










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 n.1 San José Mar. 2006
Distribution of bacteria and fungi in the earthworm Libyodrillus violaceous (Annelida: Oligochaeta), a native earthworm from Nigeria
*A. B. Idowu, M. O. Edema & A. O. Adeyi
Department of Biological Sciences, College of Natural Sciences, University of Agriculture, Abeokuta, Nigeria.
*Corresponding author: tomiwo2@yahoo.com
Received 16-II-2005. Corrected 05-VIII-2005. Accepted 01-IX-2005.
Abstract: Earthworms are soil invertebrates that play a key role in recycling organic matter in soils.In Nigeria, earthworms include Libyodrillus violaceous. Aerobic and anaerobic bacterial counts, as well as fungal counts of viable microorganisms in soils and gut sections, were made on twenty L. violaceous collected from different sites on the campus of the University of Agriculture, Abeokuta, Nigeria. The samples were collected between April and November, 2002. Numbers of microorganisms were higher in castings and gut sections than in uningested soil samples. The guts and their contents also had higher moisture and total nitrogen contents than the uningested soils. Bacteria and fungi isolated from the samples were identified by standard microbiological procedures on the bases of their morphological and biochemical characteristics. Isolated bacteria were identified as Staphylococcus, Bacillus spp., Pseudomonas aeruginosa, Streptococcus mutans, Clostridium, Spirocheata spp., Azotobacter spp., Micrococcus lylae, Acinetobacter spp., Halobacterium for bacteria. Yeast isolates were identified as Candida spp., Zygosaccharomyces spp., Pichia spp., and Saccharomyces spp while molds were identified as, Aspergillus spp., Pytium spp., Penicillium spp., Fusarium spp and Rhizopus spp. Of the five locations examined, the refuse dump area had the highest numbers of both aerobic and anaerobic organisms, followed by the arboretum while the cultivated land area recorded the lowest counts. The higher numbers of microorganisms observed in the gut sections and casts of the earthworms examined in this work reinforce the general concept that the gut and casts of earthworms show higher microbial diversity and activity than the surrounding soil. Rev. Biol. Trop. 54 (1): 49-58. Epub 2006 Mar 31.
Key words: Total microbial counts, Bacteria, Fungi, Earthworm casts, Gut contents.
Earthworms are soil invertebrates that play a key role in recycling organic matter in soils (Satchell 1967). They are also ecosystem engineers as they actively redesign the physical structure of the soil environment by their activities of ingesting litter and soil particles, depositing casts on the soil surface and translocating soil particles while their gut, within which many changes take place have been described as natural bioreactors. The activities of earthworms in soils have been shown to have profound impart on the soil ecosystem functioning as well as on the types and numbers of micro-flora and micro-fauna (Pederson et al. 1993). Earthworms ingest soil microorganisms along with organic residues from the soil and during passage through the earthworms intestinal tract, their population may increase. Earthworm casts have been reported to be much more microbiologically active and richer in micro-flora than their surrounding un-ingested soils (Scheu 1987, Daniel and Anderson 1992). It has also been reported that while earthworms use organic matter as their nutrient source, the microorganisms ingested along with these nutrient sources actually elaborate the enzymes that make the nutrients available for the worms use (Edward et al. 1972, Lee 1985). It is likely that the ingested microbial populations play a key role in earthworm nutrition by helping in the breakdown of organic matter, particularly the components that the earthworms cannot utilize in their natural state (Hornor et al. 1981). Several workers have compared the mineral composition of the earthworms food substrate with those of its excreted casts and reported that the earthworm s activities significantly increase the nitrogen mineralization in the soil (Sharon et al. 2002 Edwards 1972). Daniel and Anderson (1992) reported higher oxygen uptake and carbon dioxide production in casts than in surrounding soil. Earthworms can also affect the types of microorganisms in the soil as was shown by Sandra et al. (1996) who reported the suppression of pathogenic fungi during passage through the gut of some earthworm species. Despite these reports, it has remained unclear whether the differences observed in microbial populations were a result of growth during gut transit or as a result of selective consumption of organic matter richer in microorganisms or due to differences in culturability of microorganisms among other possible factors. In addition, previous studies have emphasized comparisons between casts and surrounding soil without looking at the differences in earthworm species and locations. This work was therefore aimed at a comparative examination of microbial populations in the gut sections and casts of Libyodrillus violaceous (Eudrilidae: Oligochaeta), a native earthworm species commonly found in soils from Abeokuta, Nigeria and environment (Segun and Owa 1990).
Materials and methods
Study site and sampling locations: The study was conducted at the University of Agriculture, Abeokuta, Nigeria (7º10 N, 3º 2 W) at an elevation of about 500 m above sea level in the outskirts of the town in Alabata village. Samples were taken in the arboretum, from cultivated land around the University main gate and a refuse dump area (composed mainly of agricultural waste)located in the premises. Soil texture was loamy to sandy loam while soil pH was between 6.0 and 6.4. These three locations were chosen as sampling sites because they recorded high organic matter contents (3.72% to 3.9%) and were rich in soil fauna in a previous study by the authors (Idowu et al .2003).Their relatively high organic matter contents and the presence of soil fauna (protozoans, nematodes and earthworms)could be indicative of the presence and activity of microorganisms.
Experimental design and sampling procedure: Soil sampling and specimen collection were done between April and November 2002. A day before collection of earthworm and casts, the soil was cleared of all observable casts. Cast samples collected thereafter were stored at ambient temperature (27ºC) for up to 2 weeks. Twenty earthworms belonging to Libyodrillus violaceous species were collected from each of the three locations examined. The samples /specimens were taken from a depth of about 15 cm. Fresh casts lying on the soil surfaces were also collected. All samples were collected in triplicate from each research location using sterilized soil diggers and in sterile polythene bags. Soil pH values ranged from 6.0 to 6.8. There were no significant differences in pH values obtained from all sampling sites. Soil moisture contents ranged from 21.4% to 69.3%. Earthworm specimens from each location were collected along with some surrounding soils. The soil samples were moistened with sterile water in labeled specimen bowls; the earthworms were placed on the soil samples before covering the bowls with wire mesh. They were then stored at room temperature in secluded parts of the laboratory to avoid disturbance. The earthworms and casts produced were analyzed after 2 weeks laboratory storage.
Dissection of earthworms: Each specimen to be dissected was washed in sterile distilled water, placed across the second, third and fourth fingers of the left hand (gloved) with the anterior end pointing forward. The fine edge of a flamed pair of dissecting scissors was inserted into the ventral surface at the region of the clitella and with the body wall slightly raised up with the scissors; an incision was made longitudinally along the earthworm. Sterile dissecting pins were used to hold the earthworm down on a board, stretching out the body wall to expose the internal structures. The gut was then freed from surrounding blood vessels and nephridia with a flamed forceps and separated into three sections: foregut, mid gut and hindgut. The gut sections were washed in sterile distilled water to free their contents before being suspended in other bottles containing clean sterile distilled water.
Abiotic measurements: Replicate soil and cast samples were mixed thoroughly in distilled water to supernatant samples in the ratio 1:5 (w/v)soil or cast: water. The soil or cast: water suspensions were used to determine soil pH by a combined glass electrode and a pH meter (Mettler-Toledo, Essex M3509 Type 340). Moisture contents of soil and cast samples were estimated as percentages in triplicates by drying the samples at 105 o C for 24 hr in a hot air oven (A.O.A.C.1990). Soil temperatures were recorded with a soil thermometer dipped into the soil at the depth of sample collection (15 cm).Total soil nitrogen was determined by the macro-Kjeldahl method (Jackson 1962, Black 1965).
Biological analyses: Replicate portions of ratio 1:5 (w/v)sample: sterile distilled water was made for soils, casts and contents of the three gut regions of the earthworm specimens. The preparations were homogenized. 1ml subsample was taken from each of the above preparations and homogenized in 9ml sterile distilled water to make 10-fold serial dilutions of up to 108. A 0.1ml aliquot of the seventh (107) and eighth (108) dilutions were each inoculated in duplicates by the pour plate technique on Sabouraud Dextrose Agar medium supplemented with 0.01%(w/v)Streptomycin Sulphate for isolation and enumeration of fungi, Nutrient Agar media for isolation of bacteria and Plate Count Agar for total viable counts. Fungal plates were incubated at 25ºC for 5 days while bacterial plates were incubated at 30ºC for 3 days. One set each of bacterial plates were incubated under anaerobic conditions in anaerobic jars using Oxoid gas generating kit (Oxoid, U.K.). At intervals, colony-forming units were determined by making total counts on the incubated plates using a colony counter. Representative colonies emerging from the plates were grouped according to their cultural characteristics, purified by repeated sub-culturing and maintained on appropriate agar slants as stock cultures. The bacterial isolates were tested for Gram reaction (Claus 1992), morphology, motility, catalase and oxidase reactions, citrate utilization, coagulase production, starch hydrolysis and sugar fermentation (Harrigan and McCance 1976, Seeley and Van Demark 1972). The isolates were then identified with reference to Sneath et al. (1986). Yeast isolates were examined for assimilation of sugars and citrate, pseudomycelium formation and urease production before being identified according to Kreger van Rig (1984) and Barnett et al.(1990). Microscopic examination of young, actively growing moulds was on the basis of structures bearing spores and on the spores themselves; presence or absence of septation, rhizoid or other tissues. They were then identified by their micro-morphology as well as the color and micro-morphology of their sporulating structures and conidia according to Barnett and Hunter (1972) and Onions et al.(1981).
Statistical analysis: Data were analyzed with SPSS 10.0 for windows software. One-way ANOVA was used to test for differences in numbers of micro-flora from different locations, soils and casts. Pearsons correlation coefficient was used to test the relationship between micro-flora counts in different locations.
Results
Earthworm casts had the lowest moisture content values, followed by soil samples and gut contents with the gut sections having the highest moisture content values (Table 1). Total soil nitrogen concentration ranged from 0.51 to 0.88% with the lowest values obtained for the soils while the highest values were obtained for the casts. The total nitrogen values were higher in the fresh casts than in the casts obtained after two weeks. Table 1 also shows the total microbial counts in undigested soil samples, casts, gut sections as well as gut contents of the earthworm specimens collected from the three locations examined. Counts were in the order of 104. Aerobic counts ranged from 0.5 cfu/ml to 4.8 cfu/ml, anaerobic counts ranged from 0.7 cfu/ml to 3.9 cfu/ml while fungal counts ranged from 1.4 cfu/ml to 3.8 cfu/ml. Aerobic plate counts increased from the fore gut section to the mid and hind gut sections with the gut contents recording the highest counts. Soil samples had the next highest aerobic counts followed by casts. Counts were higher in the fresh casts than in the old casts. Generally, both the gut sections and contents of specimens recorded the highest anaerobic plate counts followed by fresh casts and un-ingested soil while casts produced after two weeks recorded the lowest anaerobic counts. Significant differences were observed in fungal counts from different locations, with the samples from arboretum recording the highest numbers while the refuse dump area had the lowest. With respect to total microbial load in the various locations investigated, the refuse dump area had the highest counts followed by arboretum while the farmland area had the least counts. Casts obtained after two weeks had lower micro-flora counts than fresh casts. Micro-flora were found to increase in numbers during gut passage in all worms collected from the different soil samples. Fifteen bacterial species belonging to 10 genera and 4 yeast species were isolated and characterized while 5 mold species were observed and identified. Bacterial isolates consisted of a variety of both gram-positive and gram-negative rods and cocci. A combination of cultural, morphological and biochemical characteristics were used in identifying bacterial isolates as Staphylococcus aureus (gram, catalase, coagulase and citrate positive, oxidase negative cocci occurring in bunches) Bacillus spp., (gram, catalase, oxidase and citrate positive, coagulase negative rods) Pseudomonas aeruginosa (gram, citrate and coagulase negative, catalase and oxidase positive rods), Streptococcus mutans (gram and citrate positive, catalase, oxidase and coagulase negative cocci in chains) Clostridium spp. (gram, oxidase and citrate positive, coagulase negative rods), Spirocheata spp. (gram, citrate and coagulase negative, catalase and oxidase positive rods), Azotobacter spp. (gram and coagulase negative, catalase and oxidase positive rods), Micrococcus lylae (gram, oxidase and citrate positive, catalase negative cocci), Acinetobacter spp. (catalase positive, gram, oxidase, citrte and coagulase negative rods), Halobacterium spp. (gram and coagulase negative, catalase, oxidase and citrate positive cocci). Yeast isolates were identified as Candida spp., Zygosaccharomyces spp., Pichia spp., Saccharomyces spp. while molds were identified as, Aspergillus spp., Pytium spp., Penicillium spp., Fusarium spp., Rhizopus spp. All yeast isolates reproduced vegetatively by budding and were able to grow in glucose and starch (except Pichia spp.), none was positive for xylose fermentation and none grew in arabinose, methanol or nitrate. Only Zygosaccharomyces spp. was able to utilize fructose.
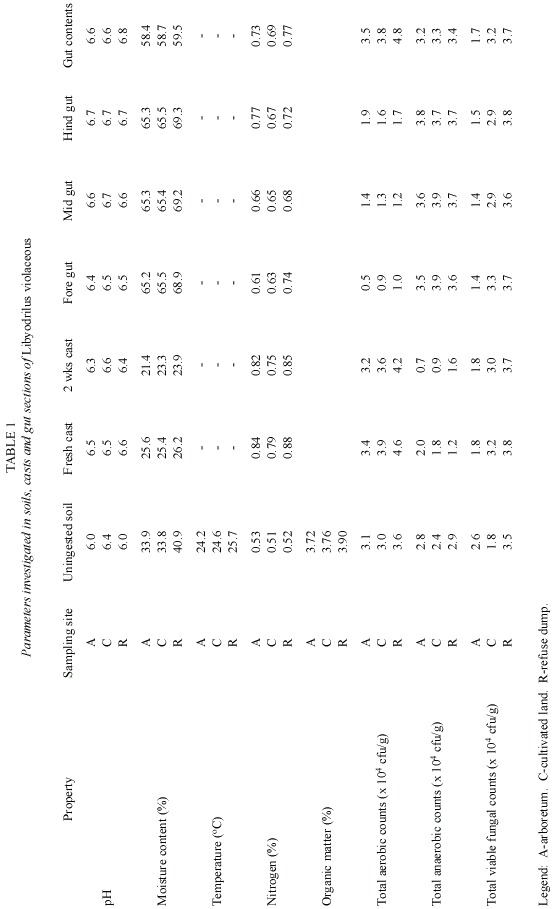
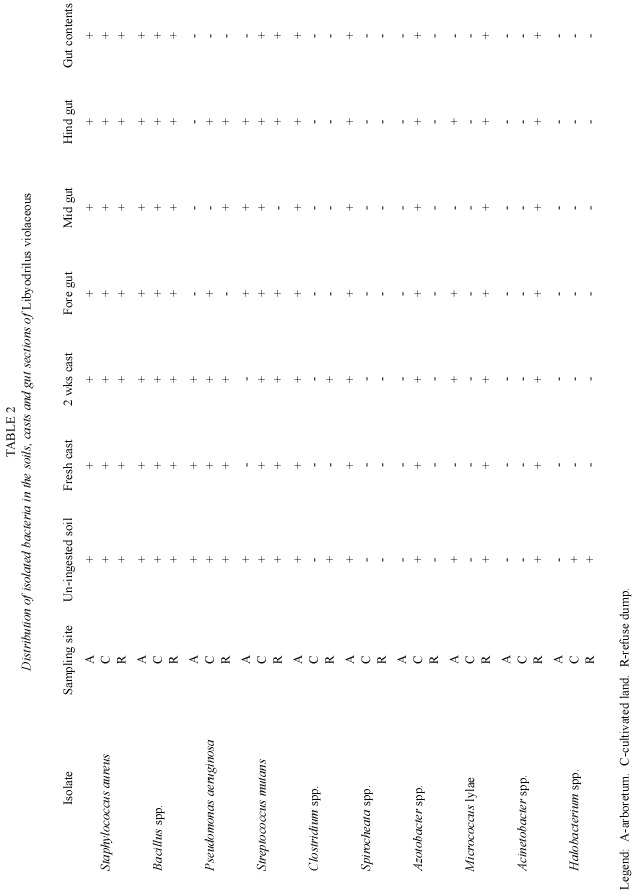
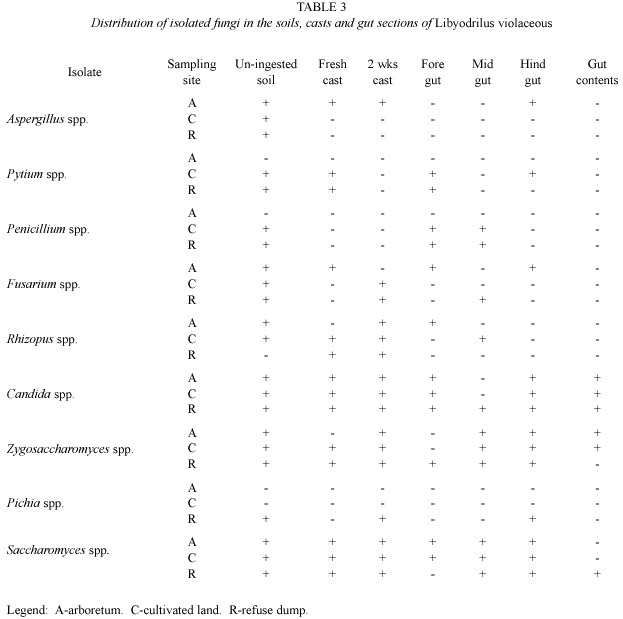
Tables 2 and 3 show the distribution of isolated micro-flora in the soils, casts, gut sections and gut contents of Libyodrilus violaceous collected from the three different locations sampled. Among the bacteria, Staphylococcus aureus and Bacillus spp. were isolated from all the locations and in all samples investigated. Spirocheata spp. was isolated only in samples from the arboretum while Azotobacter was isolated only from cultivated land samples. Micrococcus and Acinetobacter were isolated from all samples from the refuse dump area. Distribution was more varied for the fungal isolates. However, Pythium spp.did not occur at all in samples from the arboretum while Pichia spp.was isolated only from the soil, hindgut and two weeks cast of samples from the refuse dump area.
Discussion
The results of total aerobic and anaerobic counts of micro-flora obtained from soils, earthworm castings, gut sections and gut contents of earthworms indicated that total numbers of microorganisms were higher in castings and gut sections than in the un-ingested soil samples. Daniel and Anderson (1992) in a related study also reported increase of over 100% in the total plate counts of total Actinomycetes and bacteria when samples of the gut were compared with surrounding soil samples. The guts and their contents also had higher moisture and total nitrogen contents than the soils. Counts were higher in fresh earthworm casts than in the casts collected after two weeks. Earthworms have been shown to greatly influence the chemical, physical and microbiological properties of the soils they inhabit (Tiwari 1993). The findings of the present study thus confirm the concept that the earthworm gut might be a specialized microhabitat of enhanced microbial activities in soils (Karsten and Drake 1995). Lee (1985) suggested that the increase in microbial population in the gut of earthworms might be due to the utilization of additional nutrients available as a result of enzymatic breakdown of ingested materials. In the present study, the high nitrogen and water contents recorded in the earthworms guts may also have contributed to this by stimulating microbial activity. Brown (1995) and Winding (1997) were of the opinion that selective feeding of earthworms on materials with higher microbial counts accounted for the population increase in casts produced. This is doubtful in the present study as the microbial flora of undigested soil samples was less when compared with those obtained for the earthworm gut and casts. Results of earlier studies have demonstrated that the feeding habits of earthworms allow for the ingestion of various kinds of organic matter; including dead plant materials and soil micro-organisms (bacteria, yeasts, protozoans, etc.) which are rich in nitrogen content (Worm facts 2001). This feeding habit has profound impact on the soil ecosystem functioning by affecting the availability of organic detritus that serve as food for soil microorganism.(Savin et al. 2000). Pederson et al (1993) stated that the passage of soil detritus and microorganisms through the gut of earthworms have significant, impact on microbial population as recorded in the present study. This is because the gut of earthworms has been described by Worm Digest (1994) as a natural bioreactor which increases the density of beneficial microbes in materials that are excreted. Ten genera of bacteria, five molds and four yeasts were identified from the soils and the gut sections of the earthworms collected.
The great similarity observed in the types of micro-organisms identified in soil samples and those identified in various gut sections of earthworm implies that earthworm ingest with soil and detritus, resident microorganisms which might be of dietary importance to the earthworms without any bias (Savin et al. 2000). It has also been reported that earthworms use organic matter as a source of nutrition but depend on micro-organisms such as bacteria, yeast and fungi for nutrients which are extracted from the large quantities of micro-organisms ingested with materials as they pass through the worms gut for the digestion of food materials especially those that the earthworm cannot digest by itself (Worm Facts 2001). Edward et al. (1972), further stated that earthworm derive their nutrition from organic matter in the form of plant materials, living protozoa, rotifers, nematodes, bacteria, fungi and other micro-organisms, and decomposing remains of large and small animals most of these are extracted form the large quantities of soil that pass through the gut.
The microbial counts of earthworms residing in the refuse dump area were the highest of the sampling locations. This is probably due to the high rate of decomposition of organic matter resulting in more available nutrients for microbial growth and reproduction. Earthworms collected from the cultivated farmland recorded the least counts, probably because of the intensive use of the land for farming resulting in constant turning of soil. The increase in the counts of bacteria and yeasts along the gut of all earthworms analyzed, especially in those of the refuse dump area and arboretum, suggests the growth of microbial population in the gut probably due to increase in availability of nutrients in the gut. This could have been as a result of digestion of ingested materials either by microbial enzymes produced within the gut or by the gut wall of these earthworms. This line of thought agrees with the report of Worm digest (1994) that the earthworms gut is a natural bioreactor, which increases the microbial densities in the gut and in excreted materials. The refuse dump area on the University campus is made up of mostly agricultural wastes that are rich in organic matter, which earthworms and micro-organisms can make use of for their growth and multiplication. The arboretum recorded the next highest numbers of micro-flora. This area is usually shaded and characterized by cool, undisturbed soil profile with relatively low temperature, a combination that is ideal as a habitat for earthworms and microorganisms alike. On the other hand, the cultivated land area is open, frequently disturbed by human activities and generally not a very favorable environment for soil flora and fauna hence the lower numbers of microorganisms recorded from this sampling location in the present study was not totally surprising. The excessive use of chemical fertilizers characteristic of modern farming could also have contributed to the low microbial loads observed in this location. Most changes in agricultural technology have ecological effects on soil organisms that can affect higher plants and animals, including man.
The findings of the present study reinforce the general concept that the gut and casts of earthworms tend to be much more microbiologically active than the surrounding soil. Enhancing the growth of these soil organisms can serve as a basis for the development of living soils by optimizing the potentials of the beneficial biotic populations identified in this work.
References
AOAC. 1990. Official Methods of Analysis, Association of Official Analytical Chemists (AOAC), Washington D.C. [ Links ]
Barnett, H. L. & B. B. Hunter. 1972. Illustrated genera of imperfect fungi. APS, Saint Paul, Minnesota, USA. 218 pp. [ Links ]
Barnett, J. A., R. W. Payne & D. Yarrow. 1990. Yeasts: Characteristics and Identification Cambridge Universit, London. 1002 p. [ Links ]
Black, C. A. (ed.). 1965. Methods of soils analysis. Agronomy No. 9 part 2. Amer. Soc. Agronomy, Madison, Wisconsin, USA. [ Links ]
Brown, G. G. 1995. How do earthworms affect microflora and fauna community diversity? Plant Soil 170:209- 231. [ Links ]
Claus, D. 1992. A standardized gram-staining procedure. Wor. J. Microbiol. Biotechnol. 8: 451-452. [ Links ]
Daniel, O. & J. M. Anderson. 1992 Microbial biomass and activity in contrasting soil materials after passage through the gut of earthworms Lumbricus rubellus . (Hoffmeister) Biol. Fertil Soils 24:465-470. [ Links ]
Edwards, C. A. & J. R. Lofty. 1972. Biology of Earthworms. Chapman and Hall, London. p. 80-81. [ Links ]
Harrigan, W. F. & M. E. McCance. 1976. Laboratory methods in food and diary microbiology. Academic, London. 452 p. [ Links ]
Hornor, S. G. & M. J. Mitchell. 1981. Effect of earthworm E. Felida (Oligocheata)on fluxes of volatile Carbon and Sulfur compounds from sewage sludge. Soil Biol. Biochem. 13: 347-372. [ Links ]
Idowu, A. B., M. O. Edema & A. Z. Kolawole. "2003" (2005). A comparative analysis of fauna populations in the soil of University of Agriculture, Abeokuta, Nigeria. ASSET 2.(2):in press. [ Links ]
Jackson, M. L. 1962 Soil chemical analysis. Prentice Hall, New York, USA. 109 p. [ Links ]
Kreger, van Rig N. J. W.(ed.). 1984. The yeasts-a taxonomic study Amsterdam, Elsevier Science, Amsterdam. 220 p. [ Links ]
Karsten, G. R. & H. L. Drake 1995. Comparative assessment of the aerobic and anaerobic microfloras of earthworm and forest soils Appl. Environ. Microbiol 61: (3) 1039-1044. [ Links ]
Lee, K. E. 1985. Earthworms: Their Ecology and relationship with soil and land use. Academic, Sydney, Australia. 441 p. [ Links ]
Onions, A. H. S., D. Allsopp & H. O. W. Eggins. 1981 Smith s Introduction to Industrial Mycology, Edward Arnold, London. 398 p. [ Links ]
Perdesen J. C. & N. B. Hendriksen. 1993. Effects of passage through the intestinal tract of detritivore earthworms (Lumbricus spp) on the number of selected gram-negative and total bacteria. Biol. Fertil. Soils. 16: 227-232. [ Links ]
Sandra, A. M., G. P. Trenor & M. John. 1996. Fate of some fungal spores associated with wheat straw decomposition on passage through the guts of Lumbrious terrestries and Aponrechoclea longa. Soil Biol. Biochem. 28: 533-537. [ Links ]
Satchell, J. E. 1967. Lumbricidae, p. 259-322. In A. Burges & F. Raw (eds.).Soil biology, Academic, London. [ Links ]
Savin, C. M. & A. A. Jose. 2000. Burrowing a better future: Earthworm and the Soil. RINHEWS 7(1):2-3. [ Links ]
Scheu, S. 1987. Microbial activity and nutrient dynamics in earthworm cast (Lumbricidae). Biol. Fertil. Soils 5: 230-234. [ Links ]
Seeley, H. W.,(Jr.) & P. J. Van Demark. 1972. Microbes in action-a laboratory manual of Microbiology. Freeman, San Francisco. 361 p. [ Links ]
Segun, A. O. & S. O. Owa. 1990. Two new Keffia species (Eudrilinae:Oligochaeta) from Nigeria. J. Nat. Hist. 24: 1507-1515. [ Links ]
Sharon, L. L. & F. H. Paul. 2002. Interaction of the earthworm Diptocardia mississippiensis with microbial and nutrient dynamics in a subtropical spodosol. Soil Biol. Biochem. 33: 1411-1417. [ Links ]
Sneath, P. H. A., N. S. Mair, M. E. Sharpe & J. G. Holt (ed.). 1986. Bergeys manual of systematic bacteriology. Vol. 2, Williams and Wilkins, Baltimore. Ohio, USA. [ Links ]
Tiwari, S. C. & R. R. Mishra. 1993.F ungal abundance and diversity on earthworm casts and in undigested soil. Biol. Fertil. Soils 16:131-134. [ Links ]
Winding, A., R. Ronn. & N. B. Hendrisken. 1997. Bacteria and protozoa in soil microhabitat as affected by earthworms. Biol. Fertil. Soils 24:133-140. [ Links ]
Worm Facts. 2001. Biology of worms. Department of Public Works, Environmental Programs Division. Los Angeles County, Los Angeles, California, USA. p.1-3. [ Links ]
Internet reference
Worm Digest Magazine. "1994". Earthworms: The benefits. www.wormdigest.org; downloaded August 5, 2005). [ Links ]

