










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.53 suppl.1 San José May. 2005
Chlorophyll a and turbidity patterns over coral reefs systems of La Parguera Natural Reserve,Puerto Rico
Ernesto Otero &Kelly K.Carbery
Department of Marine Science,University of Puerto Rico,Isla Magueyes Laboratories,Box 908,Lajas,Puerto Rico 00667. e-mail:eotero@uprm.edu
Received 15-I-2004. Corrected 07-X-2004. Accepted 29-IV-2005.
Abstract:Studies of temporal and spatial changes in phytoplankton biomass and turbidity provide essential information on coral reef ecosystem function and health.Fluctuation of phytoplankton biomass responds to several factors including nutrient inputs,both anthropogenic and natural,while turbidity is mostly affected by sediment resuspension or transport from terrestrial systems.These parameters can be used as sentinels of significant environmental factors "modifying "coral reef systems.A chlorophyll a concentration (Chl a )and turbidity (Turb)in situ logger was installed at 10 stations from June 4 to July 7,2003 in La Parguera Natural Reserve (Southwestern Puerto Rico)to assess short-term temporal and geographic variation in patterns of phytoplankton biomass and turbidity at pre-selected sites as part of an interdisciplinary long-term study.Average station Chl a variation was 0.17-1.12 µg l-1 and 0.2-23.4 NTU for Turb.Results indicate that the western near-coastal stations had higher levels of Turb and Chl a .The easternmost mid shelf station,Romero reef,was similar to coastal stations probably due to nutrient and suspended sediment inputs from a source external to our study area to the east,Guánica Bay.Comparisons between different sampling days indicate significant differences between days for most stations suggesting that one-time discrete sampling may not be representative of average water column conditions and illustrate the dynamic nature of coral reef systems.Further work is warranted to assess seasonal changes that integrate short-term (daily)variability in both Turb and Chl a .
Key words:Water quality,coral reef,turbidity,chlorophyll a ,in situ monitoring.
The water column provides connectivity among different coral reef habitats.This connectivity is expressed among other processes as diel fish migrations,transport/dispersal of plankton,larvae and pathogens,as well as dissolved and particulate matter (Andrefouet et al.2002).From a mechanistic point of view, the water column is the main interacting medium between coral reef systems and terrestrially dominated ones (estuaries and watersheds). Thus,studies of water quality are of paramount importance to provide clues to processes related to function and biodiversity of coral reef habitats, such as material transport within and among different coral reef systems,influx of land derived materials,species composition, and environmental health (Rogers 1990).
Phytoplankton biomass and turbidity have been frequently used as indicators of water quality in coral reef systems (Hunter and Evans 1995,Coles and Ruddy 1995).In general,fluctuation of phytoplankton biomass (expressed as chlorophyll a concentration;Chl a )responds to various factors including light and temperature regime,natural and anthropogenic nutrient sources,grazing (van Duyl et al.2002),and water residence time (Delesalle and Sournia 1992).Turbidity over coral reef systems is mostly controlled by sediment resuspension and/or transport from terrestrial systems and has been observed to be related to gorgonian and sponge abundance and inversely to scleractinian growth (Carricart-Ganivet and Merino 2001)effecting in such way modulation of coral reef biodiversity (Telesnicki and Goldberg 1995).Understanding the temporal and spatial patterns of Chl a and turbidity provides crucial information regarding ecologically significant fluctuations associated with coral reef function and health (Nemeth and Nowlis 1999).
Newer approaches have enabled continuous monitoring (Berkelmans et al.2002)of Chl a and turbidity by deployment of self-contained instrumentation over periods of weeks to months.These instruments overcome obstacles associated with discrete sampling by decreasing sampling effort while increasing temporal resolution.The utilization of this type of instruments permits a detailed assessment of the presence or absence of fluctuations in the quality of the water column habitat of coral reef systems and potential relationships with biological, physical and geomorphological patterns defining specific coastal settings (Rivera-Monroy et al.2004).This work describes the initial results of continuous monitoring of Chl a and turbidity in coral reef systems of La Parguera Natural Reserve,Puerto Rico.The data provide baseline information related to spatial and short term temporal variations of both variables within a wide range of environmental scenarios that help shape coral reef systems structural and functional framework within the study area.
Materials and methods
The study was conducted in La Parguera Natural Reserve,Southwestern Puerto Rico (Fig.1).Ten stations were established to encompass the range of terrestrial,coastal, and reef interactions that may be expressed as changes in water constituents.The position of these stations was determined using a Magellan 315/320 GPS unit.The reserve extends 8-10 km offshore and encompasses a series of habitats including coastal mangrove channels and lagoons,extensive seagrass beds, submerged, patch, platform, and shelf edge reef formations,as well as coastal embayments (García et al.1998).Environmental conditions in the region are characterized by southeasterly winds of approximately 24km/h and annual precipitation is low (<800mm;Anonymous 1983).However,sporadic intense precipitation on the order of 12 cm/24 hrs may occur,which induces significant terrestrial inputs of sediments and, furthermore, increases the chance of nutrient and sediment inputs from alloch-thonous sources to the east,such as Guánica Bay (Fig.1).Tides are semidiurnal with an average range of 0.5 m,precluding large tidal currents.Weather conditions during the study were characterized by clear skies,low or no rainfall,southeasterly winds of 24-32 km/h (wave heights of 0.5-1.5 m.)Such scenarios excluded possible observations of rain effects on our measurements and promoted increased flushing due to sustained windy conditions.In addition,the environmental setting during our study increased the opportunity for observation of sediment resuspension effects on turbidity induced by water currents in shallow areas or breaking waves in more exposed mid shelf reefs,especially during daytime peak winds.
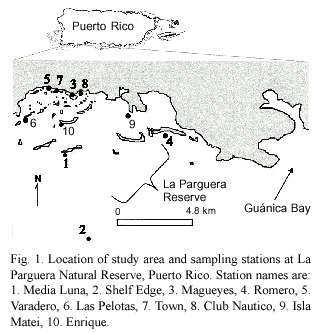
A Self Contained Underwater Fluorescence Apparatus (SCUFA;Turner Designs)was deployed consecutively at each of the above stations for periods of 24-48 hrs and programmed to record chlorophyll a fluorescence (Chl a FL)and relative turbidity (RelTurb) every 10 min from June 4 to July 7,2003. The SCUFA was anchored at deeper stations (7-20 m)keeping the sensor at least 0.6 m from the bottom.At shallow stations (1.5-2 m) the SCUFA was deployed using buoys 0.6 m under water.Instrument calibration was conducted in the laboratory before the campaign. Chlorophyll a calibration was conducted by recording in vivo fluorescence readings from the SCUFA using the provided SCUFASOFT software (Turner Designs)from samples collected from areas encompassing the expected range of Chl a (samples were collected from the same stations under study).Replicate 40 ml aliquots of each sample were filtered through 25 mm o.d.,0.4 µm Polycarbonate filters (Osmotics)for extractive analysis of Chl a after maceration and refiltration followed by fluorometric analysis per Otero et al.(2000).Regression analysis was used to derive calibration coefficients selecting zero (0)as y-intercept (Chl a =0.346Chl a Fl; r2 =0.68;n=10)Calibration for turbidity was conducted using laboratory prepared formazin standards (Wilde and Gibs 1998).Standards were prepared for a range of 0-40 nephelometric turbidity units (NTU),the SCUFA was dipped in such standards and its recordings regressed as described for Chl a (NTU=1. 1853RelTurb; r2 =0.99;n=8).Instrumental drift for Chl a was checked during sampling using a solid standard unit (Turner)between deployments and was <10%throughout the campaign.No visible fouling of the instrument lenses was observed at any time.Initial 20-min tests indicated SD s of 0.02 and 0.1 stability for Chl a Fl and Rel Turb (n=20), respectively.
Data smoothing was conducted by calculation of a four bin running average in order to dampen sudden spikes in the record possibly caused by large suspended particles.Samples were examined for uniformity of variances and the majority did not comply with the equal variance assumption of parametric analysis, therefore Kruskal Wallis One Way ANOVA with Dunn s multi-comparison analyses were conducted to determine differences among stations.Significant differences among sampling days at each station were determined using a Mann and Whitney U test on data for stations where the SCUFA was deployed for more than 48 hr,which excluded Romero,Varadero,and Magueyes stations from the analysis.
Results
Turbidity and Chl a time courses for three stations located at the outer shelf (shelf edge), an inner reef (Las Pelotas),and a coastal station (Town)are presented in Fig.2.Turbidity shows drastic fluctuations in the Town station frequently reaching 100 NTU while Chl a fluctuated independently reaching values of 3-13 µg l-1 .In contrast,the mid reef site,Las Pelotas, observed almost synchronized fluctuations in Chl a and Turbidity reaching values of approximately 0.3 µg l-1 and 1.7 NTU,respectively.
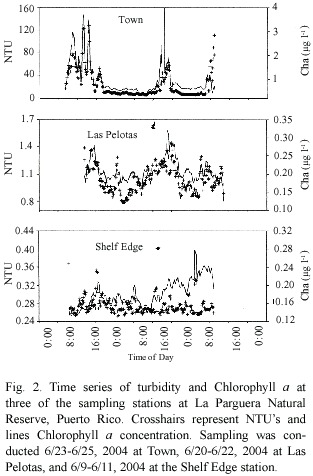
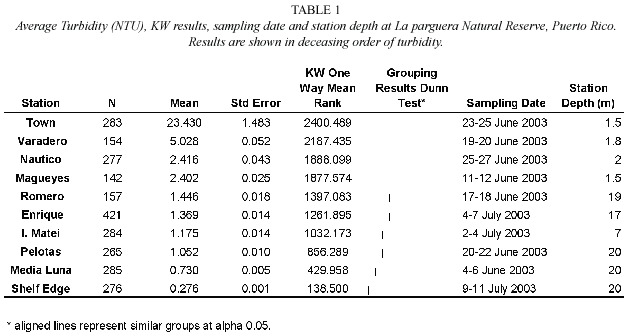
The lowest values for both measurements were observed at the Shelf Edge station where turbidity fluctuated from 0.5 to 0.7 NTU and Chl a showed an increasing trend over the 48 hr period from 0.13 to 0.22 followed by a sudden drop.Turbidity (Table 1)was higher near the coast with average station values of 1.2-23.4 NTU being maximal at the Town station.The average turbidity at Media Luna was significantly lower (50%)than at Romero,even though both reefs are similarly exposed to wave action.Coastal stations also contained higher Chl a than all other stations with the exception of Romero (Table 2)while the lowest Chl a values were found at the Shelf Edge and Enrique reefs. Examination of daily fluctuations demonstrated significant differences between days in most of the stations examined.There was no significant difference (p=0.95)for turbidity at the Shelf Edge,Las Pelotas,and Club Nautico stations while there was no difference in Chl a only for the Town station.Significant changes between days averaged 6-48%for turbidity and 10-27%for Chl a .
Discussion
Chlorophyll a concentration in the Great Barrier Reef (GBR)ranged from 0.16 to 0.7 µg l-1 according to Liston et al.(1992)and Furnas et al.(1990).In Curacao,average Chl a was found to be 0.26 to 1.1 µg l-1 with the higher values being found at the mouth of the local harbor and in sewage-contaminated reef waters (VanDuyl et al.2002).Turbidity,which may significantly modify coral reef assemblages, has been observed to attain a wide range of values in coral reef systems.Loya (1976)found higher coral cover (75%)in reefs of western Puerto Rico with a turbidity of 1.5 NTU while lower coral cover (<30%)was found in waters with 5.5 NTU.This is in agreement with findings by Coles and Ruddy (1995) where a positive relationship between Montastrea and Pocillopora spp.and turbidities lower than 1 NTU was observed.Although higher turbidity may influence the physiological state of corals, such as a decrease in P/R ratio (Telesnicki and Goldberg 1995),it does not preclude the establishment of high coral cover.Species adapted to such environments,such as Galaxaea sp., may reach up to 50%coral cover (Larcombe et al.2001)in marginal systems where high levels of diversity may be reached (Perry and Larcombe 2003).
Our results indicate a general land-ocean gradient probably due to the closer coupling of terrestrial,coastal,and bottom-derived materials in the water column in shallower regions that is reflected both in Turb and Chl a . Because of the predominant dry weather conditions during this effort,most of the land-sea gradient was presumably due to the effects of water turbulence in shallow water areas.The interaction between bottom features and waves and tidal currents may produce wave diffraction patterns,longshore currents (Otero et al. 1987),and eddies (Deleersnidjer et al.1992) that result in complex circulation patterns in coral reef systems expressed as spatially and temporally patchy distribution of water column constituents.The maximum turbidity and Chl a at the Town station indicates a source of suspended material and nutrients at that station or nearby to the east.The nearness of this station to the coastal village of La Parguera suggests the influence of anthropogenic effects due to higher coastal development,increased boat traffic,and occasional sewage inputs (faulty sewage distribution system) superimposed on the daily wind cycle,which can be observed as increments in these variables during the afternoon.In contrast,the coastal station,I.Matei, farthest to the east from La Parguera had even lower turbidity and Chl a than some mid and outer shelf reefs.Apart from lesser anthropogenic influence,the contrast of I.Matei with other coastal stations is probably due to its proximity to Montalva Bay,immediately eastward.Montalva Bay is fairly protected from waves by adjacent coral reefs making the bay a sink for suspended solids.Therefore, the outflow of water from the bay towards the I.Matei station results in lower turbidity than other coastal stations.
Romero reef was a clear outlier from the rest of the reef stations.Water at Romero reef remained murkier than I.Matei and contained similar Chl a as Varadero and Magueyes stations,thus,based on water column characteristics,Romero can be classified as a coastal station.Romero reef is one of the most wave-exposed reefs in our study.There is a high probability that the presence of higher turbidity and chlorophyll plumes is the result of preferential transport of fine particles by wave dissipation processes (Roberts et al.1992)in the vicinity of Romero.However,exposure to waves alone is unlikely to explain the higher content of materials at Romero since Media Luna,a similarly exposed reef,was only 50% as turbid and contained 60%of the Chl a as the Romero station.The proximity of Romero reef to terrestrial inputs from Guánica Bay to the east may explain this difference.
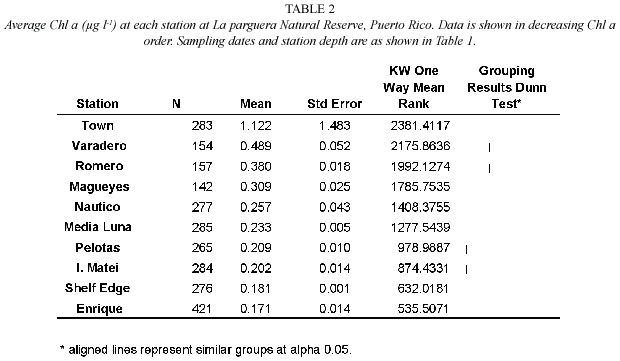
The areal distribution of Chl a had a more dynamic pattern than Turb based on similarities/ dissimilarities among stations.The Town and Shelf Edge stations had the maximum and minimum Chl a (Table 2)while Chl a at Romero and Magueyes stations followed Varadero and were grouped together.Chlorophyll a results suggest that 1)the inner western stations contained,received,or produced significantly higher phytoplankton biomass due to local inputs of nutrients and the transport generated by the general east-west wind regime and 2) phytoplankton biomass was higher towards the eastern/midshelf portion of the study area due either to biomass inputs or nutrient enhanced phytoplankton production from the east.This is supported by the presence of lower Chl a values towards the central-western midshelf region of the study area suggesting the presence of nutrient sources towards the east.
Other reef-related processes are known to produce significant short-term shifts in phytoplankton (Chl a )biomass that might contribute to the observed Chl a patterns,such as nutrient competition between benthic algae and phytoplankton (Fong et al.1993)and depletion of planktonic microbial species by benthic filter feeders (Van Duyl et al.2002).Although grazing of zooplankton exert top-down control on phytoplankton,it is mainly dependent on population density (Valiela 1995),thus,their role in controlling phytoplankton distribution relative to the previously mentioned factors is uncertain in the present study.
The interplay of the above physical and biological processes resulted in the observed daily fluctuations in Chl a and turbidity in most of the stations.The range of daily changes (differences between daily averages)was 6- 48%for turbidity and 10-27%for Chl a at stations found to have significant changes in those variables.Chl a fluctuation was lower than that observed by Liston et al.(1992) who found approximately 50%oscillation in chlorophyll fluorescence in bottom waters of the GBR with no apparent link to tidal currents or water intrusion.Our data indicate the presence of significant short-term changes in water constituents in coastal areas and some reefs of our study site.Bell et al.(1993)have proposed an eutrophication threshold of 0.3 µg l-1 Chl a as indicator of detrimental nutrification effects that could have given rise to the demise of acroporid species in the GBR.Our data show values close to that threshold mostly at coastal stations (14%of the observations at I.Matei and Club Nautico and 76-100%from Magueyes and westward were at or higher than 0.3 µg l-1 Chl a ).Also,Chl a at Romero reef was frequently over 0.3 µg l-1 (85%of the observations)while the other reef sites were close to that Chl a =3%of the time.Overall, we consider these variations minimal since other factors that influence the distribution of Chl a and turbidity,such as rainfall,and terrestrial and estuarine inputs,were minimal during the period of this study.
Our examination of Chl a and turbidity levels in the study area suggests inshore systems as significant sources of materials (nutrients and suspended solids)especially near the town of La Parguera,suggesting anthropogenic influences as a major factor enhancing phytoplankton biomass production and/or resuspension of bottom sediments. Additionally, Romero reef may be classified as a coastal station based on water column characteristics,which indicate the presence of nutrient sources towards the southeastern portion of our study site.Our results underscore the importance of considering daily fluctuations in both parameters especially under this complex set of coral reef interactions, and,furthermore,in order to establish seasonal and long-term water quality trends.
Acknowledgments
The authors thank Lumarie Perez and Yaritza Rivera for their help in the laboratory. Thanks are also due to Lisamarie Carrubba for editorial help during the preparation of the manuscript.The work effort described below is part of a larger interdisciplinary initiative on Coral Reef Ecosystem Studies (CRES)funded by NOAA NOS CSCOR COP.
Resumen
Los estudios sobre cambios temporales y espaciales de biomasa fitoplanctónica y de urbidad, proveen información esencial sobre la función y salud de los sistemas arrecifales.Fluctuaciones de la biomasa de fitoplancton responden a factores como entradas de nutrientes de fuentes antropogénicas y/o naturales,mientras que la turbidez responde mayormente a la resuspensión de sedimentos o la entrada de sedimentos de sistemas terrestres.Por tal motivo,estos parámetros pueden ser indicadores de factores ambientales que modifican los arrecifes coralinos.Un recolector in situ,de datos de concentración de clorofila a (Chl a )y turbidez (Turb),fue instalado en 10 estaciones del 4 de junio al 7 de julio del 2003 en la Reserva Natural de La Parguera (Suroeste de Puerto Rico)para establecer patrones geográficos y temporales de biomasa de fitoplancton y turbidez como parte de un estudio interdisciplinario. La variación promedio de Chl a por estación fue 0.17-1.12 µg l-1 mientras para Turb fue 0.2-23.4 NTU.Los resultados indican que las estaciones costeras al oeste tenían niveles mayores de Turb y Chl a mientras que las aguas de un arrecife en la plataforma intermedia al este,Romero, tenían características similares a las estaciones costeras. Esto se debe a la proximidad de Romero a una fuente de nutrientes y sedimentos suspendidos al este del área de estudio,la Bahía de Guánica.Se encontraron diferencias significativas para Chl a y Turb entre diferentes días de muestreo en la mayor parte las estaciones,lo que sugiere que muestreos discretos no son necesariamente representativos de las condiciones promedio de la columna de agua e ilustra lo dinámico de los sistemas arrecifales.Se necesita trabajo que integre la variabilidad a corto plazo (diaria)en el estimado estacional de Chl a y Turb.
Palabras clave:Calidad de agua,arrecife coralino,turbidez,clorofila a,monitoreo in situ .
References
Andrefouet,S.,P.J.Mumby,M.McField,C.Hu &F.E. Muller-Karger.2002.Revisiting coral reef connectivity.Coral Reefs 21:43-48. [ Links ]
Anonymous.1983.Preliminary Environmental Impact Statement and Management Plan for La Parguera National Marine Sanctuary.Puerto Rico Department of Natural Resources.PRDNR.San Juan,Puerto Rico. [ Links ]
Bell,P.,T.Tomascik &F.G.Walton-Smith.1993.The demise of the fringing coral reefs of Barbados and of regions in the Great Barrier Reef (GBR)lagoon -impacts of eutrophication,p.1-7.In R.N.Ginsburg (Comp.).Global Aspects of Coral Reefs:Health, Hazards and History.RSMAS,Univ.Miami. [ Links ]
Berkelmans,R.,J.C.Hendee,P.A.Marshall,P.V.Ridd,A.R. Orpin &D.Irvine.2002.Automatic weather stations: Tools for managing and monitoring potential impacts to coral reefs.Mar.Technol.Soc.J.36:29-38. [ Links ]
Carricart-Ganivet,J.P.&M.Merino.2001.Growth response of the reef-building coral Montastrea annularis along a gradient of continental influence in the southern Gulf of Mexico.Bull.Mar.Sci.68:133-166. [ Links ]
Coles,S.L.&L.Ruddy.1995.Comparison of water quality and reef coral mortality and growth in southeastern Kaneohe Bay,Oahu,Hawaii,1990 to 1992,with conditions before sewage diversion.Pac.Sci.49: 247-265. [ Links ]
Deleersnidjer,E.,A.Norro &E.Wolanski.1992.A three- dimensional model of the water circulation around an island in shallow water.Cont.Shelf Res.12: 891-906. [ Links ]
Delesalle,B.&A.Sournia.1992.Residence time of water and phytoplankton biomass in coral reef lagoons. Cont.Shelf Res.12:939-950. [ Links ]
Fong,P.,R.M.Donohoe &J.B.Zedler.1993.Competition with macroalgae and benthic cyanobacterial mats limits phytoplankton abundance in experimental microcosms.Mar.Ecol.Prog.Ser.100:97-102. [ Links ]
Furnas,M.J.,A.W.Mitchell,M.Gilmartin &N.Revelante. 1990.Phytoplankton biomass and primary production in semi-enclosed reef lagoons of the central Great Barrier Reef,Australia.Coral Reefs 9:1-10. [ Links ]
Garcia,J.R.,C.Schmitt,C.Heberer &A.Winter.1998. La Parguera,Puerto Rico,USA,p.195-212.In B. Kjerfve (ed.).Caribbean Coral Reef,Seagrass and Mangrove Sites.Coastal Region and Small Island Series.CARICOMP/Unesco,Paris. [ Links ]
Hunter,C.L.&C.W.Evans.1995.Coral reefs in Kaneohe Bay,Hawaii:Two centuries of western influence and two decades of data.Bull.Mar.Sci.57:501-515. [ Links ]
Larcombe,P.,A.Costen &K.J.Woolfe.2001.The hydro-dynamic and sedimentary setting of nearshore coral reefs,central Great Barrier Reef shelf,Australia: Paluma Shoals,a case study.Sedimentology 48: 811-835. [ Links ]
Liston,P.,M.J.Furnas,A.W.Mitchell &E.A.Drew.1992. Local and mesoscale variability of surface water temperature and chlorophyll in the northern Great Barrier Reef,Australia.Cont.Shelf Res.12:907-922. [ Links ]
Loya,Y.1976.Effects of water turbidity and sedimentation on the community structure of Puerto Rican corals. Bull. Mar. Sci. 26:450-466. [ Links ]
Nemeth,R.S.&J.S.Nowlis.1999.Monitoring the effects of land development on the near-shore reef environment of St.Thomas,USVI.Bull.Mar.Sci.69: 759-775. [ Links ]
Otero,E.,F.Nieves &J.E.Corredor.1987.Patterns of tar ball accumulation on a Lunate Coral Key at La Parguera, Puerto Rico.Carib.J.Sci.23:123-129. [ Links ]
Otero,E.,R.Culp,J.Noakes &R.Hodson.2000. Allocation of particulate organic carbon from different sources in two contrasting estuaries of southeastern USA.Limnol.Oceanogr.45:1753-1763. [ Links ]
Perry,C.T.&P.Larcombe.2003.Marginal and non- reef-building coral environments.Coral Reefs 22: 427-432. [ Links ]
Rivera-Monroy,V.H.,R.R.Twilley,D.Bone,D.L.Childers, C.Coronado-Molina,I.C.Feller,J.Herrera-Silveira, R. Jaffe, E. Mancera, E.Rejmankova,J.E.Salisbury &E.Weil.2004.A conceptual framework to develop long-term ecological research and management objectives in the wider Caribbean region. Bioscience 54:843-856. [ Links ]
Roberts,H.H,P.A.Wilson &A.Lugo-Fernandez.1992. Biologic and geologic responses to physical processes: examples from modern reef systems of the Caribbean- Atlantic region Cont.Shelf Res.12:809-834. [ Links ]
Rogers,C.S.1990.Responses of coral reefs and reef organisms to sedimentation.Mar.Ecol.Prog.Ser. 62:185-202. [ Links ]
Telesnicki,G.J.&W.M.Goldberg.1995.Effects of turbidity on photosynthesis and respiration of two south Florida reef coral species.Bull.Mar.Sci.57: 527-539. [ Links ]
Valiela,I.1995.Marine Eological Processes,2nd Ed. Springer-Verlag.New York.686p. [ Links ]
Van Duyl,F.C.,G.J.Gast,W.Steinhoff,S.Kloff,M.J.W. Veldhuis &R.P.M.Bak.2002.Factors influencing the short-term variation in phytoplankton composition and biomass in coral reef waters.Coral Reefs 21:293-306. [ Links ]
Internet reference
Wilde,F.D &J.Gibs.1998.Calibration (Turbidity): U.S.Geological Survey Techniques of Water-Resources Investigations,Book 9,Chap.A6.,Sect. 6.7,(Also available on-line http://water.usgs.gov/owq/FieldManual /Chapter6/6.7.2.html downloaded May 3,2004). [ Links ]

