna de Tamiahua, Veracruz. Tesis doctoral, Fac. de Ciencias, UNAM, México D.F., México. [ Links ]
Lucano-Ramírez, G., S. Ruiz-Ramírez & J.A. Rojo-Vázquez. 2006. Composición por tallas y ciclo reproductivo de Pseudupeneus grandisquamis (Pisces: Mullidae) en el Pacífico central Mexicano. Rev. Biol. Trop. 54: 195-207. [ Links ]
Mendenhall, W. 1987. Introduction to probability and statistics. PWS-Kent Publishing, Boston, Massachussets, USA. [ Links ]
Microsoft. 1992. Excel. Manual de usuario. Referencia de funciones. Microsoft, Redmond, Washington, USA. [ Links ]
Ramírez-Granados, R. 1977. Estudio de la factibilidad de una terminal pesquera en Manzanillo, Col. Depto. Pesca, México 2: 268-338. [ Links ]
Rodríguez-Gutiérrez, M. 1992. Técnicas de evaluación cuantitativa de la madurez gonádica en peces. AGT, México DF. [ Links ]
Rojas, M.J.R., E. Maravilla & F. Chicas B. 2004. Hábitos alimentarios del pargo mancha Lutjanus guttatus (Pisces: Lutjanidae) en Los Cóbanos y Puerto La Libertad, El Salvador. Rev. Biol. Trop. 52: 163-170. [ Links ]
Rojas-Herrera, A.A., M. Mascaró & X. Chiappa-Carrara. 2004. Hábitos alimentarios de los peces Lutjanus peru y Lutjanus guttatus (Pisces: Lutjanidae) en Guerrero, México. Rev. Biol. Trop. 52: 959-972. [ Links ]
Safran, P. 1992. Theoretical analysis of the weight-length relationship in fish juveniles. Mar. Biol. 112: 545- 551. [ Links ]
Saucedo-Lozano, M., G. Gónzalez Sansón & X. Chiappa Carrara. 1999. Alimentación Natural de jóvenes de Lutjanus peru (Nichols y Murphy, 1922) (Lutjanidae: Perciformes) en la costa de Jalisco y Colima, México. Ciencias Marinas 25: 381-400. [ Links ]
Sierra, L.M. & O.A. Popota. 1988. Velocidad de digestión, ritmo diario de la alimentación y raciones alimenticias de algunas especies de peces del Golfo de Batabanó, Cuba. Instituto de Oceanología, Acad. Cien. Cuba, La Habana, Cuba. [ Links ]
Sudekum, A.E., J.D. Parrish, R.L. Radtke & S. Ralston. 1991. Life history and ecology of large jacks in undisturbed, shallow, oceanic communities. Fish. Bull. U.S. 89: 493-513. [ Links ]
Taylor, C.C. 1958. Cod growth and temperature. J. Conseil 23: 366-370. [ Links ]
Taylor, C.C. 1960. Temperature, growth and mortality–the Pacific cockle. J. Conseil 26: 177-124. [ Links ]
Torres-Aguilar, M. 2002. Estudio preliminar de edad y crecimiento del jurel, Caranx caninus (Günther, 1867), con base en vértebras y espinas, en las costas de Guerrero y Michoacán. M.Sc. Thesis, UNAM, México. [ Links ]
van der Heiden, A.M. 1985. Taxonomía. Biología y evaluación de la ictiofauna demersal del Golfo de California, p.149-200. In A. Yánez-Arancibia (ed.). Recursos pesqueros potenciales de México. La Pesca acompañante del camarón. Progr. Univ. de Alimentos. ICMyL, UNAM, México D.F., México. [ Links ]
von Bertalanffy, L. 1938. A quantitative theory of organic growth (inquiries on growth laws. Human Biology 10:181-213. [ Links ]
Walford, L.A. 1946. A new graphic method of describing the growth of animals. Biol. Bull. 90: 141-147. [ Links ]
Zar, J.H. 1996. Biostatistical analysis. Prentice Hall, New Jersey, USA. [ Links ]


{kind=link}