










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 n.2 San José Jun. 2006
Estructura de tallas, madurez gonádica y alimentación del pez Diapterus rhombeus (Gerreidae) en el sistema fluvio-deltaico Pom-Atasta, Campeche, México
Arturo Aguirre-León1 & Silvia Díaz-Ruiz2
1 Universidad Autónoma Metropolitana-Xochimilco. Departamento El Hombre y su Ambiente. Calzada del Hueso 1100, Col. Villa Quietud, México 04960, D.F. Fax: (52) 54 83 74 69; agla1269@correo.xoc.uam.mx
2 Universidad Autónoma Metropolitana-Iztapalapa. Departamento de Hidrobiología. Apartado Postal 55-535, México 09340, D.F. Fax: (52) 58 04 47 38; sdr@xanum.uam.mx
Recibido 07-III-2003. Corregido 11-II-2004. Aceptado 09-VI-2004.
Abstract. Size structure, gonadic development and diet of the fish Diapterus rhombeus (Gerreidae) in the Pom-Atasta fluvial-deltaic system, Campeche, Mexico. The fish Diapterus rhombeus was studied during an annual cycle from 1992 to 1993 in the fluvial-deltaic Pom-Atasta system associated with Terminos Lagoon, Campeche, Mexico. It is a dominant species in the system, based on its numeric abundance, weight, high frequency and wide distribution. A total of 745 individuals were obtained, with a weigth of 2 890.2 g and length ranging from 3.0 to 16.7 cm. The annual variation of the allometric coefficient b was from 2.71 to 3.345. The condition factor varied from 0.711 to 0.934. The statistical analysis shows significant differences (p< 0.05) between the seasons of the year and the habitats of the system for the weight, the longitude and the condition factor K, which reflects the space-temporal utilization of the system for the species. The population present at Pom-Atasta, consists mainly by juvenile and few preadults individuals in gonadal stages I, II, and III, and more females than males were recorded. This species utilizes the system as a nursery area, growth and feeding area. It has a varied trophic spectrum, and consumes at least eight different groups. Its principal food items are undetermined organic matter, foraminifers, ostracods and tanaidaceans. It is a first order consumer. The Pom-Atasta system is located in a zone of intense fishing and oil activity, so it is important to advance in the knowledge of its fishing resources. Rev. Biol. Trop. 54(2): 599-611. Epub 2006 Jun 01.
Key words: Diapterus rhombeus, biology, ecology, Pom-Atasta, Gulf of Mexico.
La costa de Campeche en el Sur del Golfo de México representa una región costera prioritaria para realizar estudios de evaluación ecológica de sus recursos pesqueros, debido a que es la segunda zona pesquera más importante de dicho país, con gran actividad industrial, incluyendo la explotación petrolera. En la costa de Campeche, la Laguna de Términos y los sistemas fluvio-deltaicos asociados, contienen una gran variedad de recursos pesqueros que se explotan de manera artesanal (Yáñez-Arancibia y Aguirre-León 1988). El sistema fluvio-deltaico Pom- Atasta es parte de la región de la Laguna de Términos, en donde las poblaciones de peces utilizan los hábitats del sistema durante sus ciclos de vida, integrando una compleja comunidad en diversidad, distribución, abundancia y tramas tróficas (Aguirre-León et al. 1998). Diversos estudios realizados en dicha región, han mostrado que Diapterus rhombeus (Cuvier) tiene una amplia distribución y abundancia en los distintos ambientes de la región de Términos, incluyendo los sistemas fluvio-deltáicos asociados a ella (Amezcua-Linares y Yáñez-Arancibia 1980, Aguirre-León y Yáñez-Arancibia 1986, Vera-Herrera et al. 1988, Ayala-Pérez et al. 2001) como también en la Sonda de Campeche (Yáñez-Arancibia y Sánchez-Gil 1986) y en el sistema Pom-Atasta (Ayala-Pérez et al. 1993, Aguirre-León et al. 1993, Cabrera-Díaz 1995, Gómez-Montes 1997 y Aguirre-León et al. 1998). El conocer las características de la población de esta mojarra en el sistema estudiado, ayudará a detectar cambios en su comportamiento biológico y ecológico, debidos a las alteraciones del sistema provocadas por la degradación del ambiente causada por el hombre. Es importante señalar que el sistema Pom-Atasta, esta sujeto a diversos impactos ambientales, debidos en parte a la construcción de infraestructura petrolera conectada con la Sonda de Campeche, dragados para el tendido de tuberías que transportan gas natural a través de la Laguna de Pom. Asimismo, por la tala de manglar para exploración petrolera, instalación de plataformas de perforación, fabricación de carbón y postes para vivienda, como también por los asentamientos humanos en la Laguna de Atasta que alberga cooperativas pesqueras. Debido al poco conocimiento sobre la biología y ecología de D. rhombeus en el sistema Pom- Atasta, la finalidad de este trabajo es analizar la estructura de tallas, la relación talla-peso, factor de condición, madurez gonádica y hábitos alimenticios de esta fracción de la población, en diferentes épocas climáticas así como en diferentes localidades del sistema, discutiendo la utilización del sistema como parte de su ciclo biológico. Estos aspectos permitirán generar conocimiento sobre el estado actual de la población en el sistema Pom-Atasta y a su vez podrá ser utilizado por tomadores de decisiones para su conservación en la zona costera de Campeche.
Material y métodos
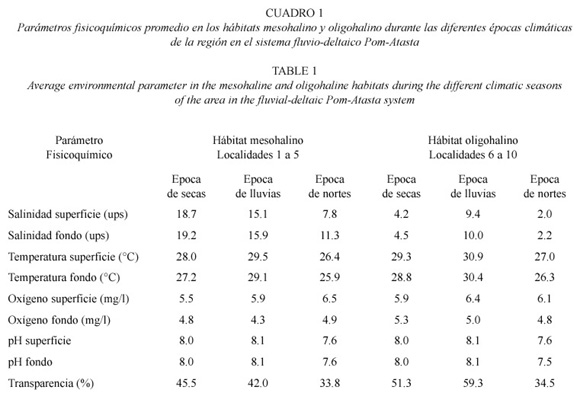
Area de estudio: El sistema fluvio-deltaico Pom-Atasta se localiza en la región occidental de la Laguna de Términos entre 18º30 y 18º35 N y 91º50 y 92º20 W (Fig. 1A). Esta ubicado dentro de la llanura costera de Campeche formada por los ríos Grijalva y Usumacinta (Gutiérrez- Estrada et al. 1982). Es paralelo a la costa con superficie aproximada de 190 km2, longitud de 50 km, profundidad promedio de 2.7 m y formado por lagunas interiores de dimensiones variables. En Pom-Atasta se establece un gradiente semipermanente este-oeste de salinidad (28.5 a 0 ups), temperatura (22 a 34°C), oxígeno disuelto (1 a 10 mg/l) y transparencia (11 a 92%), debido al efecto de la marea y la morfología del sistema y el intercambio de masas de agua con la Laguna de Términos. Estas características morfológicas y ambientales determinan la existencia de dos hábitats bien definidos durante todo el año, un hábitat mesohalino (localidades 1 a 5) y un oligohalino (localidades 6 a 10) (clasificación salina, según Carriker 1967), (Fig. 1B, Cuadro 1). Amplios detalles sobre la hidrología de este sistema se encuentran en Aguirre-León et al. (1998). La vegetación costera esta caracterizada por extensos bosques de manglar de Rizophora mangle L., Avicenia germinans L. y Laguncularia racemosa Gaertn. (De la Lanza-Espino 1993). La vegetación tipo pasto arraigado al fondo es escasa y se encuentra principalmente en la Laguna de San Carlos (Halodule wrightii Ascherson) y en las Lagunas de Atasta y Pom (Vallisneria americana Michaux) (Barreiro- Güemes y Aguirre-León 1995). En esta región del Golfo de México se presentan tres temporadas climáticas: época de secas de febrero a mayo, temperatura ambiente promedio 28.1ºC, precipitación promedio 164.8 mm; época de lluvias de junio a septiembre, temperatura 28.3ºC y precipitación 976.7 mm, en ambas temporadas predominan vientos del sureste; y época de nortes (tormentas de invierno) de octubre a enero, temperatura 24.4ºC, precipitación 345.6 mm, con fuertes vientos del norte (Yáñez-Arancibia y Day 1988).
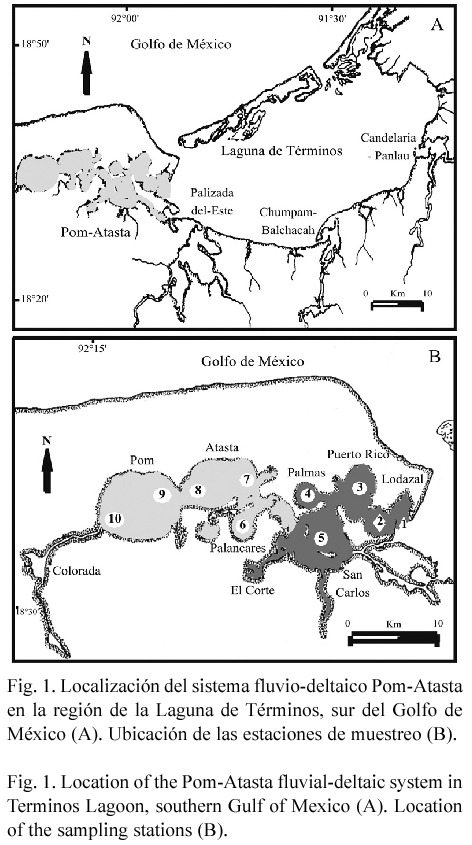
Se realizaron 13 campañas mensuales de campo de abril 1992 a abril 1993, en diez localidades de muestreo distribuidas a lo largo del Sistema Pom-Atasta. Estas se ubicaron considerando el aporte y flujo del agua dulce, el tipo de sedimento, la vegetación sumergida, el gradiente estuarino del sistema y la conexión con la Laguna de Términos (Fig. 1B). Los recolectas de peces se efectuaron con una red de arrastre de prueba camaronera de 5.0 m de largo, 2.5 m de abertura de trabajo y luz de malla de 1.88 cm, equipada con tablas de 0.5 x 0.8 m, en una lancha de fibra de vidrio de 7.0 m de eslora y motor fuera de borda de 55 HP. En cada estación se efectuó un lance con duración de 20 min, a una velocidad promedio de dos nudos, cubriéndose un área de arrastre de 3000 m2 y totalizando 130 arrastres en el periodo de estudio. Los individuos capturados se fijaron con formol al 10% para su posterior procesamiento en el laboratorio. Se realizaron mediciones de temperatura, salinidad (termosalinometro YSI), y oxígeno disuelto del agua en superficie y fondo (oxímetro YSI), transparencia (disco de sechii) y profundidad (sondaleza), así como observaciones de vegetación sumergida.
Se estimó la relación talla-peso de la población por medio de la ecuación potencial: P=aLb (Ricker 1975) y el factor de condición utilizando la expresión K= P(100)/Lb (Pauly 1984); donde P= Peso (g), L= longitud total (cm), a= ordenada al orígen y b= pendiente de regresión; K= factor de condición, b= exponente. La determinación de sexo y madurez gonádica de los individuos se realizó de acuerdo con el criterio de Nikolsky (1963). Se calculó el índice gonadosomático (IGS) para las hembras usando la expresión: IGS=Wg(100)/Wt (Rossemblum et al. 1987); donde IGS= índice gonadosomático, Wg= peso de la gónada, Wt= peso del individuo. Para el análisis de la alimentación y hábitos alimenticios se utilizaron los métodos: numérico N= n(100)/N, donde N= porcentaje numérico de un alimento dado, n= suma de los elementos de un alimento en todos los estómagos, N= suma de los elementos de los alimentos en todos los estómagos; gravimétrico P= p(100)/P, donde P= porcentaje en peso de un alimento particular, p= suma del peso de este alimento en todos los estómagos, P= suma del peso del contenido estomacal de todos los estómagos; y de frecuencia F=ne(100)/Ne, donde F= frecuencia de aparición de un tipo de alimento, ne= número de estómagos con un tipo de alimento, Ne= número de estómagos no vacíos examinados (Yáñez-Arancibia et al. 1986). La identificación de los taxa alimenticios se hizo hasta donde fue posible según el grado de digestión del alimento. Fue calculado el índice de repleción IR= Pc/Pt para estimar el grado de llenado de los estómagos, donde IR= índice de repleción, Pc= peso contenido estomacal, Pt= peso total del estómago (Molina-Ocampo 1993). Los parámetros de la población, se compararon entre épocas del año y hábitats del sistema utilizando ANOVA con significancia p< 0.05 (Daniel 1997), de resultar diferencias significativas, se determinó entre que épocas y hábitats mediante la prueba de Tukey (Montgomery 1984).
Resultados
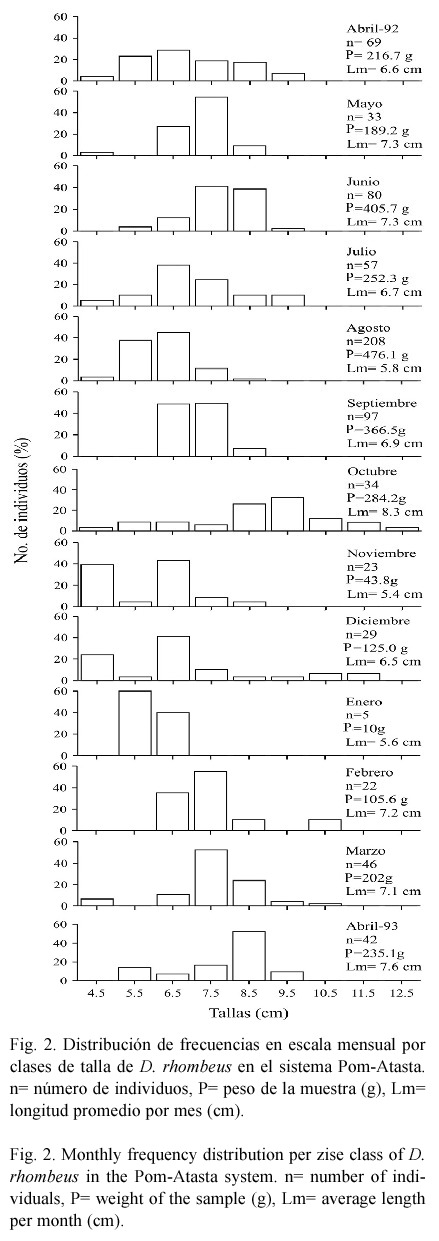
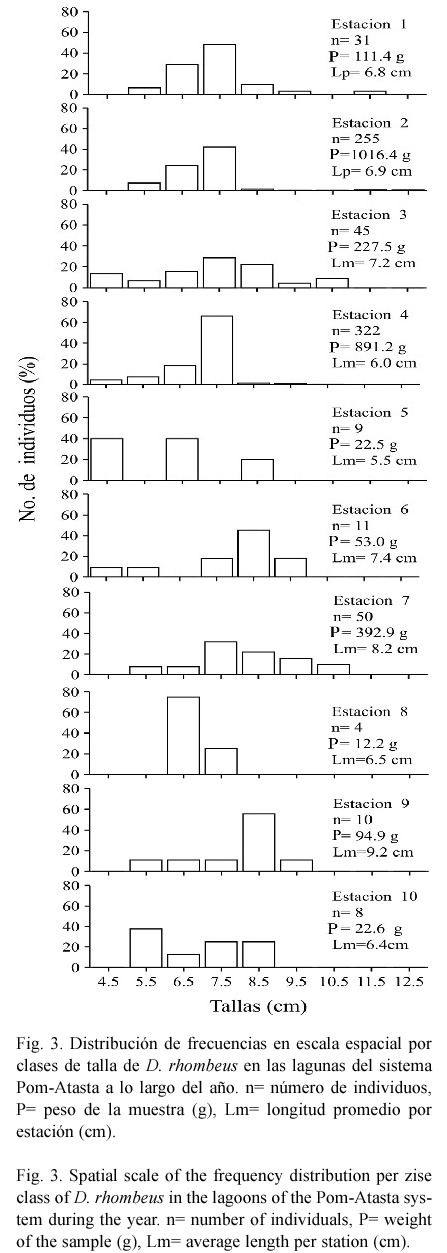
Estructura de tallas. Se recolectaron 745 individuos con un peso total de 2 890.2 g, intervalo de longitud total de 3.0 a 16.7 cm y longitud promedio de 6.6 cm. La población estuvo representada principalmente por individuos juveniles y escasos preadultos. En escala temporal (Fig. 2) la captura total de la temporada de lluvias (junio a septiembre) fue la mayor con 442 individuos, un peso total de 1 510.3 g, longitud de 3.0 a 10.0 cm, con promedio de 6.4 cm. Para la temporada de secas (febrero a mayo) la captura total fue de 212 individuos, con un peso de 923.0 g, longitud de 3.3 a 16.7 cm, con promedio de 7.1 cm. En esta temporada, solo dos individuos registrados en mayo presentaron las longitudes más grandes (14.4 y 16.7 cm). En la temporada de nortes (octubre a enero) se obtuvo la menor captura con 91 individuos, un peso de 456.9 g, longitud de 3.5 a 12.4 cm con promedio de 6.8 cm. De acuerdo con ANOVA se encontraron diferencias significativas entre las épocas del año para el peso y la longitud, la prueba de Tukey señaló que fueron entre secas con lluvias (peso, p=0.00033 y longitud, p=0.00002) y entre lluvias con nortes (peso, p=0.000026 y longitud, p=0.0275). En escala espacial a lo largo del año (Fig. 3), D. rhombeus se registró con mayor abundancia en las lagunas del hábitat mesohalino (Fig. 1) (estaciones 1 a 5, variación annual promedio de salinidad de 4.1 a 23.3 ups), con 662 individuos, un peso de 2 315.1 g, longitud de 3.0 a 10.5 cm con promedio de 6.5 cm. En las lagunas del hábitat oligohalino (estaciones 6 a 10, variación anual promedio de salinidad de 0.9 a 12.4 ups) se capturaron 83 individuos, con un peso de 575.1 g, longitud de 4.3 a 16.7 cm con promedio de 8.0 cm. En estas lagunas, los dos individuos con mayor longitud fueron capturados en la estación nueve. El ANOVA indicó diferencias entre los hábitats para el peso y la longitud, la prueba de Tukey señaló que fueron para peso, P= 0.000009 y para longitud, P= 0.000009.
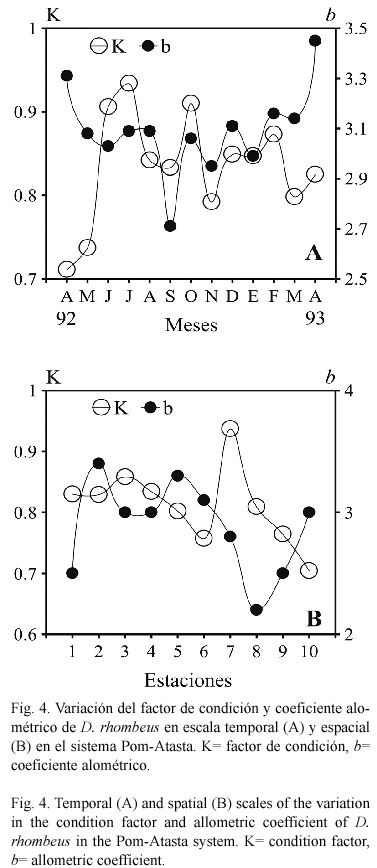
Relación talla-peso y factor de condición. La variación temporal y espacial de estos parámetros se integra en la figura 4. El coeficiente alométrico b varió a lo largo del año de 2.71 en septiembre 1992 a 3.45 en abril 1993 (Fig. 4A). El valor anual de b para la población fue de 3.168, con una r2 de 0.950. La variación mínima y máxima del factor de condición promedio (K) a lo largo del año, fue de 0.711 en abril (época de secas) a 0.934 en julio de 1992 (época de lluvias) respectivamente, con un valor promedio anual de 0.846. Los valores promedio de K fueron de 0.789 en la temporada de secas, de 0.879 para la de lluvias y de 0.850 en la de nortes. El ANOVA indicó diferencias entre las épocas del año para K, la prueba de Tukey señaló que fueron entre secas con lluvias (p=0.000022), entre lluvias con nortes (p=0.000088) y entre nortes con secas (p=0.000022). En escala espacial, el coeficiente alométrico b varió de 2.504 a 3.494 en las lagunas del hábitat mesohalino (Fig. 4B), y de 2.951 a 3.151 en el oligohalino. La variación del factor de condición a lo largo del sistema fue de 0.704 a 0.937, con un promedio anual de 0.812. En las lagunas de mayor salinidad el valor promedio fue de 0.831, y en las lagunas más dulces fue de 0.794 (Fig. 4 B). El ANOVA indicó diferencias entre los hábitats para K, la prueba de Tukey señaló que p=0.000009.
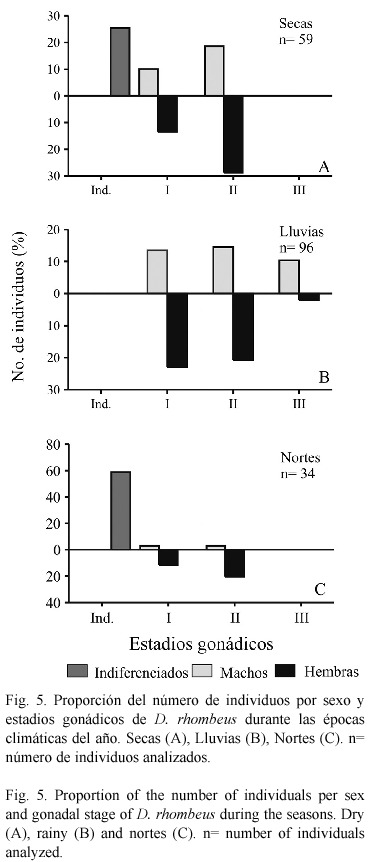
Madurez gonádica. La frecuencia de sexos y estadios gonádicos para cada época climática se resumen en la figura 5, donde se observan desde individuos indiferenciados hasta estadio III, variando el número de individuos para cada estadio en relación con la temporada climática. En la época de secas, predominó una mayor proporción de hembras en estadio II, así como organismos indiferenciados, la proporción macho / hembra fue de 1.0 / 1.5. En la época de lluvias se presentaron hembras en estadios I y II, siendo mayor la proporción del estadio I, además solo en esta temporada fueron registrados pocos individuos en fase III, la proporción macho / hembra fue de 1.0 / 1.2. Para la época de nortes aumento el número de organismos indiferenciados y disminuyo la proporción de estadios I y II en ambos sexos, la proporción macho / hembra fue de 1.0 / 2.2. En el sistema, D. rhombeus fue más frecuente en el hábitat mesohalino (localidades 1 a 5) en estadios I, II y sólo en la laguna del Pom (estación 7) correspondiente al hábitat oligohalino se encontraron organismos en estadio III (nueve machos y dos hembras). No se observaron valores altos del IGS para las hembras, este vario de 0.10 a 0.89 durante las épocas climáticas, excepto para octubre en la época de nortes (IGS = 2.0), lo que sugiere que la especie ocupa el sistema principalmente como área de crianza y alimentación. En el sistema el índice IGS fue mayor en el hábitat oligohalino (0.16 a 1.6), debido a la presencia de algunos individuos en fase III; en el hábitat mesohalino vario de 0.01 a 0.44. La madurez gonádica en relación con la longitud total de los individuos en ambos sexos, fue para el estadio I de 3.0 hasta 8.0 cm, para el estadio II de 5.5 hasta 8.0 cm y para el estadio III de 8.2 hasta 10.0 cm.
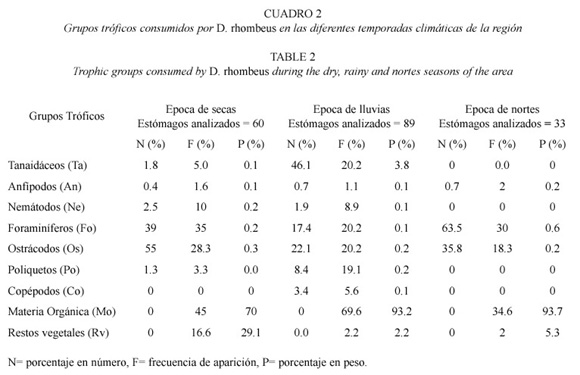
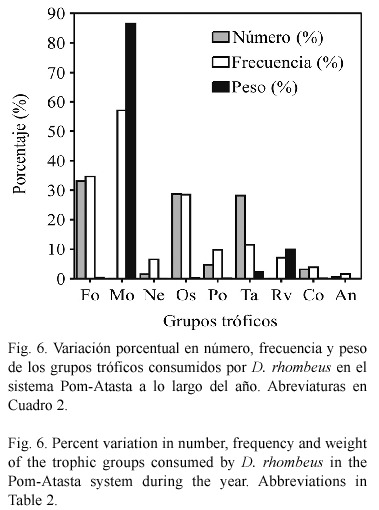
Hábitos alimenticios. El alimento se compone al menos de ocho grupos tróficos a través del año, presentando algunas variaciones en cada época climática y localidad del sistema (Cuadro 2). Los valores porcentuales de estos grupos en escala anual se informan en la figura 6. Las presas más abundantes en número, peso y frecuencia, en escala espacial y temporal fueron: materia orgánica no determinada (Mo), foraminíferos (Fo), ostrácodos (Os) y tanaidáceos (Ta). Para cada una de las temporadas climáticas, estos grupos representaron en número el 94% en la época de secas, 39.5% en la de lluvias y 99.3% en la época de nortes; por peso el 70.5% en secas, 93.4% en lluvias y 94.5% en nortes y por frecuencia de 28.3 a 45% en secas, de 20.2 a 69.6% en lluvias y de 18.3 a 34.6% en nortes. En escala anual las mismas presas representan el 61.8% en número, el 87% en peso, y de 17.7 a 57.1% en frecuencia, por lo que estos son los grupos tróficos más importantes en la dieta de D. rhombeus. Con base en los valores del índice de repleción (0.142 a 0.8), se encontró un 89% de estómagos semillenos a lo largo del ciclo analizado, lo que sugiere que el sistema representa un área importante de alimentación.
Discusión
D. rhombeus utiliza los diferentes sistemas fluivo-deltaicos de la región, la Laguna de Términos y la Sonda de Campeche para desarrollar distintas etapas de su ciclo de vida. Es una especie bien adaptada a las condiciones estuarinas y marinas de esta región costera, por lo que es posible capturarla de manera regular en los hábitats de estos ecosistemas. Su mayor abundancia numérica y biomasa se ha registrado hacia la porción occidental de la Laguna de Términos donde se ubica el sistema Pom- Atasta (Aguirre-León et al. 1982, Aguirre-León y Yáñez-Arancibia 1986, Cabrera-Díaz 1995), y la salinidad de esta zona es menor debido a las grandes descargas de agua dulce proveniente de los sistemas fluvio-deltaicos Palizada del Este y Chumpam-Balchacah. En esta zona de la Laguna de Términos, Ayala-Pérez et al. (2001) encontró que su mayor abundancia se asocia con la presencia de vegetación sumergida sobre todo hacia el litoral interno de la Isla del Carmen. En la Sonda de Campeche también se registró con más abundancia hacia la misma región hasta una profundidad promedio de 40 m (Yáñez-Arancibia y Sánchez-Gil 1986). En Pom-Atasta, D. rhombeus estuvo representado en su mayoría por individuos juveniles y una proporción muy baja de preadultos, por lo que utiliza el sistema para desarrollar etapas tempranas de su ciclo de vida como crianza, crecimiento y alimentación.
La estructura mensual y espacial de tallas de D. rhombeus (Fig. 2 y 3) refleja a través del tiempo y del gradiente ambiental del sistema, que prevalece una población juvenil a lo largo del año, capturándose solo dos individuos con mayor talla en la época de secas (salinidad promedio de 10.3 ups) y hacia las lagunas de baja salinidad (promedio de 5.5 ups). No obstante lo anterior, se encontraron diferencias (p< 0.05) para la longitud y el peso entre las épocas del año y entre los hábitats, lo cual refleja las adaptaciones de la especie a las condiciones hidrológicas que prevalecen a lo largo del año y del gradiente ambiental del sistema. Esta predominancia de juveniles ha sido observada de manera similar en otros sistemas fluvio-deltaicos de la región (Amezcua-Linares y Yáñez-Arancibia 1980, Reséndez- Medina 1981, Vera-Herrera et. al. 1988, Nitsh 1992, Hernández-Sánchez 1994) como en hábitats de la Laguna de Términos donde la salinidad también es baja (Vargas-Maldonado et al. 1981, Aguirre-León y Yáñez-Arancibia 1986). Ayala-Pérez et al. (2001) informó para la Laguna de Términos, individuos de 2.5 a 18.0 cm de longitud total con moda de 7.5 cm, con predominancia de tallas de 7.5 a 9.0 cm, las cuales correspondieron también a juveniles. Asimismo, esta mojarra ha sido informada en otros sistemas costeros del Golfo de México (Reséndez-Medina y Kobelkowsky 1991, Matheson y Gilmore 1995) y Mar Caribe (González 1985, Arenas-Granados y Acero 1992, Cervigón 1993). En la costa de Brasil Araujo y Santos (1999), enfatizaron que para la Bahía de Sepetiba predominan juveniles de 1.5 a 7.5 cm la mayor parte del año, comportamiento que también se observo de manera similar en el sistema Pom-Atasta en Campeche.
Los valores del coeficiente alométrico b, como los de K a lo largo del año, se relacionan con la estructura de tallas y la abundancia numérica de juveniles en escala espacial y temporal (Figs. 2, 3 y 4A). Esto refleja cómo D. rhombeus se adapta temporalmente al sistema, ya que los valores promedio más bajos de K correspondieron a la época de secas, y los más altos a lluvias y nortes, encontrándose diferencias entre las épocas (P< 0.05). Los valores promedio de K entre los hábitats son parecidos (Fig. 4B) pero igualmente bajos, registrándose el mayor número de individuos juveniles y la talla promedio más pequeña en las lagunas de mayor salinidad (estaciones 1 a 5). No obstante, que en la Laguna de Atasta se registraron dos individuos de tallas mayores como lo muestra la talla promedio de la estación 9 (Fig. 3), se encontró diferencia significativa entre los hábitats para los valores de K (p< 0.05). Aguirre-León y Yáñez-Arancibia (1986) encontraron un comportamiento similar del coeficiente b en escala temporal y espacial para el litoral interno de la Isla del Carmen y para otros sistemas fluvio- deltaicos de la Laguna de Términos, con predominancia de juveniles. Ayala-Pérez et al. (2001), informó valores del coeficiente b cercanos a 3.0 para un ciclo anual en la Laguna de Términos, mencionando pulsos de K entre marzo y septiembre, sin embargo, no acotó los valores de estos picos. Por su parte, González (1985) para Isla Margarita, Venezuela, informó individuos de 9.9 a 21.2 cm de LT, un valor anual de b de 2.75 y valores altos de K (mayores a 1.0) para tallas de 11.0 a 16.0 cm para machos y hembras desde octubre hasta marzo.
La presencia mayoritaria de juveniles de D. rhombeus representados principalmente por estadios de madurez I y II en todo el año, indica que esta especie utiliza el sistema principalmente como área de crianza, crecimiento y alimentación, ya que fueron identificados individuos de sexo indiferenciado con mayor abundancia en las épocas de nortes y secas, y los valores del factor de condición son bajos durante todo el año (Fig. 5). Un comportamiento parecido fue informado por Hernández-Sánchez (1994) para el sistema Chumpam-Balchacah, al oeste del Pom-Atasta. Debido a que no se registraron hembras maduras de esta especie, los valores del IGS son bajos a lo largo del año, por lo que este comportamiento sugiere que D. rhombeus utiliza otros ambientes de la región para madurar. Etchevers (1978) informó valores de IGS mayores a 4.0 en hembras maduras para septiembre en Isla Margarita, Venezuela. En este estudio el valor más alto (2.0) se encontró para hembras en estadio III en octubre. Al parecer la especie migra a zonas someras de la plataforma continental adyacente a la Laguna de Términos donde realiza su reproducción. Esto fue propuesto para el sur del Golfo de México por Aguirre-León y Yáñez-Arancibia (1986), ya que en el sistema lagunar de Términos, estos autores identificaron pocos individuos en estadio III y ninguno en fase IV. Por su parte Yáñez- Arancibia y Sánchez-Gil (1986) encontraron mayor abundancia de esta especie hacia la porción occidental de la plataforma somera del Campeche, con individuos de 12.5 cm de longitud promedio, sin embargo, no realizaron análisis gonádico. Al respecto Austin (1971) para aguas de Puerto Rico y Etchevers (1978) en Isla Margarita, Venezuela, encontraron individuos en estadio IV y en reproducción a partir de los 12.5 cm de longitud total, registrándose entre septiembre y diciembre su desove más intenso. Asimismo, Araujo y Santos (1999) para la costa en Brasil, observaron que los individuos maduros se mueven hacia la plataforma somera adyacente para desovar entre noviembre y marzo y el reclutamiento a zonas protegidas se lleva a cabo de febrero a mayo. Por lo anterior, es razonable suponer que D. rhombeus tenga un amplio periodo de desove durante varios meses en áreas más profundas de la plataforma continental somera de Campeche en las épocas de nortes y secas, ya que como se observo en la figura 5, la presencia de individuos indiferenciados sugiere desove en otras áreas adyacentes al sistema Pom-Atasta. A pesar de la amplia distribución e importancia artesanal de esta especie en lagunas costeras mexicanas del Golfo de México, los estudios sobre su madurez, reproducción y su relación con variables hidrológicas todavía son incompletos. En el sistema estudiado, se encontró una mayor abundancia de individuos en el hábitat mesohalino (4.1 a 23.3 ups), donde la temperatura del agua varió de 25.3 a 29.4Oc y la transparencia de 18.7 a 58.6%.
El espectro trófico de D. rhombeus en Pom-Atasta es poco variado, pero presenta algunos cambios entre las diferentes épocas climáticas (Cuadro 2). Las presas más abundantes en número, peso y frecuencia que consume esta especie fueron encontradas en todas las tallas analizadas, debido a la mayor frecuencia de individuos juveniles de esta especie durante el año. En la época de lluvias se observo la presencia de copépodos en la dieta de esta mojarra, así como la ausencia de tanaidáceos, nemátodos y poliquetos en la época de nortes. Con base en los hábitos alimenticios de esta especie, fue considerada un consumidor de primer orden en el sistema. Estudios realizados sobre la alimentación de D. rhombeus en la Laguna de Términos señalan que se alimenta al menos de 20 grupos tróficos, entre los que destacan la materia orgánica no determinada, copépodos, ostrácodos, poliquetos, foraminíferos y nemátodos (Aguirre-León y Yáñez-Arancibia 1986). En la Laguna de Tampamachoco, Veracruz, se señala que la especie se alimenta de copépodos, ostrácodos, foraminíferos y restos vegetales, sin embargo no se categoriza su importancia individual (López-López et al. 1991). En otras áreas de distribución geográfica como en la costa de Colombia, se encontró que esta especie consume hasta 32 ítems alimenticios (Arenas-Granados y Acero 1992). Se observó que la dieta de D. rhombeus en el Pom-Atasta incluye grupos tróficos registrados en otros sistemas costeros, encontrándose variaciones porcentuales en la abundancia relativa de las presas, lo cual refleja las adaptaciones que posee esta especie para alimentarse de organismos del micro y meiobentos.
Agradecimientos
A la Universidad Autónoma Metropolitana- Xochimilco por el financiamiento, a través del Proyecto de investigación "Ecología y Evaluación de los Recursos Pesqueros Asociados al Sistema Fluvio-Deltaico Pom- Atasta, Campeche México" adscrito al Laboratorio "Ecología Costera y Pesquerías" del Departamento "El Hombre y su Ambiente". A Hernán Alvarez-Guillén de la Estación "El Carmen" del ICML-UNAM, en Ciudad del Carmen Campeche, por su apoyo en campo, laboratorio y biblioteca.
Resumen
Se estudió el pez D. rhombeus en el sistema fluviodeltaico Pom-Atasta, asociado a la Laguna de Términos Campeche, en un ciclo anual de 1992 a 1993. Esta especie es dominante en el sistema por su abundancia numérica, peso, frecuencia y amplia distribución. Se obtuvieron 745 individuos con un peso total de 2 890.2 g y un intervalo de longitud total de 3.0 a 16.7 cm. La variación anual del coeficiente alométrico b fue de 2.71 a 3.45. El factor de condición varió de 0.711 a 0.934. Fueron obtenidas diferencias significativas (p< 0.05) entre las épocas del año y los hábitats del sistema para el peso, la longitud y el factor de condición K, lo cual refleja la utilización espacio-temporal del sistema por la especie. La población contenida en Pom-Atasta, está constituida principalmente por individuos juveniles, y escasos preadultos, en estadios gonádicos I, II y III, registrándose más hembras que machos. La especie utiliza el sistema como área de crianza, crecimiento y alimentación. Tiene un espectro trófico variado, consume al menos ocho grupos diferentes, siendo su alimento principal materia orgánica no determinada, foraminíferos, ostrácodos y tanaidáceos. Es un consumidor de primer orden. El sistema Pom-Atasta se ubica en una zona de intensa actividad pesquera y petrolera, por lo cual es importante avanzar en el conocimiento de sus recursos pesqueros.
Palabras clave: Diapterus rhombeus, biología, ecología, Pom-Atasta, Golfo de México.
Referencias
Aguirre-León, A., A. Yañez-Arancibia & F. Amezcua- Linares. 1982. Taxonomía, diversidad, distribución y abundancia de las mojarras de la Laguna de Términos, Sur del Golfo de México (Pisces: Gerreidae). An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. 9: 213-250. [ Links ]
Aguirre-León, A. & A. Yáñez-Arancibia. 1986. Las mojarras de la Laguna de Términos, Campeche: Taxonomía, biología, ecología y dinámica trófica (Pisces: Gerreidae). An Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. 13: 369-444. [ Links ]
Aguirre-León, A., S. Díaz-Ruiz. & A. Bernal-Becerra. 1998. Ecología de Peces en Sistemas Fluvio-Deltaicos: Estudio para el Sistema Pom-Atasta en Campeche. Serie Académicos: 28. Ciencias Biológicas y de la Salud. Universidad Autónoma Metropolitana- Xochimilco. México. 64 p. [ Links ]
Aguirre-León A., O. Trejo-Buendía, L.A. Ayala-Pérez, S. Díaz-Ruiz & O.A. Avilés-Alatriste. 1993. Estructura comunitaria del necton en el sistema fluvio-lagunar Pom-Atasta, Campeche, México. V Congreso Latinoamericano sobre Ciencias del Mar. Universidad Autónoma de Baja California Sur. La Paz, Baja California Sur. México. 249 p. [ Links ]
Amezcua-Linares, F. & A. Yañez-Arancibia. 1980. Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El hábitat y estructura de las comunidades de peces. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. 7: 68-118. [ Links ]
Araujo, F.G. & A.C.A. Santos. 1999. Distribution and recruitment of mojarras (Perciformes, Gerreidae) in the continental margin of Sepetiba Bay, Brazil. Bull. Mar. Sci. 65: 431-439. [ Links ]
Arenas-Granados, P. & P.A. Acero. 1992. Organización trófica de las mojarras (Pisces: Gerreidae) de la Ciénega Grande de Santa Marta (Caribe Colombiano). Rev. Biol. Trop. 40: 287-302. [ Links ]
Austin, H.M. 1971. Some aspects of the biology of the rhomboid mojarra Diapterus rhombeus in Puerto Rico. Bull. Mar. Sci. 21: 886-903. [ Links ]
Ayala-Pérez. L.A., A. Aguirre-Léon, O.A. Avilés-Alatriste, M.T. Barreiro-Güemes & J.L. Rojas-Galavíz. 1993. Peces de sistemas fluvio-lagunares, Laguna de Términos, Campeche, pp. 596-608. S. Salazar-Vallejo & N. E. González (eds.). Biodiversidad Marina y Costera de México. Com. Nal. Biodiversidad y CIQRO. México. [ Links ]
Ayala-Pérez. L.A., B.A. Gómez-Montes & J. Ramos Miranda. 2001. Distribución, abundancia y parámetros poblacionales de la mojarra Diapterus rhombeus (Pisces: Gerreidae) en la Laguna de Términos, Campeche, México. Rev. Biol. Trop. 49: 635-642. [ Links ]
Barreiro-Güemes, M.T. & A. Aguirre-León. 1995. Estructura y producción primaria de la comunidad de pastos marinos en el sistema fluvio-lagunar Pom-Atasta, Campeche, México. VI Congreso Latinoamericano de Ciencias del Mar. Universidad Nacional de Mar del Plata. Mar del Plata, Argentina. 217 p. [ Links ]
Cabrera-Díaz, I. 1995. Ecología y dinámica poblacional de las especies de peces dominantes en el sistema fluviolagunar- estuarino Pom-Atasta, Campeche, México. Reporte Final de Servicio Social, Universidad Autónoma Metropolitana-Xochimilco. 61 p. [ Links ]
Carriker, M.R. 1967. Estuaries, pp. 442. In G.H. Lauff (ed.). American Association for the Advancement of Science, Washington D.C. (83): 757 p. [ Links ]
Cervigón, F. 1993. Los peces marinos de Venezuela. Vol. II. Fundación Científica Los Roques. Caracas, Venezuela. 497 p. [ Links ]
De La Lanza Espino, G., P. Ramírez-García, F. Thomas & A.R. Alcántara. 1993. La vegetación de manglar en la Laguna de Términos, Campeche. Evaluación preliminar a través de imágenes Landsat. Hidrobiol. 3: 29-40. [ Links ]
Etchevers, S.L. 1978. Maturity, spawning, growth and ectoparasites of the rhomboid mojarra Diapterus rhombes (Cuvier) (Pisces: Gerreidae) South of Margarita Island, Venezuela. Bull. Mar. Sci. 28: 385-3912. [ Links ]
Gómez-Montes, B.A. 1997. Biología de las especies dominantes Eugerres plumieri y Diapterus rhombeus en el sistema fluvio-deltaico Pom-Atasta, Campeche, México. Informe de Servicio Social. Universidad Autónoma Metropolitana-Xochimilco. 41 p. [ Links ]
Gónzalez Cabellos, L.W. 1985. Relación longitud-peso y factor de condición de la caitipa, Diapterus rhombeus (Cuvier, 1829) (Pisces: Gerreidae) del sur-oeste de la Isla Margarita, Venezuela. Bol. Inst. Oceanogr. Univ. de Oriente 24: 15-22. [ Links ]
Gutierrez-Estrada, M., V.M. Malpica-Cruz & J. Martínez- Reyes. 1982. Geomorfología y sedimentos recientes del sistema Lagunar Atasta-Pom. Campeche, México. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. 9: 89-100 [ Links ]
Hernández-Sánchez, A.R. 1994. Dinámica poblacional de Diapterus rhombeus (Cuvier)y Eugeres plumieri (Cuvier) (Piscies: Gerreidae), en el sistema Chumpan-Blachacah, sur del Golfo de México. Informe de Servicio Social. Universidad Autónoma Metropolitana-Xochimilco. 22 p. [ Links ]
López-López, E., M. Salgado-Mejía & A. Guzmán-del Proo. 1991. Un análisis estacional de la ictiofauna de la Laguna de Tampamachoco, Veracruz, y sus hábitos alimentarios. An. Esc. Nal. Cienc. Biol. México. 34: 81-107. [ Links ]
Matheson, R.E, Jr. & R.G. Gilmore Jr. 1995. Mojarras (Pisces: Gerreidae) of the Indian River Lagoon. Florida. Bull. Mar. Sci. 57: 281-285. [ Links ]
Molina-Ocampo, R. 1993. Hábitos alimenticios de peces pelágicos menores de importancia comercial del Golfo de California, México. Tesis de Maestría. Instituto Tecnológico de Estudios Superiores de Monterrey, México. 107 p. [ Links ]
Montgomery, D. 1984. Design and análisis of experimental. Williams. New York. 538 p. [ Links ]
Nitsh, S.L. 1992. Ictioecología del sistema fluvio-lagunar Candelaria-Panlau asociado a la Laguna de Términos, Campeche (Sur del Golfo de México). Tesis de Licenciatura. Escuela Nacional de Estudios Profesionales-Iztacala. Universidad Nacional Autónoma de México. 180 p. [ Links ]
Nikolsky, G.V. 1963. The Ecology of Fishes. Academic. Londres. 352 p. [ Links ]
Pauly, D. 1984. Fish Populations Dynamics in Tropical Waters. A Manual for Use with Programmable Calculators. ICLARM Studies and Reviews 8. International Center for Living Aquatic Resources Management. Manila, Filipinas. 325 p. [ Links ]
Reséndez-Medina, A. 1981. Estudio de los peces de la Laguna de Términos, Campeche, México. Biotica. 6: 345-423. [ Links ]
Reséndez-Medina, A. & A. Kobelkowsky-Díaz. 1991. Ictiofauna de los sistemas lagunares costeros del Golfo de México. Univ. y Cienc. 8: 91-110. [ Links ]
Ricker, W.E. 1975. Computation and interpretation of biological statistic fish populations. Department of Environment Fisheries and Marine Service. Bull. Fish. Res. Bd. Canada. (191): 382 p. [ Links ]
Rossenblum, M., J. Pudney, & I. Callard. 1987. Gonadal morphology enzyme histochemistry and plasma steroid levels during the annual reproductive cycle of male and female brown bullhead catfish, Ictalurus nebulosus Lesueur. J. Fish. Biol. 31: 325-341. [ Links ]
Vargas-Maldonado, I., A. Yáñez-Arancibia & F. Amescua- Linares. 1981. Ecología y estructura de las comunidades de peces en áreas de Rhizophora mangle y Thalassia testudinum de la Isla del Carmen, Laguna de Términos sur del Golfo de México. An. Inst. Cienc. Mar Limnol. Univ. Autón. México 8: 241-266. [ Links ]
Vera-Herrera, F., J.L. Rojas-Galavíz, C. Fuentes Yaco, L.A. Ayala-Pérez, H. Alvarez-Guillén & C. Coronado- Molina. 1988. Descripción ecológica del sistema fluvio- lagunar-deltáico del río Palizada, p. 51-88. In A.
Yáñez-Arancibia & J.W. Day, Jr (eds.). Ecología de los Ecosistemas Costeros en el sur del Golfo de México: La Región de la Laguna de Términos. Instituto de Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México. Coastal Ecology Institute, Louisiana State University. México.
Yáñez-Arancibia, A. & P. Sánchez-Gil. 1986. Los peces demersales de la plataforma continental del sur del Golfo de México. Vol. 1. Caracterización del ecosistema y ecología de las especies, poblaciones y comunidades. Inst. Cienc. del Mar Limnol. Univ. Nal. Autón. México. Pub. Esp. 9: 1-230. [ Links ]
Yáñez-Arancibia, A. & A., Aguirre-León. 1988. Pesquerías en la región de la Laguna de Términos, p. 431-452. In Yáñez-Arancibia, A. & J.W. Day, Jr. (eds.). Ecología de los ecosistemas costeros en el sur del Golfo de México: La región de la Laguna de Términos. Instituto de Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México. Coastal Ecology Institute, Louisiana State University. México. [ Links ]
Yáñez-Arancibia, A. & J.W. Day Jr. 1988. Ecological characterization of Terminos Lagoon, a tropical lagoonestuarine system in the southern Gulf of Mexico, p. 431-440. In Yáñez-Arancibia, A. & J.W. Day, Jr. (eds.). Ecología de los ecosistemas costeros en el sur del Golfo de México: La región de la Laguna de Términos. Instituto de Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México. Coastal Ecology Institute, Louisiana State University. México. [ Links ]
Yañez-Arancibia A., A.L. Lara-Domínguez, A. Aguirre- Léon & S. Díaz-Ruiz. 1986. Feeding ecology of tropical estuarine fishes in relation to recruitement processes, p. 73-93. In Yañez-Arancibia, A. & D. Pauly (eds.). IOC/FAO Workshop on Recruitment in Tropical Coastal Demersal Communities. Ciudad del Carmen, Campeche. México. IOC/FAO Unesco. Workshop Rep. No. 44 (Suppl.). [ Links ]

