










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.53 suppl.1 San José May. 2005
Linking larval history to juvenile demography in the bicolor damselfish Stegastes partitus (Perciformes:Pomacentridae)
Richard S.Nemeth
University of New Hampshire,Department of Zoology,Durham,NH,USA 03824.Present Address:Center for Marine and Environmental Studies,University of the Virgin Islands,2 John Brewer s Bay,St.Thomas,US Virgin Islands,00802-9990.Phone:(340)693-1380,Fax:(340)693-1385;rnemeth@uvi.edu
Received 15-I-2004. Corrected 18-VIII-2004. Accepted 29-III-2005.
Abstract:Otolith-based reconstructions of daily larval growth increments were used to examine the effect of variation in larval growth on size and age at settlement and post-settlement growth,survival and habitat preferences of juvenile bicolor damselfish (Stegastes partitus Poey).During August 1992 and 1994,newly settled S. partitus were collected from Montastraea coral heads and Porites rubble piles in Tague Bay,St.Croix,U.S. Virgin Islands (17 °45 N,64 °42 W).Daily lapillar otolith increments from each fish were counted and measured with Optimas image analysis software.S.partitus pelagic larval duration was 23.7 d in 1992 (n =70)and 24.6 d in 1994 (n =38)and larval age at settlement averaged 13.0 mm total length both years.Analysis of daily otolith increments demonstrated that variation in larval growth rates and size and age at settlement had no detectable effect on post-settlement survivorship but that larger larvae showed a preference for Montastraea coral at settlement.Late larval and early juvenile growth rates showed a significant positive relationship indicating that growth patterns established during the planktonic stage can span metamorphosis and continue into the benthic juvenile phase.Larval growth rates during the first two weeks post-hatching also had a strong effect on age to developmental competence (ability to undergo metamorphosis)in both 1992 and 1994 with the fastest growing larvae being 8 d younger and 0.8 mm smaller at settlement than the slowest growing larvae.These differential growth rates in early stage larvae established trajectories toward larval developmental competence and may prove important in biogeographical studies of larval dispersal.
Key words:Larval to juvenile transition,otoliths,metamorphosis,Caribbean,Stegastes partitus .
A pelagic larval phase is a life history characteristic common to nearly all benthic marine fishes (Sale 1980).During the pelagic phase,fish larvae will encounter an array of biological and physical conditions that influence their rate of development,larval duration and survival (Hunter 1981).The metamorphic transition into a juvenile,which is often associated with a shift from a pelagic to benthic or epi-benthic existence,is also critical to successful recruitment to the adult population.It has been suggested from field-based studies of fishes and invertebrates that pelagic larvae with different pre-settlement histories may have greater or lesser abilities to successfully make the transition to the benthic environment (Cowen 1991,McCormick and Molony 1992). Laboratory studies of temperate marine fishes have verified that larval growth rates can have a substantial influence on time to metamorphosis and on early juvenile life (Bertram and Leggett 1994).Undoubtedly,the role of larval history is essential to our understanding of the biogeography and population dynamics of marine fishes.Although laboratory studies have provided a wealth of information on the early life history of fishes,rearing conditions can be markedly different from those experienced by larvae in the natural pelagic environment. Therefore,it is imperative that variation in larval history and its effects on species distribution and population structure be examined using field-based methods which incorporate natural conditions.
This paper examines the effect of variation in early life history parameters on post-settlement growth and survival of juvenile fish using otolith microstructure analysis.Studies analyzing otoliths of temperate marine fish larvae have reported that differential larval growth rates can persist throughout the larval period and suggest that faster growing larvae metamorphose earlier and at a larger size,and have a survival advantage (Rosenburg and Haugen 1982,Meekan and Fortier 1996).Are these same patterns detectable in tropical marine fish and is there a potential selective advantage to differences in larval history?By comparing the growth history of juveniles surviving for a period of time after metamorphosis with individuals at or before settlement one can determine the potential selective advantages to ontogenetic differences in timing and intensity of growth (Meekan and Fortier 1996).With most tropical benthic marine fishes,the transition from larvae to juvenile (pelagic to benthic)typically occur overnight. Since most benthic reef fishes,especially damselfishes (Pomacentridae),are very site attached at settlement (Sale 1980),one can confirm the settlement date of larvae to within a 24 hr period by conducting daily census and collection of newly settled larvae at specific reef locations during the lunar recruitment cycle.
This study tested four specific hypotheses concerning the larval/juvenile transition of bicolor damselfish (Stegastes partitus Poey) by comparing age and growth data derived from lapillar otolith increments:1)Is post-settlement survivorship affected by variation in larval growth or size and age at settlement?, 2)Are size and age at settlement influenced by larval growth histories? 3)Is there a relationship between larval and juvenile growth rates and 4)Does larval size or age influence habitat selection at settlement?
Materials and methods
As with most pomacentrids of the genus Stegastes ,the bicolor damselfish is territorial at settlement and displays strong site fidelity making it ideal for following the fate of individuals after settlement.Settlement of larval S.partitus was monitored between 29 July and 24 August, 1992 on 24 isolated Montastraea annularis coral heads and Porites porites rubble piles on the fore reef of Tague Bay,St.Croix,United States Virgin Islands (17 °45 N,64 °42 W). To examine the effect of conspecific adults on settlement patterns,half of the rubble piles and coral heads were randomly selected and had adults either added or removed to establish conspecific densities of zero (0)or one (1)fish m-2 .On half of the experimental units with and without adults newly settled S.partitus were collected daily using quinaldine and handnets whereas on the other units recruitment was censused daily and the survivors (recruits)were collected at the end of the study.Once collected, each fish was measured to the nearest 0.5 mm and preserved in 70%ETOH before otoliths were extracted.This experiment was repeated between 15 June and 20 August,1994.
Three comparisons were conducted using the daily growth bands and settlement mark visible on each otolith.First,the size and age at settlement of newly settled larvae and surviving recruits were compared to estimate the effect that larval size and age at settlement had on post-settlement survival.This comparison assumed that newly settled larvae collected daily (settlers)provided the full range of larval sizes and ages at settlement whereas the recruits collected at the end of the study represented a subset of all fish that had settled and successfully survived several weeks on the reef. The relationship between otolith radius up to the settlement mark and fish length at settlement was calculated using individuals which had zero or one post-settlement increment (i.e.collected on day of settlement).Size at settlement of the surviving recruits were back-calculated and compared using linear regression techniques.The second test examined the influence that larval growth rates had on larval duration and size at settlement. Growth rates of larvae with planktonic durations within one standard deviation of the mean were compared to fish that had shorter and longer larval durations.Data were ananlyzed by comparing the first four 6-day intervals (days 1-6,7-12,13-18 and 19-24)using analysis of variance to examine temporal differences in larval growth among age classes.Lastly,the relationship between larval and juvenile growth rates was examined.This analysis used the average width of daily otolith growth increments during late larval (last 5 d presettlement) and early juvenile (first 10 d postsettlement) stages of recruits which had survived more than 10 days post-settlement.To minimize the potential temporal effects on variation in larval condition (Kerrigan 1996),these comparisons were conducted on larvae arriving to the reef during a single settlement pulse (around the August new moon in 1992 and 1994).Finally the effect of size and age on habitat preferences at settlement were examined using two-way ANOVAs with substrate (Montastraea coral vs. Porites rubble)and conspecific adults (present or absent)as independent variables.
To examine otolith microstructure,lapillar otoliths were lightly polished with 4000 grit paper then permanently mounted on glass slides with cyanoacrylate glue.The lapillus with the clearest ring structure was viewed on a compound microscope at 400x or 600x magnification using polarized light and immersion oil.The formation of daily otolith increments in juveniles of the bicolor damselfish have been validated by Robertson et al.(1988)and Nemeth (1996).It was assumed that larval increments were formed daily as well.Growth rings were easily recognized and the transition between larval and juvenile stages (settlement) was visible as a distinct mark (Brothers et al. 1976,Victor 1982,Pitcher 1988).This settlement mark separated the wider pre-settlement (larval)increments (mean =8.5 µm ±0.85 s.d.) from the narrower post-settlement (juvenile) increments (mean =4.0 µm ±0.75 s.d.).Using this natural marker,pelagic larval duration was calculated by counting the increments from the settlement mark to the lapillus nucleus on each otolith three times (Victor 1982).If the counts were greater than ±1 day,a fourth count was made and the three closest numbers were averaged and rounded to the nearest whole number.The width of daily otolith increments was measured using BioScan Optimas imaging software.These data provided an estimate of daily growth rates during larval and juvenile periods (Pannella 1971,Victor 1986).
Results
The majority of S.partitus settled on the reef within a week of the new moon in August in both 1992 (93%)and 1994 (75%).This uni-modal pattern of settlement has been documented previously for this species (Robertson et al.1988,1990).Of the recruits that were censused,72%and 82%survived to the end of the study in 1992 and 1994,respectively, and thus provided a reasonable comparison of larval characteristics of post-settlement fish. Analysis of the larval otolith increments indicated that the 1992 cohort was approximately 1 d younger,on average,than the 1994 cohort (Fig.1,Table 1).
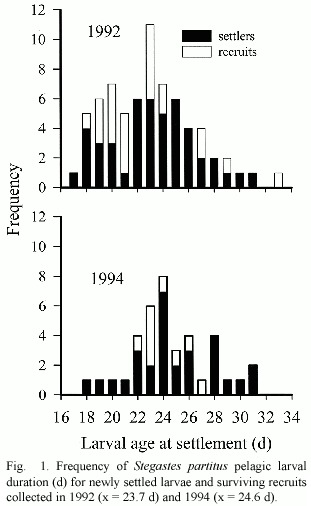
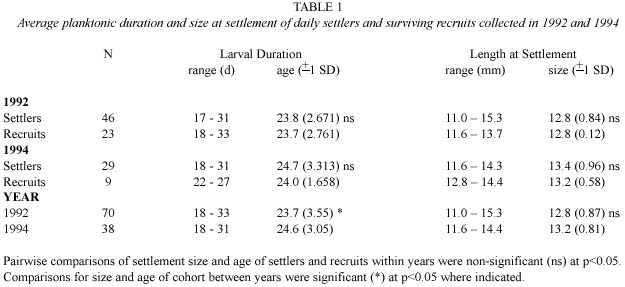
A regression equation was derived from the significant positive relationship between fish length at settlement and the radius of the lapillar otolith at the settlement mark (Fig.2). Analysis of covariance showed that this relationship (i.e.slope of the regression lines)was homogeneous among 1992 and 1994 cohorts (F1,104 =0.41,p =0.52).This equation was then used to back calculate the size at settlement of recruits.Analysis of these results indicated that size at settlement did not differ significantly between settlers and surviving recruits in either year nor among years (Table 1).
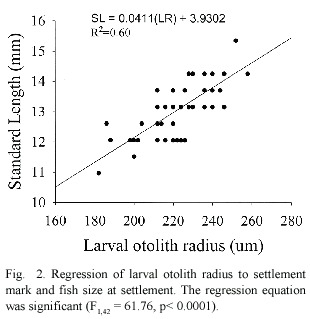
The 1992 and 1994 cohorts were each divided into three age classes representing young,average and old larvae.For all age classes larval otolith growth rates generally increased during the first two weeks post-hatching then declined until settlement (Fig.3). Significant differences in pelagic larval growth rates (using otolith increment width)were found among fish with short,average and long larval durations especially for the 1992 cohort during the early part of their larval life (Fig. 3).Larval growth rates during the second week post-hatching had a highly significant influence on larval duration in 1992 (F1,67 =18.65, P <0.001,R2 =0.47).In 1994 the first week post-hatching had a moderate influence on larval duration (F1,36 =4.76,p<0.05,R2 =0.12). In general S.partitus that had fast growth rates during the first third of their larval life settled at a younger age than fish experiencing slower growth rates.Larval growth rate,however,had no effect on post-settlement survival.
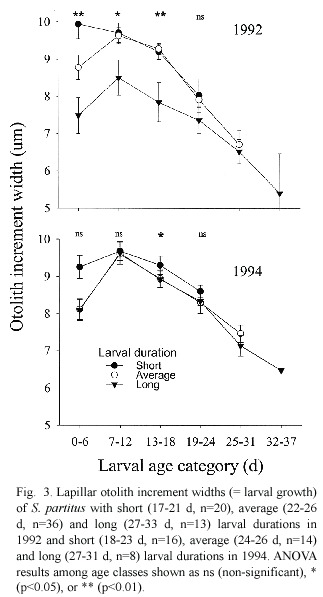
Based on otolith increment width data, somatic growth trajectories from size at hatching (2.1 mm,Robertson et al.1988)to settlement for the three age classes were calculated for 1992 data using the biological intercept method (Campana 1990).The biological intercept equation La =Lc +(Oa – Oc )(Lc -L)(Oc – Oo )-1 calculates fish length as some previous age (La )based on otolith size at age a .The equation incorporates fish size (Lo )and otolith size (Oo )at hatch (biological intercept),and fish length (Lc )and otolith size (Oc )at capture. These trajectories estimated the point where the average size and age at settlement intersected for young,average and old S.partitus larvae (Fig.4).
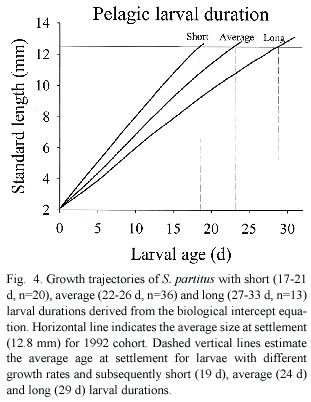
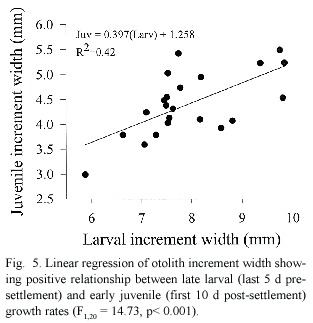
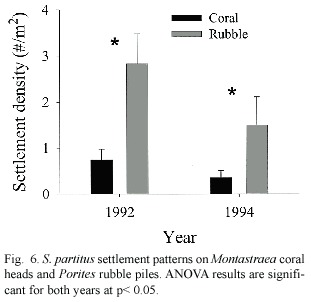
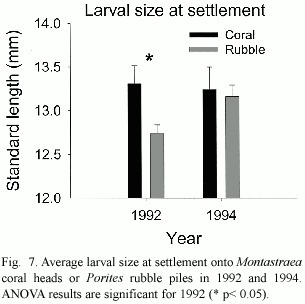
Further analysis of otolith growth patters found that larvae with the fastest larval growth rates prior to settlement continued growing faster than average after settlement (Fig.5). Larval age,however,had no effect on post-settlement growth (F 1,20 =0.23,p =0.63).S. partitus showed a strong preference for rubble substrata in both years (Fig.6)but the presence of adults had no effect on settlement nor recruitment patterns in either year.Although larval age had no effect on settlement patterns,larger larvae showed a preference for Montastraea coral at settlement,an effect that was present only in 1992 (Fig.7).
Discussion
Based on the width of otolith increments, growth rates of larval bicolor damselfish varied considerably from hatching through settlement. In general,otolith increment widths (i.e.larval growth)increased to a maximum 10 to 15 d after hatching then showed a marked decrease beginning around day 18 and continuing until settlement.Although the sample sizes were relatively small,especially for 1994,the initial period of rapid growth in the first two weeks post-hatching was consistent with the larval growth patterns of S.partitus from the San Blas Islands,Panama (Wilson and Meekan 2002).Decreased larval growth as S.partitus larvae approached developmental competence (ability to undergo metamorphosis as inferred from reductions in otolith increment width)was similar to that described for the bluehead wrasse (Thalassoma bifasciatum )(Victor 1986,Searcy and Sponaugle 2000). Slowed growth at competency and the ability to delay metamorphosis with the resultant flexibility in the timing of settlement may be adaptive for reducing risk of mortality by maintaining a smaller size in the plankton (Scheltema 1971, Pechenik et al.1984)while maximizing the chance of return to coastal waters (Jackson and Strathmann 1981,Victor 1986).
Day 17 in the life of larval S.partitus corresponded to the youngest and smallest (11 mm SL)fish collected at settlement suggesting that S.partitus in the Virgin Islands are unable to undergo metamorphosis and settlement prior to this minimum age.Average pelagic larval durations in this study (23.8 d and 24.6 d for 1992 and 1994)were similar to those found by Wellington and Victor (1989)in St.Croix (28.8 d)but were considerably shorter than those found by Robertson et al.(1988)(36.5 d)and Wilson and Meekan (2002)(33.5 d)in Panama. The average size (SL)of newly settled S .partitus collected in 1992 (12.8 mm)and 1994 (13.2),however,was comparable with the studies from St.Croix (12.8 mm,Wellington and Victor 1989)and Panama (13 mm,Robertson et al.1988,12.2 mm,Wilson and Meekan 2002)indicating that size at settlement may be constrained by factors such as predation by reef-based plankton feeders (i.e.if larvae get too big they may be more visible to predators at settlement).
Analysis of otolith microstructure indicated that individuals with shorter larval durations had faster larval growth rates and settled at smaller sizes than fish with longer larval durations and slower larval growth rates.The early life-history patterns found for S.partitus suggest that differential growth rates in early stage larvae establish trajectories toward larval competence and provide a mechanism for determining pelagic larval duration (Houde 1987).These patterns were also reported for T.bifasciatum (Searcy and Sponaugle 2000). The primary difference in larval traits between these two species is that rapid otolith growth occurred 1-14 d post-hatching for S.partitus whereas it occurred 30 to 40 d post-hatching for T.bifasciatum .Leis (1983,1991)found that larvae from pelagic spawned eggs (e.g. Labridae)tend to disperse into waters farther offshore than larvae hatched from demersal eggs (e.g.Pomacentridae).Pelagic spawned eggs most likely disperse into off shore oligotrophic waters where food concentrations may be low relative to nutrient enriched coastal waters (Hernandez-Leon 1991).Demersal eggs hatch immediately into coastal waters and may have access to greater concentrations of food and faster initial growth rates.McCormick and Molony (1992)found that goatfish larvae (Upeneus tragula )that were provided more food,grew faster and settled 10 d earlier than the slowest growing fish (14 d vs.24 d).Slow growth has been shown to increase time to settlement in temperate species as well (Chambers and Leggett 1987).
Variation in growth rates early in larval life may reflect differences in morphological development (Liem 1991),larval feeding behavior (Blaxter 1986),environmental variability in food supply (Kingsford 1990), maternal effects (Chambers and Leggett 1996), or a combination of these factors.Functional morphological studies have shown that larval fishes pass through several critical stages in the development of their feeding apparatus (Otten 1982,Dilling 1989,Liem 1991).The first week post-hatching is associated with large amounts of variation in feeding ability leading to differential rates of growth.For instance,percent feeding success of many species of temperate marine fishes increases dramatically from day 2 (20%-70%success)to day 15 (85%-95% success)post-hatching (Coughlin 1992,review by Blaxter 1986)and may explain the variation in early larval growth of S.partitus .Moreover, water temperature,rainfall and wind components have been identified as important determinants of larval growth of S.partitus (Wilson and Meekan 2002).Unfortunately information on environmental variabilty in St.Croix was not available during this study.
In this study,the fastest growing larvae settled 8 d earlier and were 0.8 mm smaller than the slowest growing larvae.Analysis of these data for daily settlers versus surviving recruits produced no evidence that younger,smaller larvae were at a disadvantage after settlement. These data are supported by a laboratory study that separated the effects of size and age on survival and found no effect of either life history trait on the rate of predation of newly metamorphosed winter flounder (Pleuronectes americanus )(Bertram and Leggett 1994).Although size at settlement of S.partitus larvae had no effect on post-settlement traits,larger S.partitus larvae showed a preference for Montastraea coral over Porites rubble.Although sample sizes were small,the advantage of larger larvae selecting coral substrate over rubble may be related to size-dependent preferences in food and shelter (Nemeth 1997,1998).
Variability in late-larval growth rates appeared to span metamorphosis and continue into the juvenile benthic phase.S.partitus which had elevated larval growth rates near the end of the pelagic period, continued to grow faster than average after settlement and thus compensated for their slightly smaller size at settlement.This is in contrast to T.bifasciatum where variability in early-life history traits did not appear to affect juvenile growth rates (Searcy and Sponaugle 2000).Larvae with faster growth rates prior to settlement most likely reflected better condition at settlement which may have improved foraging success and possibly survivorship (Booth and Hixon 1999). Understanding these relationships between larval and juvenile phases will begin to narrow the gap in our knowledge of the linkages between reproduction and recruitment.
Acknowledgments
This project was funded by NSF grant OCE-9018724 to P.Sale.Additional funding for travel and research was obtained from the American Museum of Natural History s Lerner Grey Fund for Marine Research and University of New Hampshire Center for Marine Biology. The St.Croix Yacht Club and B.Warner of University of California Santa Barbara provided generous logistical support.Special thanks to D.Nemeth,N.Tolimieri,K.Gestring and H.Shaw for diving assistance.This manuscript was greatly improved by comments from P. Sale,D.Breitburg,L.Harris,H.Howell,T.Lee and two anonymous reviewers.
Resumen
Reconstruyendo aumentos diarios de otolitos se compará la variación en crecimiento larval sobre el tamaño y la edad de asentamiento,y el crecimiento post-acentamiento, sobrevivencia y preferencia de hábitat,del pez damisela bicolor (Stegastes partitus Poey)joven.En agosto de 1992 y 1994 se recolectó S.partitus recientemente asentados en colonias de Montastraea y en escombros de Porites en Tague Bay,St.Croix,Islas Vírgenes Estadounidenses (17 °45 N,64 °42 W).A cada pez se le midió el aumento diario en el otolito con el programa de análisis de imágenes Optimas .El estadío de larva pelágica de S.partitus duró 23.7 d en 1992 (n =70)y 24.6 d en 1994 (n =38)y,al asentarse,el largo total promedio en ambos años fue de 13.0 mm.Los otolitos indican que las variaciones en las tasas de crecimiento,tamaño y edad de asentamiento no tienen un efecto detectable en la sobrevivencia post-asentamiento.Sin embargo,las larvas más grandes mostraron preferencia por asentarse en el coral Montastraea .Las tasas de crecimiento larval tardío y juvenil temprano tuvieron una correlación positiva significativa:los patrones de crecimiento establecidos durante el estadío planctónico pasan la metamorfosis y continúan en la fase juvenil béntica.La tasa de crecimiento larval durante las primeras dos semanas después de eclosionar también tiene un efecto sobre la edad a la que concluyeron su desarrollo (momento de la metamorfosis)en 1992 y 1994.Las larvas de crecimiento rápido fueron 8 d más jóvenes y 0.8 mm más pequeñas al asentarse.Este crecimiento diferencial en el estadío larval temprano establece diferencias en la duración del desarrollo larval y puede ser importante en estudios biogeográficos sobre dispersión de larvas.
Key words:Transición de larva a adulto,otolitos,metamorphosis,Caribe,Stegastes partitus .
References
Bertram,D.F.&W.C.Leggett.1994.Predation risk during the early life history periods of fishes:separating the effects of size and age.Mar.Ecol.Prog.Ser.109: 105-114. [ Links ]
Blaxter,J.H.S.1986.Development of organs and behavior of teleost larvae with special reference to feeding and predator avoidance.Trans.Am.Fish.Soc.115:98-114. [ Links ]
Booth,D.J.&M.A.Hixon.1999.Food ration and condition affect early survival of the coral reef damselfish, Stegastes partitus .Oecologia 121:364-368. [ Links ]
Brothers,E.B.,C.P.Mathews &R.Lasker.1976.Daily growth increments in otoliths from larval and adult fishes. Fish. Bull. 74:1-8. [ Links ]
Campana,S.E.1990.How reliable are growth back-calculations based on otoliths? Can. J. Fish. Aquat. Sci. 47:2219-2227. [ Links ]
Chambers,R.C.&W.C.Leggett.1987.Size and age at metamorphosis in marine fishes:an analysis of laboratory-reared winter flounder (Pseudopleuronectes americanus )with a review of variation in other species. Can. J. Fish. Aquat. Sci. 44:1936-1947. [ Links ]
Chambers,R.C.&W.C.Leggett.1996.Maternal influences on variation in egg sizes in temperate marine fishes. Am. Zool. 36:180-196. [ Links ]
Coughlin,D.J.1992.The ontogeny of feeding behavior of larval fishes.Ph.D.Diss.,Boston University, Boston. 170 p. [ Links ]
Cowen,R.K.1991.Variation in the planktonic larval duration of the temperate wrasse Semicossyphus pulcher . Mar. Ecol. Prog. Ser.69:9-15. [ Links ]
Dilling,L.1989.An ontogenetic study of the jaw mechanism and feeding modes in Amphiprion frenatus and A.polymnus .B.Sc.Honor s Thesis.Harvard University,Boston.49 p. [ Links ]
Hernandez-Leon,S.1991.Accumulation of meso-zooplankton in a wake area as a causative mechanism of the "island mass effect ".Mar.Biol.109:141-147. [ Links ]
Houde,E.D.1987.Fish early life dynamics and recruitment variability.Am.Fish.Soc.Symp.2:17-29. [ Links ]
Hunter,J.R.1981.Feeding ecology and predation of marine fish larvae,p.33-79.In R.Lasker (ed.).Marine Fish Larvae.University of Washington,Seattle. [ Links ]
Jackson,G.A.&R.R.Strathmann.1981.Larval mortality from offshore mixing as a link between precompetent and competent periods of development.Am.Nat. 118:16-26. [ Links ]
Kerrigan,B.A.1996.Temporal patterns in size and condition at settlement in two tropical reef fishes (Pomacentridae: Pomacentrus amboinensis and P. nagasakiensis ).Mar.Ecol.Progr.Ser.135:27-41. [ Links ]
Kingsford,M.J.1990.Linear oceanographic features:A focus for research on recruitment processes. Austral. J.Ecol.15:391-401. [ Links ]
Leis,J.M.1983.Coral reef fish larvae (Labridae)in the East Pacific barrier.Copeia 3:826-828. [ Links ]
Leis,J.M.1991.The pelagic stage of reef fishes,p.183- 230.In P.F.Sale (ed.).The ecology of fishes on coral reefs.Academic,San Diego. [ Links ]
Liem,K.F.1991.A functional approach to the development of the head of Teleosts:implications on constructional morphology and constraints,p.231-249.In N.Schmidt-Kittler &K.Vogel (eds.).Constructional Morphology and Evolution.Springer-Verlag,Berlin. [ Links ]
McCormick,M.I.&B.W.Molony.1992.Effects of feeding history on the growth characteristics of a reef fish at settlement.Mar.Biol.114:165-173. [ Links ]
Meeken,M.G.&L.Fortier.1996.Selection for fast growth during the larval life of Atlantic cod Gadus morhua on the Scotian shelf.Mar.Ecol.Prog.Ser. 137:25-37. [ Links ]
Nemeth,R.S.1996.Ecological patterns during ontogeny of the bicolor damselfish,Stegastes partitus Poey.Ph.D. Diss.,Univ.New Hampshire.160 p. [ Links ]
Nemeth,R.S.1997.Spatial patterns of bicolor damselfish populations in Jamaica and St.Croix are determined by similar post-settlement processes.Proc.8 th Int. Coral Reef Symp.1:1017-1022. [ Links ]
Nemeth,R.S.1998.The effect of natural variation in substrate architecture on the survival of juvenile bicolor damselfish. Environ. Biol.Fishes.53:129-141. [ Links ]
Otten,E.1982.The development of a mouth-opening mechanism in a generalized Haplochromis species: H.elegans Trewavas,1933 (Pisces,Cichlidae).Neth. J.Zool.32:31-48. [ Links ]
Pannella,G.1971.Fish otoliths:daily growth layers and periodical patterns.Science 173:1124-1127. [ Links ]
Pechenik,J.A.,R.S.Scheltema &L.S.Eyster.1984. Growth stasis and limited shell calcification in larvae of Cymatium parthenopeum during trans-Atlantic transport.Science 224:1097-1099. [ Links ]
Pitcher,C.R.1988.Validation of a technique for reconstructing daily patterns in the recruitment of coral reef damselfish. Coral Reefs 7:105-111. [ Links ]
Robertson,D.R.,D.G.Green &B.C.Victor.1988.Temporal coupling of production and recruitment of larvae of a Caribbean reef fish.Ecology 69:370-381. [ Links ]
Robertson,D.R.,C.W.Petersen &J.D.Brawn.1990.Lunar reproductive cycles of benthic-brooding reef fishes: reflections of larval biology or adult biology?Ecol. Monogr.60:311-329. [ Links ]
Sale,P.F.1980.The ecology of fishes on coral reefs. Oceanogr.Mar.Biol.Ann.Rev.18:367-421. [ Links ]
Scheltema,R.S.1971.Larval dispersal as a means of genetic exchange between geographically separated populations of shallow-water benthic marine gastropods.Biol.Bull.140:284-322. [ Links ]
Searcy,S.P.&S.Sponaugle.2000.Variable larval growth in a coral reef fish.Mar.Ecol.Prog.Ser.206:213-226. [ Links ]
Victor,B.C.1982.Daily otolith increments and recruitment in two coral-reef wrasses,Thalassoma bifasciatum and Halichoeres bivittatus .Mar.Biol.71:203-208. [ Links ]
Victor,B.C.1986.Delayed metamorphosis with reduced larval growth in a coral reef fish (Thalassoma bifasciatum ).Can.J.Fish.Aquat.Sci.43:1208-1213. [ Links ]
Wellington,G.M.&B.C.Victor.1989.Planktonic larval duration of one hundred species of Pacific and Atlantic damselfishes (Pomacentridae).Mar.Biol. 101:557-567. [ Links ]
Wilson,D.T.&M.G.Meekan.2002.Growth-related advantages for survival to the point of replenishment on the coral reef fish Stegastes partitus (Pomacentridae). Mar.Ecol.Prog.Ser.231:247-260. [ Links ]

