Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.53 n.1-2 San José Jun. 2005
Aves acuáticas de la Laguna de Agua Dulce y el Estero El Ermitaño, Jalisco, México
Salvador Hernández Vázquez
Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras. Universidad de Guadalajara. Gómez Farias No. 82, San Patricio-Melaque, Municipio de Cihuatlán, Jalisco, CP 48980. México. Fax:(335)56331; sahernan@costera.melaque.udg.mx
Recibido 26-V-2004. Corregido 20-VII-2004. Aceptado 11-VIII-2004.
Abstract
Water birds from Agua Dulce lake and El Ermitaño estuary, Jalisco, Mexico. Waterbird abundance, and seasonal and spatial distribution, were studied in two natural water pools at Jalisco, México, from December 1997 through November 1998. Maximum monthly abundance in Agua Dulce lake and El Ermitaño estuary was 86 471 birds (29 686 in Agua Dulce and 56 785 in Ermitaño), with a total cummulative abundance of 179 808 individuals (66 976 in Agua Dulce and 112 832 in Ermitaño). A total of 87 waterbirds species were recorded, 78 in Agua Dulce and 73 in Ermitaño.The higher species richness and abundance was observed during winter, when migratory species arrived. Most species prefered shallow waters, except seabirds which prefered protected areas such as dunes in Agua Dulce. Other groups, like ducks and related species, prefered low salinity areas, for example in the south-east area of Ermitaño. The higher abundance of the shorebirds was found when the water level on the estuary was low. Herons were seen often at areas with high salinity and influenced by tides (e.g. mouth of Ermitaño). Rev. Biol. Trop. 53(1-2): 229-238. Epub 2005 Jun 24.
Key words: Coastal waterbirds, seasonality, distribution, Agua Dulce lagoon, El Ermitaño estuary, Jalisco, México.
Los ecosistemas acuáticos costeros son considerados como los ecosistemas más productivos del mundo (Day et al .1989, Costanza et al .1993, Barbier et al .1996), y en ellos se desarrollan procesos biogeoquímicos importantes para muchas especies de plantas y animales (Costanza et al. 1993). Es por esto que los altos niveles de productividad de los ambientes acuáticos permiten que puedan sustentar muchas especies de aves migratorias y residentes (Warnock y Takekawa 1995), particularmente las que tienen hábitos acuáticos (Davidson et al .1991, Ens et al. 1994).
Las aves acuáticas neárticas utilizan los humedales costeros de México ya sea para permanecer durante la temporada de invierno o para realizar paradas de descanso o recuperación de energéticos durante su migración hacia el sur o hacia el norte. De esta manera evitan el estrés fisiológico de los climas desfavorables y pueden explotar los recursos alimenticios que están disponibles sólo durante una parte del año (Howes y Bakewell 1989). Este puede ser el caso de los humedales de Jalisco, y en particular de la laguna de Agua Dulce y del estero El Ermitaño, caracterizados por ser áreas pequeñas y con una gran diversidad de hábitats, lo que atrae a un mayor número de aves (Hernández-Vázquez y Mellink 2001).
Dentro de este contexto, no hay mucha información sobre las aves estuarinas en el municipio de Tomatlán, y en particular de la laguna Agua Dulce y estero El Ermitaño, por lo que el objetivo de este estudio es realizar la primera descripción detallada de la composición de especies, abundancia, distribución estacional y espacial de las aves acuáticas con respecto al nivel del agua y salinidad.
Materiales y métodos
Área de estudio: El sistema lagunar Agua Dulce-Ermitaño se encuentra en la en la costa del Pacifico Mexicano, en la reserva Playón de Mismaloya, Jalisco, México (19º 58 - 20º 05 N y 105º 27 - 105º 33 W). La laguna de Agua Dulce. Se ubica en la playa denominada "Mismaloya", a 4 km de el poblado Portezuelos. Tiene una superficie de 696 hectáreas con una orientación de noroeste a sureste y profundidades que varían entre 2 y 4 m. Su principal aporte de agua es por el estero El Ermitaño ya que se encuentra unido a este por una compuerta artificial. Alrededor de la laguna hay una superficie aproximada de 604 m2 de selva baja caducifolia al margen del cerro rocoso, una planicie de Batis maritima presente con diferentes densidades a todo el margen de la laguna, al igual que Laguncularia racemosa y Conocarpus erectus que en el extremo noroeste y sureste alcanzan una altura de hasta tres metros. También existe la presencia de Thypha sp.,en su extremo pegado al mar se extienden grandes dunas que albergan a Acacia sp., Prosopis sp. y Opuntia sp. El estero El Ermitaño tiene una superficie de 318 hectáreas con profundidades entre 0.8 a 3 m. Sus aportes de agua más importantes es por el río María García y de los canales de irrigación provenientes de la presa Cajón de Peña. La comunicación con el mar se lleva a través de una boca-barra que se encuentra al noroeste y depende de la apertura de la boca-barra de conexión. La orilla más próxima a la porción continental de está franja costera se encuentra circundada por L. racemosa, Thypha sp. y Pragmites australis.
Metodología: Se realizaron visitas mensuales en los dos sitios de estudio, entre diciembre de 1997 a noviembre de 1998. En cada visita se realizaron recorridos a lo largo del borde interno del cuerpo de agua y de las zonas de playa contiguas a estos. El borde interno del cuerpo de agua se recorrió utilizando una lancha de 5-7 m de eslora con un motor fuera de borda de 6.5 caballos de fuerza. Los censos se iniciaron por las mañanas y cada censo requirió entre 3 a 4 h. Las aves se identificaron de forma visual con la ayuda de binoculares 10x50 y telescopio 15-60x. El nombre científico se realizó con base en el A. O. U.(1998). Las aves se clasificaron en cuatro grupos con base a la forma de obtener su alimento: aves marinas, patos y afines, aves playeras, garzas y afines, y un quinto grupo denominado "otros" donde se agruparon especies de las familias Anhingidae, Accipitridae, Jacanidae y Alcedinidae.
Se buscaron diferencias en el número de especies e individuos entre los dos esteros por medio del análisis no paramétrico de Friedman a un nivel 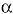 =0.05 (Zar 1974). La densidad fue obtenida dividiendo el número total de aves entre la superficie del humedal y fue utilizada para comparar el número de aves de cada grupo entre los dos humedales. La relación entre el nivel del agua, salinidad y el número de aves de cada grupo se analizó con el coeficiente de correlación de rangos de Spearman (rs ) (Zar 1974).
=0.05 (Zar 1974). La densidad fue obtenida dividiendo el número total de aves entre la superficie del humedal y fue utilizada para comparar el número de aves de cada grupo entre los dos humedales. La relación entre el nivel del agua, salinidad y el número de aves de cada grupo se analizó con el coeficiente de correlación de rangos de Spearman (rs ) (Zar 1974).
Resultados
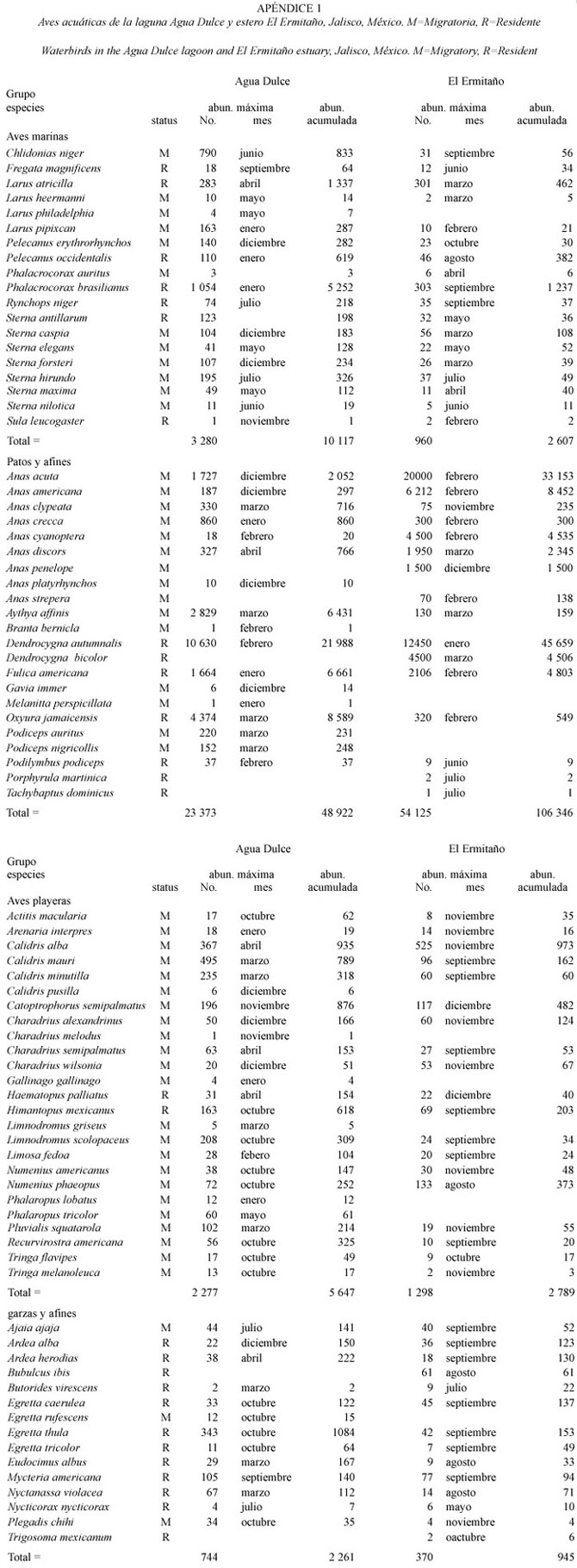
Abundancia y composición de especies. Considerando únicamente el mes de máxima abundancia para cada especie en Agua Dulce y Ermitaño hubo 86 471 aves (29 686 en Agua Dulce y 56 785 en El Ermitaño) y una abundancia acumulada (suma de las abundancias registradas mensualmente) de 179 808 aves (66 976 en Agua Dulce y 112 832 en El Ermitaño) (Apéndice 1). En las 12 visitas se identificaron 87 especies de aves asociadas a los cuerpos de agua. En la laguna de Agua Dulce se observó a un total de 78 especies; 19 aves marinas, 17 patos y afines, 25 aves playeras, 13 garzas y afines y en "otros" sólo cuatro especies. En el estero El Ermitaño hubo 73 especies; 18 aves marinas, 16 patos y afines, 19 aves playeras, 14 garzas y afines y seis especies dentro del grupo de "otros" (Apéndice 1).
Las aves marinas compartieron en ambos humedales un total de 18 especies de las cuales Phalacrocorax brasilianus y Larus atricilla fueron las más abundantes. Larus philadelphia sólo fue observada en Agua dulce (Apéndice 1). De patos y afines hubo 11 especies en ambos cuerpos de agua, de las cuales la más abundante en Agua Dulce fue Dendrocygna autumnalis y Oxyura jamaicensis y en El Ermitaño fue D. autumnalis y Anas acuta. Cinco especies sólo fueron observadas en Agua Dulce (Anas platyrhynchos, Branta bernicla, Gavia immer, Melanitta perspicillata, Podiceps auritus y Podiceps nigricollis ) y cinco en El Ermitaño (Anas penelope, Anas strepera, Dendrocygna bicolor, Porphyrula martinica y Tachybaptus dominicus). Del grupo de aves playeras 19 estuvieron presentes en los dos cuerpos de agua, las especies más abundante en ambos sitios fueron Calidris alba y Catoptrophorus semipalmatus. Hubo seis especies observadas únicamente en Agua Dulce (Calidris pusilla, Charadrius melodus, Gallinago gallinago, Limnodromus griseus,Phalaropus lobatus y Phalaropus tricolor ). En ambos humedales las garzas y afines compartieron 12 especies, siendo Egretta thula la más abundante en ambos sitios. Egretta rufescens sólo fue observada en Agua Dulce y Bubulcus ibis y Tigrisoma mexicanum sólo en El Ermitaño. El grupo "otros" tuvo abundancias, por lo que no fue incluido en los análisis y sólo se mencionan las especies con el número de aves (Apéndice 1).
Variación estacional. Los dos cuerpos de agua fueron similares en el número de especies ( 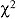 r, P>0.05, para cada grupo de aves). En Agua Dulce la densidad de especies del grupo de aves marinas fue constante durante las cuatro estaciones del año, mientras que en El Ermitaño se presentaron tres fluctuaciones; a finales del invierno, a finales de la primavera y durante el verano. Los patos y afines tuvieron sus mayores valores de la densidad de especies durante el invierno. En ambos humedales las especies de aves playeras tuvieron valores altos en invierno, verano y otoño. El grupo de garzas y afines se mantuvieron constantes durante las cuatro estaciones del año, aunque este fue mayor en El Ermitaño (Fig.1).
r, P>0.05, para cada grupo de aves). En Agua Dulce la densidad de especies del grupo de aves marinas fue constante durante las cuatro estaciones del año, mientras que en El Ermitaño se presentaron tres fluctuaciones; a finales del invierno, a finales de la primavera y durante el verano. Los patos y afines tuvieron sus mayores valores de la densidad de especies durante el invierno. En ambos humedales las especies de aves playeras tuvieron valores altos en invierno, verano y otoño. El grupo de garzas y afines se mantuvieron constantes durante las cuatro estaciones del año, aunque este fue mayor en El Ermitaño (Fig.1).
El número de individuos varió entre los dos cuerpos de agua ( r, P<0.05, para cada grupo de aves), excepto las garzas y afines ( r, P>0.05). En Agua Dulce las aves marinas tuvieron dos incrementos en la densidad; el primero en invierno y el segundo durante el verano. En El Ermitaño también se observaron dos incrementos en la densidad; el primero a finales del invierno y el segundo en otoño (Fig.2). Los mayores valores en la densidad de patos y afines fueron observados durante el invierno, auque estos fueron mucho mayor en el Ermitaño. Las aves playeras tuvieron en Agua Dulce dos fluctuaciones en la densidad; uno a finales de invierno y otro en otoño, siendo mayor el primero. En El Ermitaño también hubo dos picos en la densidad; el primero a finales de verano y el segundo en otoño. La densidad de las garzas y afines fue constante en ambos cuerpos de agua, con un ligero incremento en septiembre y octubre.
En Agua Dulce y el Ermitaño no hubo una correlación entre la densidad de individuos y el nivel del agua (rs= Spearman, p >0.05, para cada grupo de aves), excepto en las aves playeras (rs= Spearman, p<0.05), por lo que las mayores abundancias en la densidad de las aves playeras fueron registradas en niveles de agua más bajos.
Distribución espacial. Los diferentes grupos de aves presentaron diferencias en los patrones de distribución en ambos cuerpos de agua (Fig.3). Considerando las abundancias acumuladas durante el periodo de estudio se observó que en Agua Dulce hubo un mayor número de aves marinas en la boca-barra (30%), el canal (22%) y en áreas arenosas de la parte oeste (20%). En El Ermitaño este grupo de aves fueron observadas más frecuentemente en la boca-barra y en la playa (37%).
En Agua Dulce los patos y afines tuvieron una clara preferencia por sitios protegidos y someros como los que se encuentran en el canal (89%) donde se registraron las salinidades más bajas (± EE) (20.4 ± 2.13 0/00). En El Ermitaño este grupo de aves se concentraron principalmente en la parte sureste del estero (70%) y en la parte central (20%), donde se registraron los valores más bajos en la salinidad (± EE) (1.33 ± 0.6 0/00 y 2.75 ± 1.98 0/00, respectivamente).El grupo de aves playeras prefirió en Agua Dulce la boca-barra (40%) y el canal (38%), en el primer sitio hubo salinidades intermedias (25.8 ± 0.6 0/00 y más bajas en el canal (20.4 ± 2.13 0/00). En El Ermitaño prefirieron la playa junto a la boca-barra (70%), donde se registraron las salinidades más altas (3.16 ± 2.04 0/00).
Aunque el grupo de garzas y afines se observó distribuido en todo el cuerpo de agua, el mayor porcentaje de aves observadas en Agua Dulce se registro en los manglares localizados en el canal (35%) y en la parte este de la laguna (25%), mientras que en El Ermitaño las mayores abundancias fueron observadas en la orilla del espejo de agua de la parte este del estero (38%) y en la desembocadura del río (32%).
Discusión
Abundancia y composición de especies. En ambos cuerpos de agua se registraron 87 especies de aves acuáticas, lo que representa el 30% de las especies acuáticas reportadas para México y el 73% de las especies acuáticas litorales reportadas para la costa de Jalisco (Howell y Webb 1995). Estos valores en el número de individuos y de especies son superiores a los reportados para otros humedales de la costa de Jalisco (Hernández-Vázquez 2000, Cupul-Magaña 2000, Hernández-Vázquez y Mellink 2001). La alta abundancia y riqueza de especies de Agua Dulce y El Ermitaño se atribuye a que representan unos de los cuerpos de agua con mayor tamaño de la costa de Jalisco, propiciando una mayor diversidad y extensión de hábitats.
En Agua Dulce fueron observados en enero dos individuos de Melanita perspicillata y en febrero dos individuos de Branta bernicla. En ambos casos las aves se encontraron descansado en áreas someras, rodeadas de vegetación densa. Estas especies han sido reportadas para Baja California, Sinaloa y Nayarit (Howell y Webb 1995), por lo que estos son los primeros registros para el estado de Jalisco. En diciembre se observó a 1500 individuos de Anas penelope alimentándose en El Ermitaño. Aunque esta especie ya había sido reportada para las tierras altas de Jalisco (Howell y Webb 1995), este representa el primer registro para la zona costera del estado. En todos los casos las especies fueron observadas durante periodos cortos alimentándose y en descanso.
Variación estacional. Los mayores incrementos en la abundancia de aves marinas observados en los dos cuerpos de agua fueron determinados principalmente por la presencia de P. brasilianus y en menor cantidad por la llegada de L. atricilla. La primera especie es un ave residente abundante en la zona. L atricilla es una especie reportada para Jalisco como visitante de invierno (Howell y Webb 1995).
Las especies que más influyeron en el número de patos y afines fue D. autumnalis (marzo y diciembre) y A. acuta. Con base a recorridos de campo se constató que durante el periodo reproductivo de D. autumnalis (mayo- agosto) los individuos se desplazaron fuera de los ambientes acuáticos en busca de áreas adecuadas para anidar, como vegetación densa y huecos de árboles, etc. Una vez concluido el periodo reproductivo un mayor número de individuos se concentran en los humedales costeros, donde encuentran lugares óptimos para alimentarse y descansar, este es el caso de Agua Dulce y El Ermitaño. A. acuta fue registrada durante todo el invierno, lo que indica que estos cuerpos de agua y principalmente El Ermitaño son un sitio importante para esta especie migratoria, donde puede satisfacer sus necesidades de alimentación y descanso durante el invierno.
Las mayores densidades de aves playeras se presentaron en condiciones de boca-barra abierta (noviembre, diciembre, abril y septiembre) y con niveles de agua bajos. Esta correlación negativa entre el nivel del agua y el número de aves playeras fue más evidente en El Ermitaño, donde la variación en las mareas propició un aumento en las áreas de alimentación. Este patrón no fue muy evidente en Agua Dulce debido a que durante el estudio no tuvo contacto directo con el mar, y sus ligeras variaciones en el nivel del agua estuvieron determinadas por el aporte de agua de El Ermitaño a través de una compuerta artificial. Entre las especies más abundantes se encuentran C. alba y C. mauri . Estas especies se comportaron como migratorias transitorias, ya que sus abundancias fueron observadas durante un corto periodo cuando realizaban sus migraciones hacia los sitios de invernación ubicados más al sur, o sus sitios de reproducción localizados al norte.
El ligero incremento de las garzas y afines observado en Agua Dulce entre septiembre y octubre se debió a la presencia de E. thula, y en El Ermitaño se debió a la presencia de algunas especies migratorias, como M. americana, A. ajaja y E. caerulea.
Distribución espacial. Las aves marinas es un grupo que se alimenta principalmente en ambientes marinos (Barber y Chávez 1983, Schreiber y Schreiber 1984), y su presencia en los ambientes acuáticos costeros se basa principalmente en sitios para descansar. Es por esto que la preferencia de la mayoria de las aves marinas en Agua Dulce está explicada por las áreas someras arenosas localizadas en la boca-barra y el borde ubicado al oeste de la laguna, en donde las aves se pueden posar para descansar. Además de que las extensas dunas de más de cinco metros de altura que separan la laguna del mar le proporcionaron protección contra el viento y tormentas tropicales. En El Ermitaño este grupo fue observado mayormente en las áreas arenosas que se encuentran en la boca-barra. En ambos cuerpos de agua la mayoría de las aves de este grupo se les observó en descanso.
La mayor abundancia de patos y afines se observó en aguas someras y con salinidades bajas, principalmente donde se da el aporte de agua dulce al estero, coincidiendo con lo observado por otros autores (Burger et al. 1984, Collier y Wakelin 1996, Ysebaert et al. 2000). Es en estos sitios donde el exceso de materia orgánica que se origina con las descargas de los ríos, origina un aumento en algunos invertebrados, como pequeños poliquetos, los cuales se ha demostrado que forman parte de la dieta de A. discors, A. platyrhynchos (Baldasarre et al. 1989, Kramer y Migoya 1989). De igual forma en áreas someras la vegetación de la que se alimentan los patos es más accesible desde la superficie, requiriendo menos tiempo y energía para alimentarse (Burger et al. 1984). En Agua Dulce estas condiciones se observaron principalmente en el canal localizado al sureste de la laguna, mientras que en El Ermitaño se localizan en la parte este y sureste del estero.
Las aves playeras son un grupo correlacionado positivamente con la abundancia y variedad de invertebrados (Kawaji y Shiraishi 1979, Colwell y Landrum 1993, Colwell 1993) quienes forman parte importante de su dieta (Hayes y Fox 1991, Colwell y Landrum 1993, Colwell 1993). Estos invertebrados se caracterizan por vivir principalmente en zonas intermareales y en fondos con sedimentos blandos (Ysebaert et al. 2000). Estas condiciones son encontradas principalmente en la boca barra donde la influencia de las mareas originó una mayor disponibilidad de hábitats de forrajeo como las planicies lodosas y arenosas, siendo esto un atractivo para un número más grande de aves playeras, lo que concuerda con lo observado por otros autores en otros sitios (Hayes y Fox 1991, Colwell y Landrum 1993, Colwell 1993).
El grupo de garzas y afines se distribuyó prácticamente en todos los márgenes de ambos cuerpos de agua. La amplia distribución de este grupo de aves puede atribuirse a la variedad de organismos que forman parte de su dieta (insectos, pequeños reptiles, roedores) (Custer et al. 1996). Sin embargo, un mayor número de aves fue observado en áreas más someras. La disponibilidad de alimento en ambientes someros acuáticos, como los observados en Agua Dulce y El Ermitaño, es influenciada por los cambios en el nivel de agua; la densidad de peces se reduce cuando los niveles de agua son elevados. Además de que en niveles de agua altos se restringen los hábitats de forrajeo (Custer et al. 1996), afectando la tasa de alimentación de las garzas (David 1994). En El Ermitaño un mayor número de individuos se desplazaron a áreas donde el alimento de origen marino es más accesible y en donde la riqueza, biomasa y abundancia de peces es mayor, como es el caso de la boca-barra del estero, donde las características físicas y biológicas que prevalecen por su cercanía con el mar, y cuyos procesos hidrográficos permiten que peces marinos juveniles penetren al sistema (Vega-Cendejas 1998).
Los resultados obtenidos en el presente estudio indican que La laguna de Agua Dulce y el estero El Ermitaño son los humedales de la costa de Jalisco que albergan a un mayor número de especies e individuos durante el periodo migratorio. Estos humedales mostraron ser de importancia para las aves, especialmente para las aves migratorias como un sitio de descanso y alimentación donde almacenaron energía para continuar su viaje hacia el sur o al norte, excepto para algunas especies de patos, las cuales aparentemente pasaron todo el invierno con abundancias considerables.
Agradecimientos
Se agradece a la Sociedad Cooperativa de Producción Pesquera "Cruz de Loreto". A C. Valadéz-González, B. Cesar Durand, R. Esparza Salas y L. F Alvarado Ramos por el apoyo proporcionado en el trabajo de campo. Este trabajo fue financiado por la Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad "CONABIO" (Convenio Núm. FB425/L292/97), el Canadian Wildlife Service/Latin American Program, y el Centro Universitario de la Costa Sur, de la Universidad de Guadalajara.
Resumen
Se analizó la presencia y número de aves acuáticas, así como la distribución estacional y espacial de los diferentes grupos en la laguna de Agua Dulce y el estero El Ermitaño, Jalisco, México, entre diciembre de 1997 a noviembre de 1998. Considerando únicamente el mes de máxima abundancia de cada especie hubo 86 471 aves en ambos humedales (29 686 en Agua Dulce y 56 785 en El ermitaño), y una abundancia acumulada (suma de las abundancias mensuales) de 179 808 aves (66 976 en Agua Dulce y 112 832 en Ermitaño). Se identificaron 87 especies de aves, de las cuales 78 especies estuvieron presentes en Agua Dulce y 73 especies en El Ermitaño. La riqueza y abundancia fueron influenciados por la llegada de aves migratorias por lo que el mayor número de especies e individuos se presentó durante el invierno. En general las aves mostraron preferencia por áreas someras, excepto las marinas que prefirieron áreas arenosas y protegidas por dunas como las que se encuentran en Agua Dulce. Los demás grupos como los patos y afines prefirieron las áreas con salinidades bajas, como las que se encuentran al sureste del estero El Ermitaño. La mayor abundancia de aves playeras fue observada en niveles de agua bajos, y al igual que las garzas se les observó con mayor frecuencia en sitios con salinidades altas e influenciados por las variaciones en las mareas, como los que se encuentran cerca de la boca-barra del Ermitaño.
Palabras clave: aves acuáticas costeras, estacionalidad, distribución, laguna Agua Dulce, estero El Ermitaño, Jalisco, México.
Referencias
A. O. U. 1998. Check-list of North American Birds. The American Ornithologists´ Union. Washington, D.C. 829 p. [ Links ]
Baldasarre, G. A., A. R. Brazada & E. R. Woodyard. 1989. The east coast of México. In Smith, L. M., R. L. Pederson & R. M. Kaminski (eds.). Habitat Management for Migrating and Wintering Waterfowl in North America. Texas Tech University, Lubbock. pp. 407-425. [ Links ]
Barber, R. T. & F. P. Chávez. 1983. Biological consequences of El Niño. Science 222:1203-1210. [ Links ]
Barbier, E. B., M. Acreman & D. Knowler. 1996. Economic valuation of wetland: a guide for policy makers and planners. Ramsar Convention Bureau, Gland, Switzerland. [ Links ]
Burger, J., J. R.Trout, W. Wander & G. S. Ritter. 1984. Jamaica Bay Studies VII: factors affecting the distribution and abundance of ducks in a New York estuary. Estuarine, Coastal Shelf Sci. 19:673-689. [ Links ]
Colwell, M. A. 1993. Shorebird community patterns in a seasonally dynamic estuary. Condor 95:104-114. [ Links ]
Colwell, M. A. & S. L. Landrum. 1993. Nonrandom shorebird distribution and fine-size variation in prey abundance. Condor 95:94-103. [ Links ]
Collier, K. J. & M. D. Wakeling. 1996. Instream habitat use by blue duck (Hymenolaimus malacorhynchos) in a New Zealand river. Freshwater Biol. 35:277-287. [ Links ]
Costanza, R., W. M Kemp & W. R. Boynton. 1993. Predictability, scala, and biodiversity in coastal and estuarine ecosistems; implications for management. Ambio 22:88-96. [ Links ]
Cupul-Magaña, F. G. 2000. Aves acuáticas del estero El Salado, Puerto Vallarta, Jalisco. Huitzil 1:3-8. [ Links ]
Custer, T. W., R. K. Hines & C. M. Custer. 1996. Nest initiation and clutch size of Great Blue Herons on the Mississippi River in relation to the 1993 flood. Condor 98:181-188. [ Links ]
David, P. G. 1994. Wading bird use of lake Okeechobee relative to fluctuating water levels. Wilson Bull. 106: 719-732. [ Links ]
Davidson, N. C., d ´A. Laffoley, J. P. Doody, L. S. Way, J. Gordon, R. Key, C. M. Drake, M. W. Pienkowski, R. Mitchell & K. L. Duff. 1991. Nature conservation and estuaries of Great Britain. Peterborough, Nature Conservancy Council. [ Links ]
Day, J. W. A. S. Hall., W. M. Kemp & A. Yañez-Arancibia. 1989. Estuarine ecology. Wiley, New York. [ Links ]
Ens, B. J., T. Piersma & R. H. Drent. 1994. The dependence of waders and waterfowl migrating along the East Atlantic Flyway on their coastal food supplies:what is the most profitable research program?. Ophelia Suppl. 6:127-151. [ Links ]
Hayes, F. E. & J. A. Fox. 1991. Seasonality, habitat use, and flock sizes of shoerbirds at the Bahía de Asunción, Paraguay. Wilson Bull. 103:637-649. [ Links ]
Hernández-Vázquez, S. 2000. Avifauna acuática del estero La Manzanilla, Jalisco, México. Acta Zoológica Mexicana, nueva serie. 80:143-153. [ Links ]
Hernández-Vázquez, S.& E. Mellink. 2001. Coastal water- birds of El Chorro and Majahuas, Jalisco, México, during the non-breeding season, 1995-1996. Rev. Biol. Trop.49:357-365. [ Links ]
Howell, S. N. G. & S. Webb. 1995. A guide to the birds of México and Northern Central America. Oxford. New York. 851 pp. [ Links ]
Howes, J. & D. Bakewell. 1989. Shorebird studies manual. Asian Wetland Bureau (ABW) Publication Nº 55. Kuala Lumpur. 362 pp. [ Links ]
Kawaji, N. & S. Shiraishi. 1979. Birds of the North Coast of the sea of Ariake II. The relation between food habits of sandpipers and invertebrates in the substrate. J. Fac. Agr., Kyushu Univ. 23:163-175. [ Links ]
Kramer, G. W. & R. Migoya. 1989. The pacific coast of México. In Smith, L. M., R. L. Pederson & R. M. Kaminski (eds.). Habitat management for migrating and wintering waterfowl in North America.Texas Tech University, Lubbock. pp.507-528. [ Links ]
Schreiber, R. W. & E. A. Schreiber. 1984. Central pacific seabirds and the El Niño Southern Oscillation: 1982 to 1983 perspectives. Science 225:713-716. [ Links ]
Vega-Cendejas, M. E. 1998. Trama trófico de la comunidad nectónica asociada al ecosistema de mangle en el litoral norte de Yucatán. Tesis de Doctorado en Ciencias (Biología). Universidad Nacional Autónoma de México.170 pp. [ Links ]
Warnock, E. S. & J. Y. Takekawa. 1995. Habitat preference of wintering shorebirds in a temporally changing enviroment: Western Sandpipers in the San Francisco Bay estuary. Auk 112: 920-930. [ Links ]
Ysebaert, T., P. L. Meininger, P. Meire, K. Devos, C. M. Berrevoets, R. C. W. Strucker & E. Kuijken. 2000. Waterbird communities along the estuarine salinity gradient of the Schelde estuary, NW-Europe. Biodiversity Conserv. 9:1275-1296. [ Links ]
Zar, J. H. 1974. Biostatistical Analysis. Prentice Hall, New York, 620 p. [ Links ]