










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 n.4 San José Dec. 2006
Ingemar Hedström1, Jessica Harris2 & Kimberly Fergus3
1 Boston University, College of Arts and Sciences, 725 Commonwealth Avenue, Boston, MA 02215, USA. Present address: Nairi Foundation, Apdo. 150-4021 Orotina, Costa Rica; ingemar.hedstrom@skutan.smf.se
2 Tufts University, College of Liberal Arts and Sciences, Medford, MA 02155, USA; jessica.harris@tufts.edu
3 Northern Arizona University, College of Arts and Sciences, Flagstaff, AZ 86011, USA; kmfkat@aol.com
Key words: orchid bees, Euglossa, deforestation, organic agriculture, bio-indicator, Costa Rica.
The onset of commercial agriculture and increasing urbanization has a considerable impact on natural ecosystems throughout the developing world. In Costa Rica, the national economy relies heavily upon agricultural exports, where coffee was the first successful export and today remains the number two crop for exportation, just behind bananas. Forest ecosystems across the country have been progressively converted to commercial farms and pasturelands. Besides the altered water flow, soil erosion, and loss of biodiversity caused by this deforestation (Schelhas 1996), the negative implications of conventional agriculture include pesticide residues, loss of soil productivity, and groundwater contamination. According to Humphries et al. (1995), biological diversity around the world is being lost at 1 000 times the natural extinction rate as a result of the alteration of natural habitats, pollution, and natural habitat degradation caused by human activities. This manipulation of the land is creating environmental disturbances that should be quantified.
Saunders et al. (1990) state that integrated landscape management is critical for conservation of biodiversity on a macroscale. Organic farming, in which agroecology is considered in the context of sustainable production techniques, is one such alternative. The International Federation of Organic Agricultural Movements (IFOAM) defined organic farming as agricultural systems that promote environmentally, socially, and economically sound production (Stolton et al. 2000). These systems value local soil fertility within natural ecological cycles, and prohibit the use of synthetic chemical fertilizers, pesticides, and pharmaceuticals in an effort to limit harmful external inputs.
Beginning in the 1970s, organic agriculture has progressively gained esteem throughout the world as a viable means of production as well as a healthier alternative for both humans and the natural environment (Hedström et al., 2006). Due to growing international markets for organic products, including coffee, organic farming in Costa Rica has emerged as a financially viable and “eco-friendly” alternative to conventional farming.
In order to quantitatively compare the environmental health of organic and conventional farming practices in Costa Rica, effective biological indicators for this habitat needs to be established. The use of biological indicators is one of the simplest forms of habitat assessment. Biological indicators are used as measurable surrogates for monitoring environmental health. According to Noss (1990), reliable indicators should be: sensitive to provide an early warning of change; distributed over a broad geographic area; provide a continuous assessment over a wide range of states; independent of sample size; easy and effective to measure and collect; and able to differentiate between natural cycles or trends and those induced by anthropogenic stress. As suggested by Roubik and Hanson (2004), orchid bees are supreme research subjects. With the advent of artificial baits for attracting males, orchid bees have become easy to study, and their relatively large size and spectacular colors also make them more observable than most insects.
Male orchid bees are proposed here as potential biological indicators of tropical ecosystems, specifically the ecosystems of organic and conventional coffee plantations. According to Roubik and Hansson (2004), the Euglossinae subfamily (Hymenoptera; Apidae), is composed of five genera (Aglae, Eufriesea, Euglossa, Eulaena, and Exaerete). Euglossa, with 40 species in Costa Rica (only three endemic species), contains a total of 103 described species in the Neotropics. They are long distance pollinators of low to middle- elevation plants across mainland tropical America, comprising 20-30 % of the bee community’s species in lowland forests (Ackerman 1983, Roubik and Hanson 2004). According to Dressler (1982), large numbers of males may be collected without seriously affecting the reproduction and survival of the population.
Male euglossine bees possess a highly sensitive sense of smell (Schiestl and Roubik 2003) that may allow them to detect chemical disruptions in their habitat such as synthetic pesticides and herbicides. They are highly attracted to perfumes of plants other than orchids, which they collect on specialized hairs on their forelegs and hind tibias for unknown use (Roubik and Hanson 2004). They have a memory for scents relative to their location and can trace the origin of an odor from up to 1 km away (Dressler 1982). According to a study by Ackerman (1983) in central Panama, species richness and bee abundance are correlated; both fluctuate seasonally and peak in the early wet season. Individuals are known to fly long distances, up to 2.5 km, in response to a perfume bait (Dressler 1968), and according to Janzen (1981), will seek a variety of resources in different and distant habitats when their original habitat becomes seasonally severe. Under ordinary circumstances, every orchid bee can cover an area of over 1 000 km2 a day (Roubik and Hansen 2004). Janzen (1981) suggests that distances in excess of 20 km are normal for foraging, and greater distances are feasible when resources become scarce. However, according to Ackerman (1983), species composition, evenness, and dominance ranks were virtually non-seasonal, so the male euglossine bee community seems to have some structural continuity.
The hypothesis of our study was based on two assumptions: 1) organic farms are healthier ecosystems than conventional farms, as organic farms seek to mimic the natural diversity of unaltered habitat, and 2) orchid bees may serve as viable biological indicators of ecosystem health because they exhibit many of the aforementioned qualities of biological indicators. variables such as seasonality has been taken into account to project the most effective conditions for using euglossine bees as a biological indicator of agricultural ecosystem health. We expected to demonstrate that organic agriculture is more effective than conventional agriculture in maintaining tropical ecological health.The results of the study conducted in August 2004 (mid-rainy period), were compiled with the previous findings from April 2004 (late-dry period) and June 2004 (early-rainy period). Our hypothesis is that more bees will be found within the organic farm site, implying the organic agriculture methods are less disruptive to the functions of a natural ecosystem.
Materials and methods
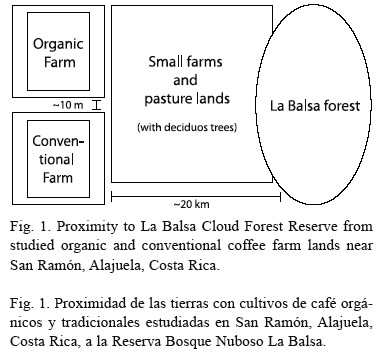
Data was collected on two adjacent coffee farms, La Paz (organic) and Cardena Farm (conventional), located near the Northern Barranca River (Norte del Río Barranca), San Ramón, Alajuela Province, Costa Rica (10º 06’ N; 84º 28’ W). Both farms are at an elevation of about 1 000 m above sea level, with an average temperature of 21 °C. There is an approximate 10 m “buffer zone” between the two farms (Fig. 1), with coffee plants inside the zone still considered conventionally-farmed coffee. The Corn Plant (Dracaena fragrans), “caña india” in Spanish, originally introduced from West Africa, is used to bolster the buffer zone. Both farms use a variety of shade tree species, including the Coral Bean (Erythrina sp.) and Guamo (Inga sp.), commonly known in Costa Rica as “poró” and “guaba”, respectively, which enrich the soil through nitrogen fixation. The farms are about 20 km from La Balsa Cloud Forest Reserve, which gradually changes into old secondary forest (Fig. 1). During the dry season (December-April) the patch of deciduous trees on the studied farms and La Balsa forest is relatively bare, yet the wet season (May-November) experiences an explosion of life as the dormant trees produce new growth.
Data were collected on April 20-21, June 29-30, and August 3-4, 2004, at four stations on each farm (eight total data collection sites) using chemical baits to attract male euglossine bees. Each local study station was spaced 50 m apart (April and June, 2004), and 100 m apart (August 2004), and 50 m into the farm from the edge. The distance between the two studied coffee farms was about 1 km apart. Eucalyptus oil (cineole 1:8), a pheromone substitute, was applied with an eyedropper to a coffee filter located on a tree at breast height (approx. 1.5 m). Five drops were reapplied to the coffee filter every 15 min. It was ensured that the drops were fully absorbed by the filter paper and did not smear any areas of the trees. The maximum number of bees observed at each local station at one time during each 15 min interval was recorded for 1.5 h. Bees had to be within approximately 0.5 m of the filter in order to be counted. A t-test was used to statistically determine differences in numbers of bees appearing on the two farms.
Results
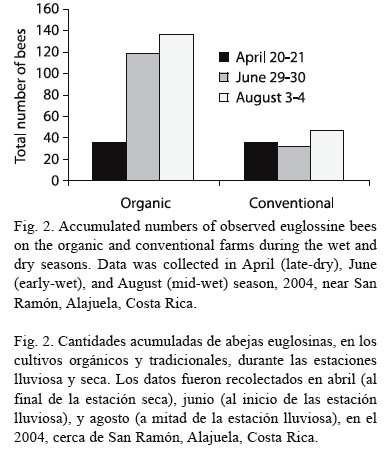
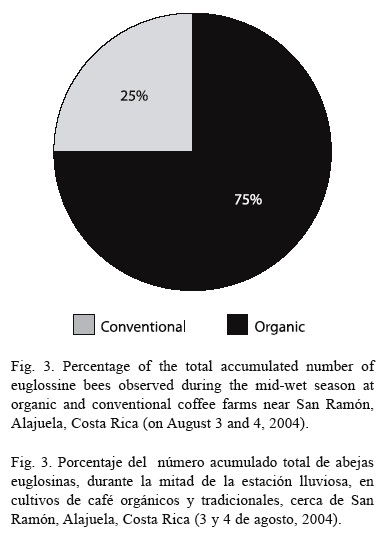
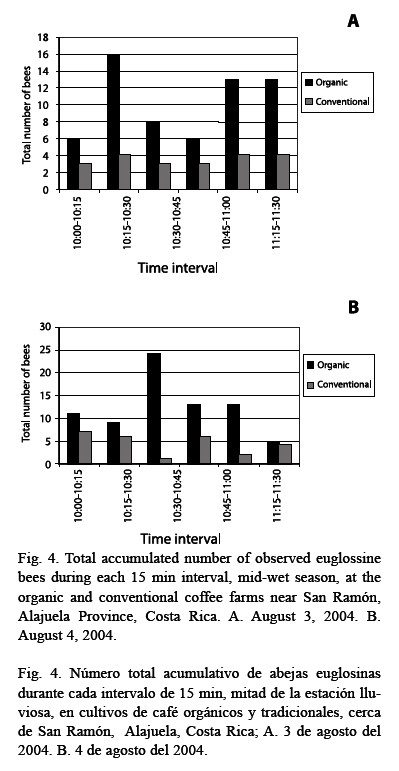
The total accumulated numbers of observed euglossine bee during the late-dry season (April 2004) were the same (Fig. 2). However, in both studies during the wet season (June 2004 and August 2004), a higher number of bees was observed in the organic as compared to the conventional farm (Fig. 2). The highest cumulative number of bees was observed within the organic farm during mid-wet season (August 2004, cf. Fig. 3). On August 3, a total of 62 bees were observed within the organic farm, while 21 bees were seen in the conventional farm. The following day, a total of 75 bees were observed at the organic farm land, while 26 bees were observed at the conventional farm (Fig. 4).
The t-calculation for August 3, 2004 was 4.57 with df= 6 and the tc = 2.015 (p= 0.003). The to for August 4 was 2.482, the df was 9, and the tc was 2.015 (p= 0.028). The null hypothesis could be rejected because the to values were greater than the tc values.
In each of the two days in August 2004, a higher average number of bees on the organic farm was observed at one time per 15 min interval. On August 3, an average of 10 bees was seen on the organic farm while an average of three bees was seen on the conventional farm (Fig. 2). The following day, an average of 13 bees was seen on the organic farm while an average of four bees was seen on the conventional farm (Fig. 3). On the second day, observations for the last 15 min (11:15-11:30) occurred under light rain showers.
Discussion
Data from the mid-wet season (August, 2004) demonstrated significant support for the hypothesis that euglossine bees are effective indicators of farm health during Costa Rica’s wet season. Hence the findings show a much higher abundance of euglossine bees within the organic farm, and the t-test further supports the hypothesis. When the results from late-wet (April) and early-wet (June) seasons were taken into account, more intricate conclusions linked to seasonality were drawn. The most significant difference in abundance between the two farms was found in the studies during the wet season. The months of July and August are in the midst of the Costa Rican wet season.
When observations were conducted during the late-dry season in April 2004, there was no difference in bees observed on the organic and conventional farms (Fig. 4). By the end of June 2004 (early-wet season), however, a considerable difference in bee abundance was found between the two farms, with many more bees observed in the organic environment. The August 2004 data (mid-wet season) continued this trend. These findings suggest that orchid bees in lowland Costa Rica were most abundant during the midst of the wet season.
Previous studies by Roubik (1993, 2001) have shown that higher numbers of male bees may occur in “hot spots”, presumably due to physical factors, nearby concentration of resources, or nests and male resting or display sites. Any absence in their populations after a major forest clearing seemed to not be due to a suddenly restricted habitat size (Becker et al. 1991, Cane 2001), but to a high level of disturbance (Roubik and Hanson 2004). Once the process of regeneration begins and disturbance diminishes, the bees may spread to formerly isolated patches. If food and nesting resources become especially abundant, secondary vegetation may even support a higher diversity of euglossines (Roubik and Hanson 2004). Therefore, it is not necessarily the size or age of the vegetation, but rather the resource quality, which matters to euglossine bees. Seasonality is directly related to this variable, as the wet season provides more forest and vegetation cover for bees to travel long distances. The remarkable stability of orchid bee populations, even in the face of ongoing deforestation, may make them ideal to use to monitor Costa Rican agroecology. However, male euglossines seem to make effective biological indicators only if that their habitat has not been altered to the extent that they cannot travel to the monitored area.
In conclusion, male euglossine bees were observed using a known attractant (cineole 1:8) at organic and conventional coffee farms. These observations have shown orchid bees to be a viable bio-indicator of organic farm health on a seasonal basis. Hence, orchid bees may offer an effective means for measuring the health of an ecosystem. They are particularly useful during the rainy season, when the area between farm and forest is green, providing a biological corridor by which the bees may travel. It is important to note that both farms sit 20 km from the nearest forest (Fig. 1) and use the same diversity of shade trees (Erytrina sp. and Inga sp.) and coffee plants, yet the orchid bees still were more abundant in the organic farm. Since the orchid bees have a acute sense of smell, it is possible that the unnatural odors of agricultural chemicals used in conventional coffee farms interfere with their search for natural fragrances. Our findings demonstrated that euglossine bees may have a natural preferencefor undisturbed habitat, and thus will choose to forage in an organic coffee farm over a conventional coffee farm. However, seasonality and its corollary, forest cover, are critical variables of this assessment. It seems that euglossine beeswill only travel in large numbers during the wet season, traveling through deciduous foliagebetween the forestandthe farm. It was onlyduring this wet season migration that a clearpreference for organic versus conventional farm ecosystems was shown. Therefore, it isquitepossible that orchid bees are an effective biological indicator of organic coffee farmsduring the wet season only.
Acknowledgments
The authors gratefully acknowledge the support provided by the Center for Sustainable Development Studies, the School for Field Studies, SFS, Atenas, Costa Rica. We thank G. Sahlén, Uppsala University, Sweden, and two anonymous reviewers for constructive comments of an earlier version of the manuscript, and E. Brier-Rosenfield, A. Crane, E. Gladkova, A. Gordon, S. Minnerly, M. Mills, O. Carter, S. Cashman, E. Kelly, A. Martin, T. Seekircher, L. Tinder, for their assistance in the field.
Resumen
Con el propósito de entender las implicaciones de la agricultura orgánica (en comparación con la tradicional) para el “estado de salud ambiental”, usamos aceite de eucalipto (1:8-cineole), para monitorear la abundancia de machos de abejas de las orquídeas (euglosinas) como posibles bioindicadores. Comparamos una finca de café orgánico (La Paz) y una tradicional (La Carena), cerca del Río Barranca en San Ramón de Alajuela, Costa Rica. Las tomas simultáneas de datos fueron realizadas en ambas fincas en abril del 2004 (a finales de la estación seca), en junio del 2004 (a principios de la estación lluviosa) y en agosto del 2004 (a mediados de la estación lluviosa). Estas abejas pueden ser bioindicadores viables de la salud de las fincas orgánicas de café (en comparación con las tradicionales) durante la estación lluviosa. Durante la estación seca no se mostró una diferencia significativa en la abundancia de las abejas de orquídeas, entre una y otra finca. Hubo un número significativo de euglosinas en las fincas orgánicas durante la estación lluviosa; sugiriendo que el aumento está vinculado con la estacionalidad y el acceso al bosque en su hábitat natural.
Palabras clave: abejas de las orquídeas, Euglossa, deforestación, agricultura orgánica, bioindicadores, Costa Rica.
References
Ackerman, J.D. 1983. Diversity and seasonality of male euglossine bees (Hymenoptera:Apidae) in Central Panama. Ecology 64: 274-283.
[ Links ]Becker, P., J.S. Moure & F.J.A. Peralta 1991. More about euglossine bees in Amazonian forest fragments. Biotropica 23: 586-591.
[ Links ]Cane, J.H. 2001. Habitat fragmentation and native bees: a premature verdict? Conservation Ecology 5 (also available online: www.consecol.org/vol5/iss1/art6).
[ Links ]Dressler, R.L. 1968. Pollination by euglossine bees. Evolution 22: 202-210.
[ Links ]Dressler, R.L. 1982. Biology of the orchid bees (Euglossini). Ann. Rev. Ecol. Syst. 13: 373-394.
[ Links ]Hedström, I., A. Denzel & G. Owens. 2006. Orchid bees as bio-indicators for organic coffee farms in Costa Rica: Does farm size affect their abundance? Rev. Bio. Trop. 54(3): 965-969.
[ Links ]Humphries, C.J., P.H. Williams & R.I. Vane-Wright. 1995. Measuring biodiversity value for conservation. Ann. Rev. Ecol. Syst. 26: 93-111
[ Links ]Janzen, D.H. 1981. Bee arrival at two Costa Rican female Catasetum orchid inflorescences, and a hypothesis on euglossine population structure. Oikos 36: 177-183.
[ Links ]Noss, R.F. 1990. Indicators for monitoring biodiversity: a hierarchical approach. Cons. Biol. 4: 355-364.
[ Links ]Roubik, D.W. 1993. Tropical pollinators in the canopy and understory: field data and theory for stratum “preferences”. J. Ins. Beh. 6: 659-673.
[ Links ]Roubik, D.W. 2001. Ups and downs in pollinator populations: where is there a decline? Conservation Ecology 5: 2 (also available online: www.consecol.org/vol5/iss1/art2).
[ Links ]Roubik, D.W. & P.E. Hanson. 2004. Orchid bees of tropical America: Biology and field guide. Instituto Nacional de Biodiversidad (INBio), Heredia, Costa Rica. 370 p.
[ Links ]Saunders, D., R.J. Hobbs & C.R. Margules. 1990. Biological consequences of ecosystemfragmentation: a Review. Cons. Biol. 5: 18-19.
[ Links ]Schelhas, J. 1996. Land use choice and change: intensification and diversification in the lowland tropics of Costa Rica. Human Org. 55: 298-306.
[ Links ]Schiestl, F.P. & D.W. Roubik. 2003. Odor compound detection in male euglossine bees. J. Chem. Ecol. 29: 253-257.
[ Links ]Stolton, S., B. Geier & J.A. McNeely (eds.). 2000. The relationship between nature conservation, biodiversity, and organic agriculture, p. 224. In The International Federation of Organic Agriculture Movement (IFOAM), Tholey-Thekey, Germany. [ Links ]

