










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 suppl.2 San José Dec. 2006
Ultraestructura de bambúes del género Dendrocalamus (Poaceae: Bambusoideae) cultivados en Costa Rica I: Dendrocalamus latiflorus
Mayra Montiel1,2 & Ethel Sánchez2
1 Facultad de Ciencias Agroalimentarias, Escuela de Zootecnia, Universidad de Costa Rica. Ciudad Universitaria Rodrigo Facio, San Pedro de Montes de Oca. Apdo. Postal 2060, San José, Costa Rica; mmontiel@cariari.ucr.ac.cr
2 Centro de Investigación en Estructuras Microscópicas (CIEMIC), Universidad de Costa Rica. Apdo. Postal 2060, San José, Costa Rica. Fax: (506) 207-3182; ethels@cariari.ucr.ac.cr
Recibido 15-VI-2006. Corregido 15-VII-2006. Aceptado 20-VIII-2006.
Abstract: The bamboo Dendrocalamus latiflorus is easily affected by sudden changes in temperature and humidity, which cause it to bloom. We used a scanning electron microscope to describe the ultrastructure of young culm (2 years old), culm bracts, buds, abaxial surface of the leaf lamina and flowers. The most noticeable ultrastructural features were the presence of two types of operculated cells in the sterile flower glums, the abundant cuticular wax, the presence of reticulated punctuations in the xylem and pollen grains with interlocking disks or cups. The operculated cells and the interlocking disks are taxonomically important features. Rev. Biol. Trop. 54(Suppl. 2): 43-50. Epub 2006 Dec. 01.
Key words: ultrastructure, scanning electron microscopy, anatomy, bamboo, Poaceae, Dendrocalamus latiflorus.
La especie Dendrocalamus latiflorus Munro 1868, es originaria de Taiwán y fue introducida a Costa Rica en 1985 por Wei-Chih Lin. Desde entonces, ha adquirido importancia económica por la gran demanda que han tenido sus retoños de delicado sabor y sus culmos maderables.
La clasificación taxonómica de esta especie fue realizada por Lin (1981), y en la actualidad los estudios utraestructurales dan muchos aportes en este aspecto, en especial los de las células epidérmicas, que por sus características, ayudan mucho en la identificación de géneros y especies de la familia Poaceae (Prat 1936, Tateoka et al. 1959, Metcalfe 1960, Jacques-Félix 1962).
Dichas características estructurales de las gramíneas han sido tradicionalmente estudiadas por medio de microscopía de luz, sin embargo, hace aproximadamente 30 años, se comenzó a utilizar el microscopio electrónico de barrido (SEM, por sus siglas en inglés). Palmer (1976) realizó estudios de cutícula tanto en materiales frescos como en fragmentos fosilizados; Tucker (1981) estudió la epidermis de gramíneas del este de África y más recientemente Bisen (1999) analizó la epidermis de bambúes de la India. Estos estudios contribuyeron a mejorar el conocimiento de la anatomía y taxonomía de la subfamilia Bambusoideae.
Aunque se han realizado estudios de epidermis de la hoja y culmos en muchos géneros y especies de bambúes de la India (Bisen et al. 1988, Bisen 1999), aún hay muchos de los cuales no se conocen en detalle esas estructuras, por ello, el objetivo de este trabajo fue describir la ultraestructura al SEM de los culmos, brácteas del culmo, láminas foliares, yemas y flores de ejemplares de D. latiflorus cultivados en Costa Rica.
El análisis de las flores y sus estructuras fue posible debido a condiciones ambientales adversas, que provocaron la floración por estrés de los ejemplares utilizados. En condiciones normales la frecuencia de floración de los bambúes puede variar desde cada tres años (Schizostachyum elegantissimum ) hasta cada 120 años (Phyllostachys bambusoides ) (Seethalakshmi y Kumar 1998).
Materiales y métodos
Secciones de culmos, brácteas del culmo, láminas foliares, yemas y flores de D. latiflorus fueron recolectadas en la Finca Río Frío, Heredia, Costa Rica, a 100 msnm. Este sitio presenta un tipo de suelo areno-limoso, una precipitación anual de 4 120 mm, una temperatura promedio de 25 ºC y una humedad relativa de 85-95%.
Las muestras fueron fijadas en una solución de glutaraldehído (2.5%) y paraformaldehído (2%) en amortiguador de fosfato de sodio (0.1 M, pH 7.4), por 24 horas a 4°C. Luego se lavaron con el amortiguador y se posfijaron con tetraóxido de osmio (2%), por una hora. Posteriormente, se lavaron tres veces con agua destilada, se deshidrataron en un gradiente ascendente de etanol (30-50%) y se secaron utilizando terbutanol en un secador por sublimación. Una vez secas, se montaron en bases de aluminio y utilizando un cobertor iónico (Eiko I-D 2) se cubrieron con 30 nm de oro, y se observaron con un microscopio electrónico de barrido (Hitachi S-570).
Resultados
Culmo: La epidermis tiene gran cantidad de cera cuticular, así como la presencia de células epidérmicas reticulares, tricomas en forma de gancho y estomas rodeados de papilas céricas (Fig. 1A). En la figura 1B se observa gran cantidad de papilas céricas así como las bases de los tricomas que han desaparecido. Las células tienen diferentes patrones de distribución y parecen reticuladas.
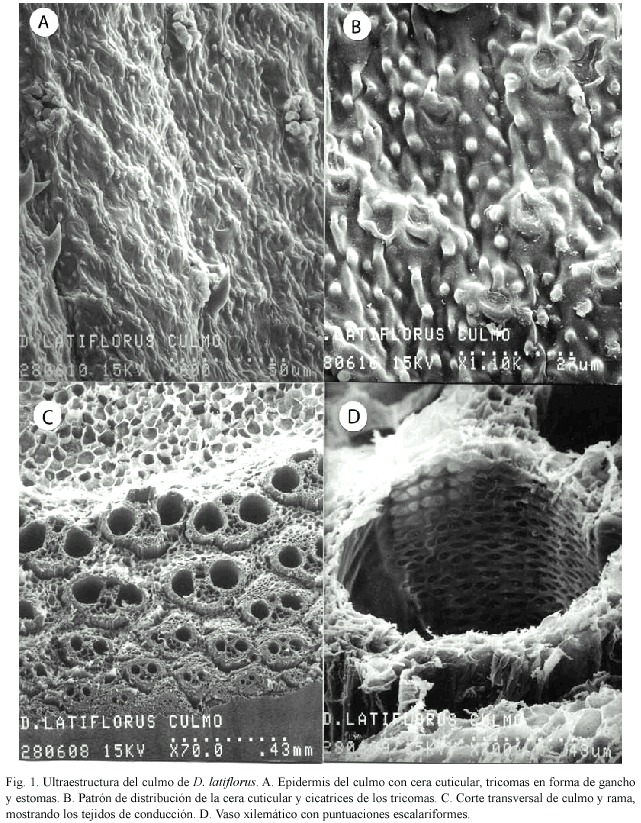
Por otra parte, los cortes transversales de culmo y rama joven tienen el lumen de tejido parenquimatoso medular que en la madurez desaparece, y los haces vasculares que se reagrupan y retraen en la periferia del culmo y rama (Fig. 1C). El haz vascular presente en esta especie, es de tipo III y está constituído por dos vasos de metaxilema, floema, parénquima, fibras y tejido esclerenquimatoso. Los vasos xilemáticos presentan la pared con puntuaciones reticuladas (Fig. 1D).
Bráctea del culmo: La superficie está recubierta casi por completo por una gran cantidad de tricomas unicelulares; también es posible observar tricomas en forma de gancho, cera cuticular y estomas rodeados de papilas céricas (Fig. 2A). En un corte transversal se observa el tejido parenquimatoso y parte de la epidermis externa (Fig. 2B). Las células epidérmicas largas se alternan con los estomas y las células subsidiarias de éstos se ven claramente (Fig. 2C). En la figura 2D se observan tricomas unicelulares largos, tricomas en forma de gancho y cera cuticular.
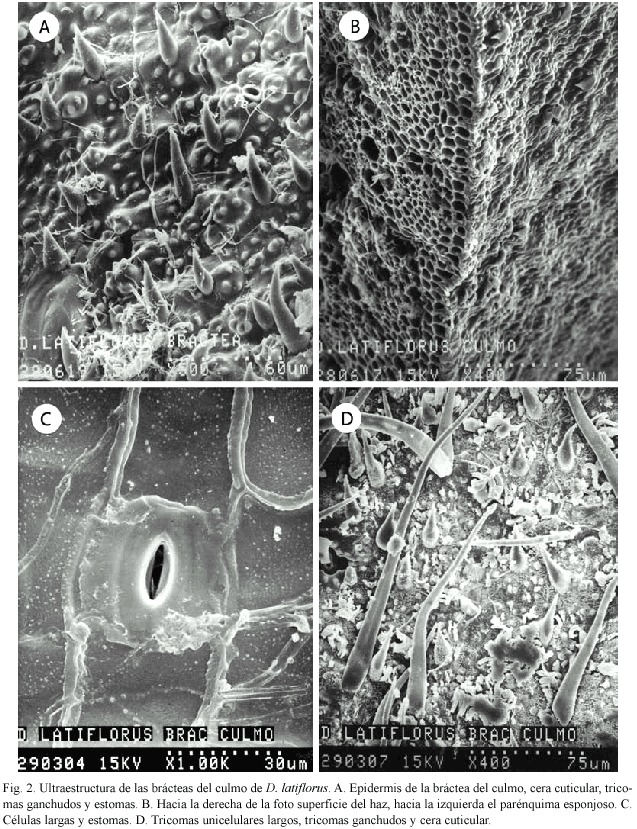
Lámina foliar: La superficie abaxial de la lámina foliar presenta pequeñas cantidades de cera cuticular, tricomas en forma de gancho y estomas de alto domo (Fig. 3A).
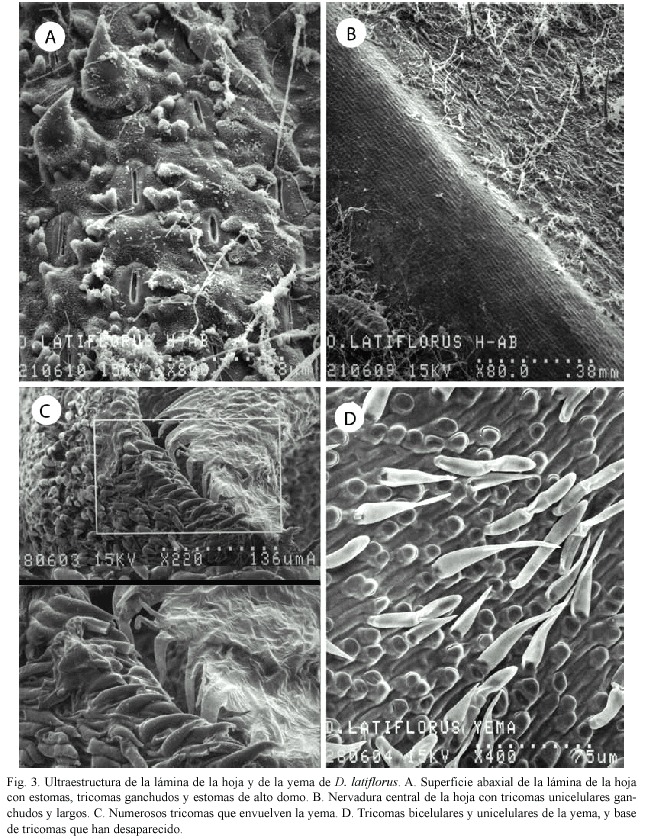
La nervadura central corresponde a la banda vascular central o área costal. En las partes laterales a ella o áreas intercostales, se observaron tricomas unicelulares largos, en poca cantidad, y cera (Fig. 3B).
Yema: En las yemas examinadas el ápice presenta múltiples tricomas unicelulares (Fig. 3C). Los tricomas unicelulares y bicelulares se observan muy bien, así como las bases de los tricomas que han desaparecido (Fig. 3D).
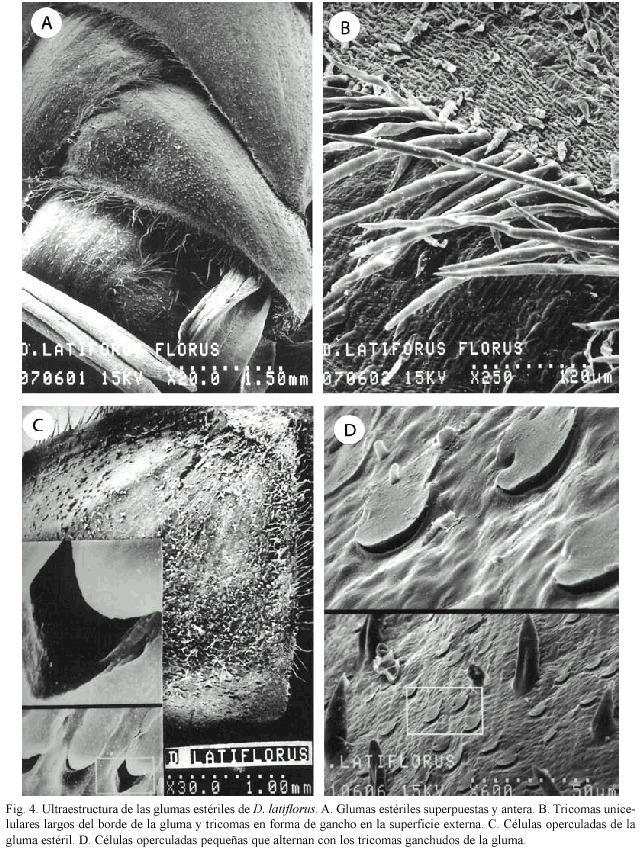
Flor: Las flores se encuentran protegidas por estructuras denominadas glumas, las cuales al tacto son coriáceas y se superponen una sobre otra para envolver las estructuras florales que están en el centro (Fig. 4A). Las glumas estériles tienen en sus bordes numerosos tricomas alargados y tricomas en forma de gancho en su superficie externa (Fig. 4B). También en ellas se observan algunas estructuras similares a "papilas o valvas", las cuales son abundantes y podrían denominarse "células operculadas" (Fig. 4C). Alternando con los tricomas en forma de gancho, aparecen otras estructuras más pequeñas de diferente forma a las anteriores, que parecieran ser células de sílice y que tienen una abertura lateral similar a la de las valvas (Fig. 4D).
Las anteras, constituidas por tejido parenquimatoso, son ditecicas porque tienen cuatro sacos polínicos en dos tecas, y cada saco polínico aloja en su interior los granos de polen (Fig. 5A).
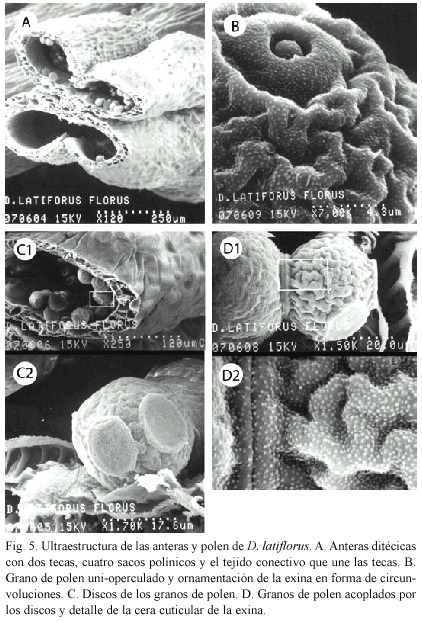
Los granos de polen son esferoidales uni-operculados y la ornamentación de su exina es en forma de circunvoluciones; además presenta una distribución ordenada de las papilas céricas, que se aprecian como puntos blancos (Fig. 5B). Sobre la exina hay discos semejantes a ventosas, los cuales permiten un acople firme entre granos de polen contiguos (Figs. 5C, 5D).
Discusión
El culmo de D. latiflorus presenta gran cantidad de cera cuticular, la forma de las papilas céricas es esferoidal y su distribución muy similar a las de la lámina de la hoja de Dendrocalamus asper, clon Taiwán. Además sus vasos xilemáticos tienen puntuaciones reticuladas.
Las brácteas del culmo son ásperas al tacto por la gran cantidad de tricomas, los estomas en ellas sólo se pudieron observar al eliminar la cera cuticular. La lámina foliar y la yema presentan menos cantidad de cera que las estructuras anteriores, pero tienen abundancia de tricomas. Estas características son muy parecidas a las de otros bambúes. Debido a la gran abundancia de cera cuticular fue imposible observar las células de sílice en el culmo y bráctea, pero tampoco se logró en la lámina foliar a pesar de presentar menor cantidad de cera.
Las flores de esta especie sorprendieron por la presencia en sus estructuras de células muy diferentes, tal es el caso de las "células operculadas" de las glumas estériles, las cuales podrían tener función de intercambio de gases o líquidos; así como los granos de polen con discos o ventosas de acople, quizá para una mayor eficiencia en la dispersión anemófila. En un estudio ultraestructural realizado por Montiel y Kozuka (1994) en gran número de pólenes de gramíneas, nunca se encontraron estos discos ni una ornamentación de la exina tan pronunciada. Por lo general, la literatura indica que en la familia Poaceae la exina no presenta diferencias significativas, siendo D. latiflorus una excepción.
Agradecimientos
Se agradece a la vicerrectoría de Investigación de la Universidad de Costa Rica el apoyo dado a este proyecto, así como al Centro de Investigación en Estructuras Microscópicas de la Universidad de Costa Rica por las facilidades brindadas. Este trabajo se realizó con la ayuda financiera de la Comunidad Europea, en el marco del Proyecto Guadua: Research for Sustainable Management and Markets of Bamboo in Colombia and Costa Rica (EU INCO ICA4-CT-2001-10091).
Resumen
Utilizando el microscopio electrónico de barrido, se evaluaron ultraestructuralmente, culmo joven (dos años de edad), brácteas del culmo, yemas, superficie abaxial de la lámina foliar y flores de Dendrocalamus latiflorus, un bambú susceptible a cambios bruscos de temperatura y precipitación, que lo hacen florecer con facilidad. Los rasgos más notables fueron la presencia de dos tipos de células operculadas en las glumas estériles de las flores, la abundancia de cera cuticular, la presencia de puntuaciones reticuladas en el xilema y los granos de polen especializados, por la presencia de discos o ventosas para su acople. Las células operculadas y los discos de acople son caracteres taxonómicos importantes.
Palabras clave: ultraestructura, microscopia electrónica de barrido, anatomía, bambú, Poaceae, Dendrocalamus latiflorus.
Referencias
Bisen, S. S., G. P. Mishra & S. C. Sharma. 1988. Scanning electron microscopy studies on culm and leaf epidermis of Indian Bamboos. Indian For. 114: 656-669. [ Links ]
Bisen, S. S. 1999. Identification and classification of Indian Bamboos (SEM Atlas of epidermis). Bishen Singh Mahendra Pal Singh, Dehra Dum, India. 200 p. [ Links ]
Jacques-Félix, H. 1962. Les Graminées d`Afrique Tropicale I: généralités, classification, description des genres. Institute de Reserche Agronomiques Tropicales et des Cultures vivientes, Paris, Francia. 345 p. [ Links ]
Lin, W. C. 1981. Subfamily Bambusoideae, p. 706-783. In Flora of Taiwan. vol. 5. Reprinted Epoch, Taipei, Taiwan. [ Links ]
Metcalfe, C. R. 1960. Anatomy of the monocotyledons. I. Gramineae. Oxford Clarendon, Oxford, Inglaterra. 731 p. [ Links ]
Montiel, L. M. & Y. Kozuka. 1994. Polen de gramíneas y sus aspectos botánicos, taxonómicos y ecológicos. Rev. Biol. Trop. 42 (Supl. 1): 21-39. [ Links ]
Palmer, P. G. 1976. Grass cuticles: A new paleoecological tool for East African lake sediments. Can. J. Bot. 54: 1725-1734. [ Links ]
Prat, H. 1936. La Systematique des Graminées. Ann. Sci. Nat. Bot. Ser. 10 (18): 165-258. [ Links ]
Seethalakshmi, K. K. & M. S. Muktesh Kumar. 1998. Bamboos of India: A compendium. Brill, Leiden, Holanda. 342 p. [ Links ]
Tateoka, T., S. Inoue & S. Kawano. 1959. Notes on some grasses IX: Systematic significance of bicelular microhairs of leaf epidermis. Bot. Gaz. 121: 80-91. [ Links ]
Tucker, A. E. 1981. A scanning electron microscope survey of the epidermis of East African Grasses I. Smithsonian Contrib. Bot. 49: 1-84. [ Links ]

