










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.53 n.1-2 San José Jun. 2005
Ecología trófica y aspectos reproductivos de Trichomycterus areolatus (Pisces,Trichomycteridae)en ambientes lóticos artificiales
Evelyn Habit 1, Pedro Victoriano 2 & Hugo Campos 3
1 Centro de Ciencias Ambientales EULA -Chile, Unidad de Sistemas Acuáticos, Universidad de Concepción, Concepción, Chile. Barrio Universitario s/n, Concepción, Chile; ehabit@udec.cl, Fax:56-41-731023.
2 Dpto. de Zoología, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile.
3 Instituto de Zoología, Universidad Austral de Chile, Valdivia, Chile.
Recibido 13-III-2003. Corregido 20-X-2003. Aceptado 24-X-2003.
Abstract
Trophic ecology and reproductive aspects of Trichomycterus areolatus (Pisces, Trichomycteridae) in irrigation canal environments. Twelve of the sixteen species of fishes present in the Itata river basin enter the irrigation canals at least once a year, constituting a habitat that is frequently utilized by the native fish assemblages. However, only Trichomycterus areolatus, a native siluriform, is found in great number throughout the entire year, being the dominant species of such artificial environments. Among the factors that could explain the relative success of T. areolatus are its benthonic habits, adaptation to rithral habitat, an offer of trophic resources that is in line with its feeding habits, and a lower abundance of predators in the canals. In this study, the trophic ecology of this species in 174 individuals inhabiting the Itata river (Octava Región, Chile) and 231 from irrigation canals that arise from this river are analyzed and compared. In addition, the length-weight relationship and seasonal frequencies of reproductive status are analyzed, with the purpose of inferring on the status of these populations and their possible condition of resident in these canals. In canals, its diet includes a greater number of different taxa as well as total number of preys consumed in comparison with the river populations, including items of greater biomass, such as lumbriculides. The selectivity analysis showed that T. areolatus prefer prey items like Chironomidae, Baetidae, Elmidae, Plecoptera and Hyallela. Although both the river as well as canal populations show an isometric type growth, they reach greater body lengths and weights in the latter. According to the reproductive states analysis in different periods of the year, populations inhabiting canal environments reproduce synchronically with those of the river, although these latter tend to show a slight delay in the reproductive activity. Rev. Biol. Trop. 53(1-2):195-210. Epub 2005 Jun 24.
Key words: Trichomycterus, feeding ecology, reproduction, irrigation canals, Chile.
La alteración de los regímenes de caudal de los sistemas fluviales provocan cambios importantes en la estructura de los ensambles de peces (Travnichek y Maceina 1994). Por lo general, se produce una disminución de la complejidad de tales agrupaciones, aunque en algunas situaciones se generan condiciones favorables para ciertas especies, dependiendo de su biología y plasticidad (Del Mar Torralva et al. 1997, Araoye 2002, Smith et al. 2003). Un tipo de alteración frecuente es la desviación y canalización del agua con fines de riego, desde el curso natural de ríos hacia canales que constituyen ambientes lóticos artificiales. Los canales de regadío conforman hábitats temporales y de fuertes perturbaciones para la mayor parte de la ictiofauna fluvial (Redding y Midlen 1992), pero a pesar de sus drásticas fluctuaciones de caudal, algunas especies persisten en ellos, representando ambientes factibles de ser colonizados por parte de la fauna íctica (Habit 1994a, 1997).
Un estudio realizado en la zona centro sur de Chile (cuenca del río Itata, 36° 12 -37° 16 S; 71° 00 -73° 10 W) muestra que 12 de las 16 especies de peces presentes en la cuenca ingresan al menos en una época del año a los canales de riego, faltando sólo aquellas que son propias del área potamal del río (Habit 1994a, Habit y Parra 2001). La mayoría de las que ingresan disminuyen drásticamente sus abundancias en períodos del cierre de canales (Habit 1997).
A diferencia de éstas, el siluriforme nativo Trichomycterus areolatus (Valenciennes, 1848) domina y persiste durante todo el año en estos ambientes artificiales (Habit 1994a). Entre los factores que pueden explicar la persistencia de las poblaciones en ambientes artificiales cuenta la concordancia entre los hábitos alimentarios y una oferta adecuada de recursos tróficos. Esto se puede reflejar tanto en altos valores del factor de condición como en la capacidad de estas poblaciones de desarrollar su ciclo reproductivo completo en este tipo de hábitat.
Las especies del género Trichomycterus son de amplia distribución en el neotrópico (de Pinna 1998, Román-Valencia 2001), estando representado en Chile por cinco especies (T. areolatus, T. chiltoni, T. rivulatus, T. chungaraensis y T. laucaensis, Arratia 1990, Dyer 2000). La especie T. areolatus es un bagre frecuente en ambientes ritrales de los ríos chilenos, distribuyéndose desde Illapel por el norte (31º S) hasta Llanquihue por el sur (41º S), (Arratia et al. 1981). En la Región del Biobío (zona centro-sur de Chile) ha sido reportado para los principales cursos fluviales, como son los ríos Biobío (Campos et al.1993a, 1993b), Andalién (Ruiz 1994), Laja (Ruiz 1996) e Itata (Habit 1994b, 1998). Si bien está considerada como una especie vulnerable a nivel nacional (Campos et al.1998), en la mayoría de los ríos mencionados presenta altas abundancias en sectores de bolones y de aguas someras, los que constituyen su hábitat preferencial (Arratia 1983, Campos 1985, Campos et al. 1993a, 1993b, Habit 1994b, 1998, Ruiz 1994, 1996).
Para ambientes lóticos naturales T. areolatus ha sido descrita como una especie bentófaga, que depreda exclusivamente insectos (Arellano et al. 1983, Campos et al. 1993a, Ruiz et al. 1993, Ruiz y Berra 1994) o que se comporta como un meso depredador carnívoro (sensu Welcomme 1979) en sistemas como el río Maipo (Duarte et al. 1971). Es decir, T. areolatus presenta cierta plasticidad trófica, ampliando su nicho trófico en determinados sistemas. En canales de riego, Habit et al. (1998) informaron la existencia de una importante oferta de fauna zoobentónica, lo que sugiere la presencia de condiciones favorables para el mantenimiento de poblaciones locales de este pez. En cuanto a su reproducción, si bien trabajos previos indican que ésta ocurre en los meses de primavera-verano (Manríquez et al. 1988, Campos et al. 1993b, Ruiz et al. 1993), se desconoce si las poblaciones locales que utilizan los canales de riego se reproducen en ellos y en los mismos períodos del año, constituyendo un hábitat adecuado para mantener poblaciones residentes de esta especie. El objetivo de este trabajo es analizar los hábitos alimentarios, reproducción y condición de las poblaciones locales de T. areolatus en canales de riego de la cuenca del río Itata, comparando con situaciones de cauces naturales y así inferir acerca de las condiciones que explican la persistencia de T. areolatus en ambientes artificiales. Se propone la hipótesis de que la dominancia de esta especie en los canales indica que establece poblaciones en buen estado, lo cual se refleja en mayores factores de condición respecto del río. Además, se postula que en los canales de riego se presenta una mayor amplitud de nicho trófico, ya que consumiría en ellos no sólo insectos sino otras presas, tales como moluscos y anélidos, más abundantes en los canales (Habit et al. 1998). Finalmente, se postula que T. areolatus desarrolla todo su ciclo de vida en los canales de riego, es decir es un residente de estos ambientes artificiales.
Materiales y métodos
Se realizaron cuatro muestreos en nueve estaciones en la cuenca del río Itata (tres estaciones de río y seis de canales asociados, Fig. 1), durante marzo (verano), mayo (otoño), septiembre (invierno) y diciembre (primavera) de 1994. Tres estaciones se ubicaron en un cauce fluvial natural, y seis en canales artificiales de riego (Fig.1). Las estaciones 1 (río Huépil: rápidos del ritrón), 2 (río Itata: pozas del ritrón) y 3 (río Itata: potamón) se ubicaron en el tramo inmediatamente aguas arriba de la bocatoma de los canales de riego estudiados. Las estaciones 4, 6 y 8 se ubicaron en la bocatoma (u origen) de los canales Cruz del Ciprés (0.5 m3 /s), Zañartu (20 m3 /s) y Quillón (2 m3 /s y revestimiento de concreto), respectivamente. Las estaciones 5, 7 y 9 correspondieron a sitios de muestreo ubicados al interior de cada uno de los mismos canales de riego. Las característica de los canales han sido descritas previamente (Habit 1994a). Debido al bajo número de ejemplares detectados en las estaciones 8 y 9, éstas no fueron consideradas en algunos de los análisis cuantitativos. Dichos sitios se diferencian por presentar características que dificultan el muestreo, principalmente por su alta velocidad de corriente y recubrimiento de concreto del canal.
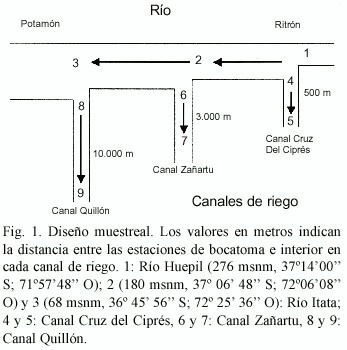
La recolección de peces en el río y canales se efectuó mediante un equipo de pesca eléctrica Elektro-Fischfangerat, motor JLO a gasolina y una red cónica de 5 mm de trama y 1 m de diámetro. Las capturas se realizaron en la mañana, dado que T. areolatus es un depredador preferentemente nocturno (Campos obs. pers.), recorriendo áreas de 50 a 100 m2 . Cada individuo fue medido (longitud total; 0,01 mm de precisión), pesado (0,01 g de precisión), fijado en terreno en formalina al 10 % y depositados en frascos debidamente rotulados.
Con el fin de tener una muestra representativa de ejemplares de todas las estaciones de muestreo, se analizó la composición del contenido estomacal de un total de 405 individuos (174 de río y 231 de canales) mediante los métodos de frecuencia de presencia y numérico. La frecuencia de presencia se refiere al porcentaje de estómagos que contienen la presa en cuestión, mientras la composición numérica corresponde al porcentaje de individuos de la presa en cuestión, respecto al total de presas contabilizadas. El aporte relativo de cada ítem a la alimentación se estimó utilizando el Indice de Importancia Numérica (IIN) (Windell 1968):

A partir de las abundancias relativas de los diferentes tipos de presa, se estimó la amplitud de nicho para cada estación de muestreo, a través del índice de Simpson (Magurran 1988), el cual considera las proporciones de abundancia de cada categoría de presas. Esta estimación no se aplicó a la estación de muestreo 9 dado el bajo número de ejemplares analizados para esta localidad. Con el fin de determinar diferencias de niveles de amplitud de nicho entre ambientes de canal y de río, se aplicó una prueba no paramétrica de diferencia de medias (Mann Whitney-U).
Para determinar el grado de diferenciación dietaria, tanto entre estaciones de muestreo como entre períodos del año, se aplicó un análisis de varianza no paramétrico de Kruskall Wallis, considerando como variables dependientes las proporciones de abundancias (log x+1) de las presas, la amplitud de nicho y el número total de presas consumidas (log x+1).
Para estimar la relación entre la disponibilidad de presas y la composición dietaria, es decir, para determinar si las presas se encontraban sobre o sub representadas en la dieta, en proporción a sus abundancias relativas en el ambiente, se estimó el coeficiente de Selectividad (E*) (Vanderploeg y Scavia 1979, en Parker 1994). Para este análisis no se consideraron los ejemplares provenientes del canal Quillón (estaciones 8 y 9), debido a que no se cuenta con información de abundancia de bentos en este sistema (Habit et al.1998). Este índice se obtiene mediante la siguiente fórmula:
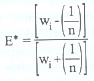
con,
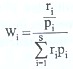
donde, ri es la abundancia relativa de la presa en la dieta, pi es la abundancia relativa de la presa en el bentos y n es el número de táxones involucrados en análisis. El índice E* varía entre -1 y +1, indicando sub o sobrerepresentación en la dieta, respectivamente. Valores cercanos a cero (-0.05 a +0.05)indican selección neutra. Considerando que su valor puede ser sesgado por errores de muestreo, sólo se incluyen para el cálculo las presas con abundancias relativas mayores al 1 % tanto en la dieta como en el ambiente. Los muestreos de zoobentos para estimar oferta alimentaria se realizaron simultáneamente a los de peces, y para los análisis correspondientes, se consideraron los datos informados por Habit et al. (1998).
Se determinó el grado de madurez sexual de 345 ejemplares de acuerdo a la nomenclatura de Ricker (1968) y utilizada en peces fluviales chilenos por Ruiz et al. (1993) y Gavilán (1993), reconociendo los estados reproductivos I (virgen) al VIII (reposo). La información obtenida fue analizada agrupando los individuos de acuerdo a su procedencia (río o canal de riego) y época de muestreo (otoño –invierno y primavera -verano). Para cada grupo se estimó la frecuencia de individuos en cada uno de los ocho estados reproductivos reconocidos y el promedio de longitud total de ellos. Se comparó la distribución de frecuencia de individuos adultos para cada uno de los ocho estados de reproducción entre los ambientes de río y canal utilizando una prueba de Kolmogorov-Smirnov para dos muestras.
Los datos de longitud y peso (591 ejemplares, 240 de río y 351 de canales) fueron ajustados a un modelo de regresión multiplicativo. Se obtuvo las ecuaciones de regresión y se compararon estadísticamente los coeficientes de alometría (o pendiente) entre río y canales de riego mediante una prueba t de Student (Zar 1984), agrupando los datos de los cuatro muestreos. Para el cálculo del factor de condición o corpulencia (K) se utilizó la fórmula de Beckmann (1948), la cual
corresponde a:
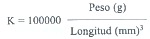
Se comparó la longitud total, peso y factor de condición entre los ambientes de río y canal agrupando la información de los cuatro muestreos para un total de 600 individuos (240 provenientes del río y 360 de canales de riego), mediante una prueba de t de Student.
Para determinar el efecto de la estacionalidad (época del año) y estación de muestreo sobre la longitud total, peso y factor de condición (K), se realizaron análisis de varianza de dos vías (ANOVA) y se utilizó el método de comparaciones múltiples de los promedios mediante la prueba de Tukey.
Resultados
Dieta: de un total de 405 estómagos analizados (174 individuos provenientes del río y 231 de canales de riego),el 27.5% de ellos se encontró vacío. De estos 111 individuos, 67 correspondieron a ejemplares capturados en canales de riego y 44 a estaciones ubicadas en el río.
La dieta de T. areolatus estuvo constituida en total por 21 táxones animales (Cuadro 1), todos presentes en individuos de canal, a diferencia de los de río que presentaron sólo un total de 15 tipos de presa. La mayor parte de los ítemes correspondió al grupo de los insectos, principalmente, en estados inmaduros, los otros tres phyla representados fueron Crustacea, Annelida y Mollusca.

Entre los canales, los ejemplares de las estaciones 6 y 7 (canal Zañartu) presentaron la mayor riqueza de ítemes alimentarios en todas las épocas de muestreo (Cuadro 2). En tanto, en el río, los individuos del ritrón tendieron a presentar una mayor variedad de presas, con excepción de invierno. El ítem básico de la dieta de T. areolatus correspondió a larvas de la familia Chironomidae, las que mostraron altos valores de IIN en las cuatro épocas del año y en todas las estaciones de muestreo. De los restantes táxones encontrados, destacaron en menor grado los efemerópteros, plecópteros y coleópteros de la familia Elmidae. Los táxones menos frecuentes resultaron ser los crustáceos Samastacus spinifrons y Daphnia sp., solamente presentes en el muestreo de invierno en ejemplares provenientes de la estación 8.S. spinifrons correspondió a un ejemplar juvenil, consumido durante su período de muda. De igual forma, el gastrópodo Gundlachia gayana sólo fue encontrado en un ejemplar capturado en la estación 7 durante el verano. Las ninfas de Lepthophlebiidae, las larvas de Limnephilidae y las larvas de Gripopterygidae fueron consumidas preferentemente en el río, mientras que sus abundancias en los estómagos de ejemplares de canales fue muy baja. A diferencia del caso anterior, los oligoquetos lumbricúlidos resultaron con mayor representación en las estaciones de muestreo ubicadas en los canales de riego, siendo Lumbriculus exclusivo de la dieta de canales (Cuadro 2). Por otro lado, en la estación 3 (potamón) se evidenció una mayor presencia de tricópteros, plecópteros y coleópteros.
De acuerdo al análisis de varianza, el único efecto significativo (p <.05) fue atribuible a las estaciones de muestreo para la variable dependiente número total de presas consumidas, el que fue significativamente más alto en las estaciones 2,6 y 7 (río Itata en Cholguán y canal Zañartu). Dado que este análisis no entregó diferencias significativas entre épocas del año, se procedió a agrupar los datos de cada localidad, y los análisis posteriores sólo se refieren a comparaciones entre estaciones de muestreo.
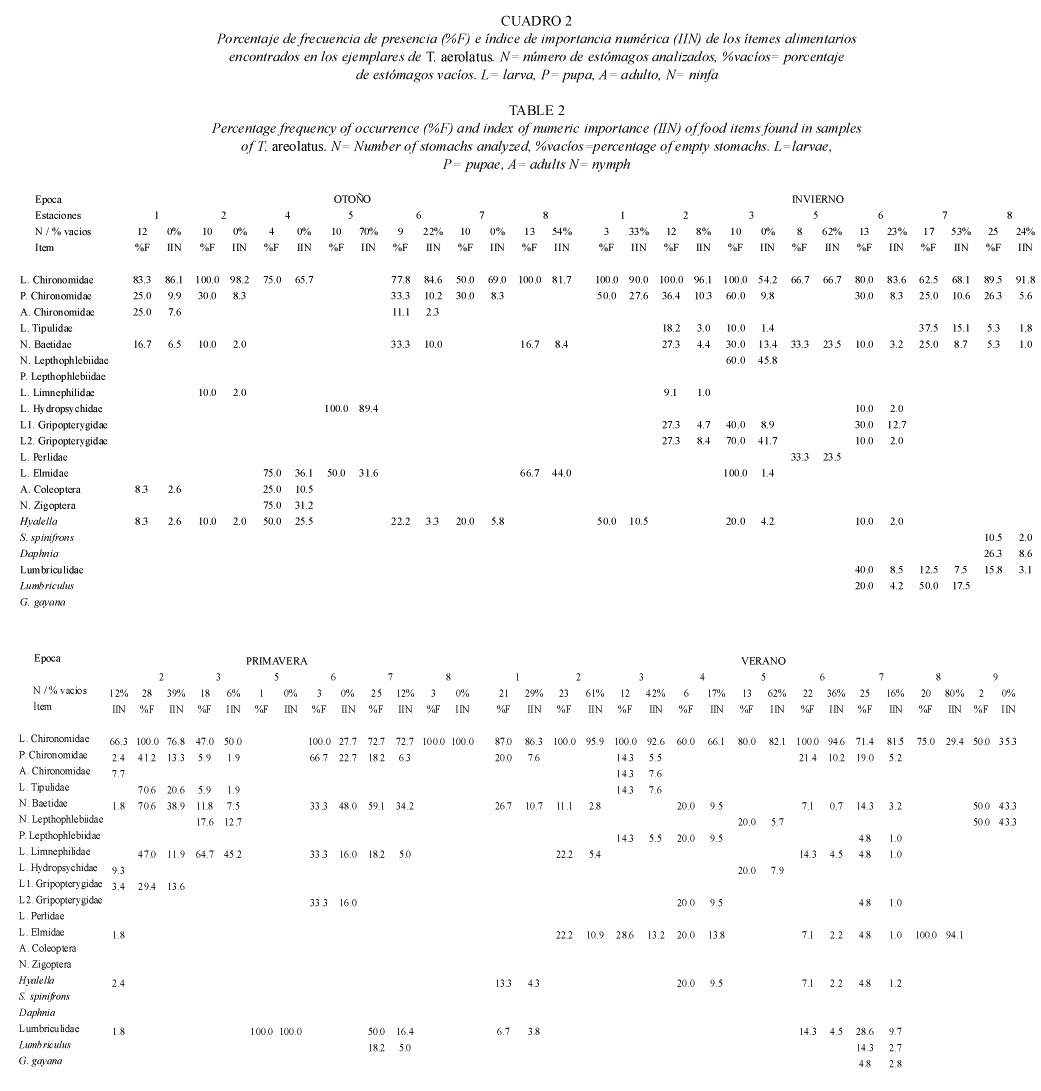
El análisis de preferencia de presas incluyó siete táxones que estuvieron representadas sobre un 1 %, tanto en el bentos como en la dieta. De éstas, Chironomidae, Baetidae, Elmidae, Hyalella y Plecoptera presentaron selectividades cercanas a +1, es decir, se encuentran sobre representadas en la dieta, a excepción de Hyalella y Plecoptera en la estación 7, donde presentan selectividades negativas (Fig. 2). Lumbriculidae y Trichoptera, presentaron mayor heterogeneidad en las distintas estaciones de muestreo, presentando índices de selectividad igual a -1 en las estaciones 2 y 4, y 3 y 4, respectivamente. Ello sugiere un consumo proporcional a los valores de oferta.
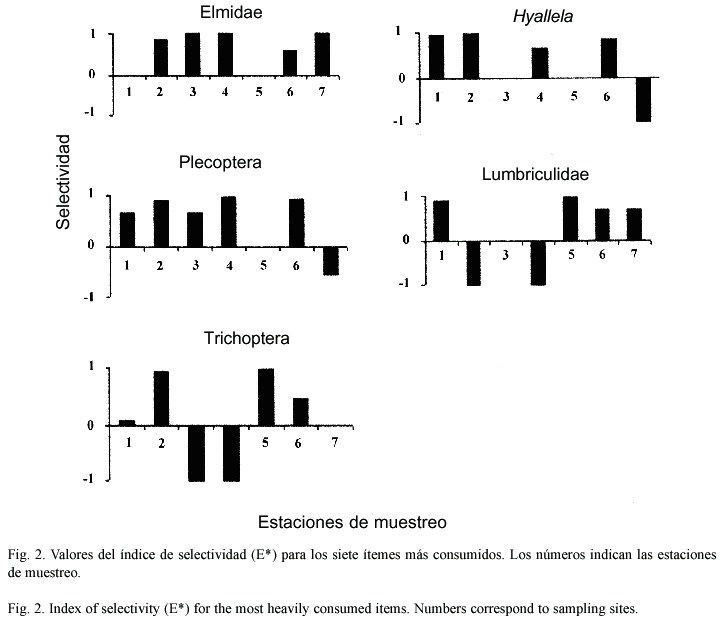
En cuanto a la amplitud de nicho la prueba de Kruskall Wallis no entregó diferencias significativas entre los peces de ambientes del cauce natural y los de ambientes de canal, aún cuando los primeros muestran una mayor varianza (Fig.3).
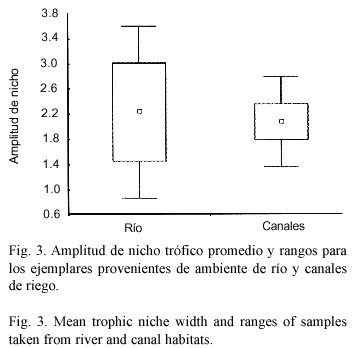
Reproducción: los individuos menores a 49.9 mm de longitud total (L.T.) correspondieron a juveniles, dado que ejemplares de este tamaño no mostraron actividad reproductiva. Estos fueron el 42.3% del total de individuos estudiados, de los cuales el 67.1% fueron capturados en la época de primavera -verano, proviniendo el 53% de canales y el 47% del río. Los ejemplares adultos capturados durante otoño -invierno, tanto en el río como en canales (machos y hembras) solo alcanzaron hasta estado reproductivo IV (en fase de desarrollo), mientras que en los meses estivales (primavera -verano) los peces presentaron estados reproductivos avanzados (V o grávidos a VIII o en reposo), (Fig.4 y 5). Los estados reproductivos avanzados fueron más frecuentes en ejemplares de tallas corporales mayores.
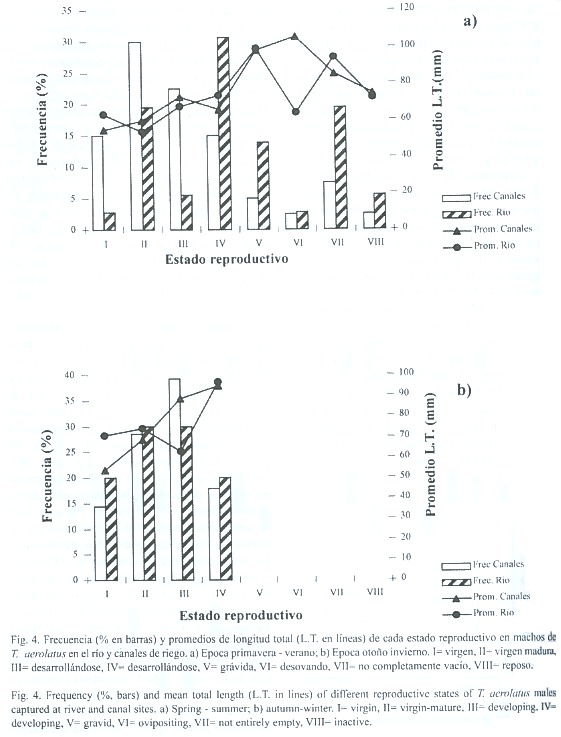
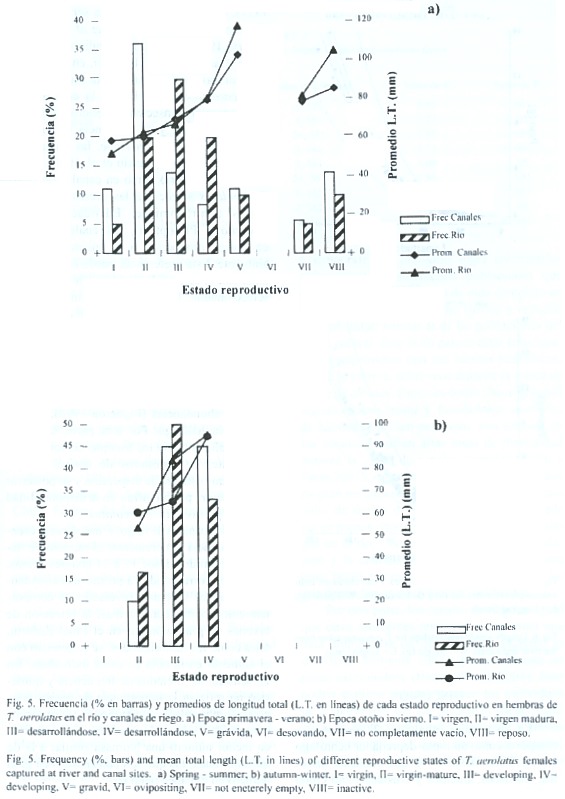
La tendencia general indica que en ambos tipos de ambientes se presentaron los mismos estados reproductivos en épocas similares. Sin embargo, la distribución de frecuencias de estados reproductivos fue significativamente diferente entre río y canales durante la época de primavera -verano (X2 =14.64,p <.05,7 g.l.). Durante dicho período, en el que se presentaron todos los estados, los canales mostraron para ambos sexos una mayor frecuencia de estados inmaduros respecto a las detectadas en el río (Fig.4a y 5a), siendo la mayor diferencia la mayor proporción de individuos en estado IV en el río respecto de los canales de riego Esto sugiere que si bien la reproducción es sincrónica en términos generales, las poblaciones de canal presentan un leve retardo respecto a las de cauce natural. En el período otoño –invierno no se detectaron diferencias significativas entre ambos ambientes.
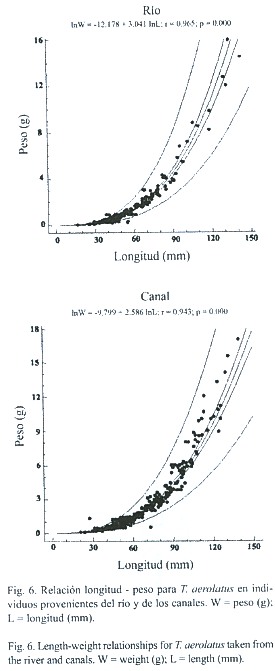
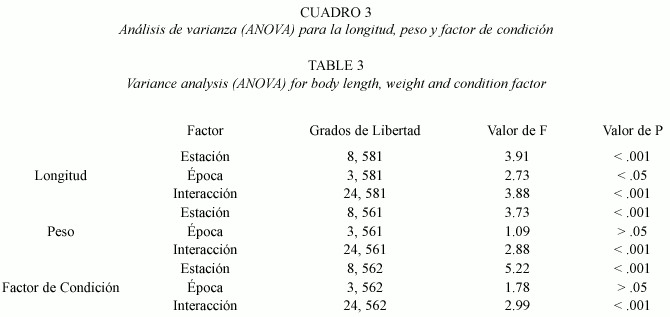
Longitud, peso y factor de condición: el intervalo de longitudes de los ejemplares capturados en estaciones de río fue igual a 16.7 –152.1 mm L.T. y 0.05 –16.01 g de peso. En los canales fue igual a 20.4 –150.2 mm de L.T. y 0.10 –17.20 g de peso. Los individuos que habitan en los canales de riego fueron significativamente más largos (t =-4.57;598 gl., p <.001) y pesados (t =-2.57, p <.001, 598 g.l.) que aquellos del río, sin embargo, no presentaron diferencias estadísticas en el factor de condición (t =-1.38,p >.05,598 g.l.). Esto se corroboró mediante la comparación de las curvas de regresión entre ambas poblaciones, las que igualmente no mostraron diferencias significativas en sus coeficientes de alometría (t =2.93,p >.05,588 g.l.), (Fig.6). En ambos casos se observó un crecimiento de tipo isométrico positivo. Al analizar mediante una Anova las diferencias de longitud, peso y factor de condición, tanto entre sitios como épocas de muestreo, sólo se detectó variación espacial en el peso y factor de condición (Cuadro 3). En tanto, la longitud mostró diferencias significativas tanto para la época como para la estación de muestreo. En todos los casos, existió interacción significativa entre los factores época y estación de muestreo.
Discusión
En los canales estudiados T. areolatus se comporta como un meso-depredador bentófago (sensu Welcomme 1979), que consume insectos, moluscos, anélidos y crustáceos, tal como en las poblaciones del río Maipo (Duarte et al. 1971). En el río Itata, tiende a ser insectívoro como en el río Biobío (Ruiz et al. 1993, Ruiz y Berra 1994) y en los ríos Andinos del sur de Chile (Campos 1985). Es decir, en los canales T. areolatus, presenta una dieta muy similar a T. itacarambiensis de Brasil, la cual depreda preferentemente insectos y anélidos autóctonos (Trajano 1997a), y se diferencia de especies como T. knerki y T. meridae, las cuales sólo consumen insectos (Baskin et al. 1980).
Tanto en el río como en canales de riego, T. areolatus selecciona las presas más abundantes (Chironomidae, Baetidae, Elmidae, Hyalella y Plecoptera), los cuales consume en mayor proporción a la encontrada en el ambiente. Esta preferencia puede estar relacionada con la facilidad de captura de los táxones seleccionados. En general, los invertebrados seleccionados son móviles y viven asociados al sustrato, a diferencia de los lumbricúlidos y tricópteros (de selectividad neutra), los cuales son organismos menos móviles. Los lumbricúlidos habitan enterrados en sustratos fangosos y sólo están expuestos cuando se encuentran en grandes abundancias (Figueroa 1998), lo cual facilita su hallazgo. Por otra parte, tanto los lumbricúlidos como los tricópteros, son menos abundantes en muestreos de deriva, lo que sugiere que esta vía de dispersión y su posterior depositación, podría influir en la disponibilidad de estas presas para T. areolatus.
En los canales de riego, T. areolatus presentó la tendencia a incrementar el número de ítemes consumidos desde 15 a 21 táxones-presas, los cuales corresponden a presas más abundantes en dichos sistemas, revelando un comportamiento oportunista. Además, la selección de táxones de gran biomasa en el canal Zañartu, tales como los lumbricúlidos, se relacionan con el aumento en la talla de estos individuos. En tal sentido, los métodos de frecuencia y numérico no reflejan la importancia de ítemes tales como lumbricúlidos, gastrópodos y crustáceos consumidos sólo en los canales. Estos aportan en menor número una biomasa similar a la de quironómidos, los que son consumidos en gran número en el cauce natural.
La reproducción de T. areolatus es estacional, presentando su actividad reproductiva a finales de la época de lluvias o comienzos de primavera, tanto en los cauces artificiales como en los naturales. La reproducción de las poblaciones estudiadas es sincrónica con aquellas descritas para los ríos Angostura, Copequén (Arellano et al.1980) y Andalién (Ruiz 1994). La época de máximo reproductivo comprende entre los meses de septiembre -noviembre, encontrándose juveniles (<49,9 mm) a partir del mes de octubre. Esta cronología es coherente con el ciclo ovárico descrito por Manríquez y Arellano (1981) y Manríquez et al. (1988) en poblaciones de Angostura y Copequén en Chile. El leve desfase encontrado en las frecuencias de estados reproductivos entre ambos ambientes, sugiere que existe algún factor no determinado en este estudio, que retarda el inicio del período de actividad reproductiva en las poblaciones de ambientes de canales.
Los canales de riego presentan una gran similitud estructural con sistemas fluviales canalizados, en los cuales aumenta la velocidad del agua y se pierde la secuencia de rápidos y pozas. De esta forma en los canales se pierde la gradualidad de los sistemas fluviales naturales hacia sectores potamales, por lo que constituyen extensas zonas de rápidos (Brookes 1994, Meyer y Molnár 1994), los que resultan un hábitat propicio para T. areolatus. Aún cuando los canales de riego son ambientes inestables y temporales, T. areolatus logra colonizarlos exitosamente, manteniendo poblaciones que pueden desarrollar su ciclo de vida completo en ellos, donde ocupa un nicho trófico y espacial (microhábitat) similar al de las poblaciones del cauce natural. Este éxito parece estar relacionado principalmente con sus hábitos bentónicos, lo que le permite sobrevivir durante la época de cierre de canales. Especies como Oncorhynchus mykiss, Salmo trutta y Basilichthys australis, de hábitos más bien pelágicos, son escasos en los canales o sufren altas tasas de mortalidad durante la época de canales cerrados (Habit y Parra 2001). Además, los salmónidos dependen en gran medida de los ritmos diarios y estacionales de deriva de presas, proceso que puede ser de menor importancia en canales de regadío que en el río. Con esto, el tiempo de alimentación y la cantidad de presas disponibles para estos peces son más restringidos respecto a los que dependen exclusivamente del bentos.
Por otra parte, los canales de riego, al igual que otros ambientes temporales presentan una baja abundancia de depredadores (Katano et al. 1998, Williams 1987), principalmente de peces salmonídeos (Habit y Parra 2001). Esto podría explicar porqué, aunque los individuos de T. areolatus del río presentan dietas similares, se reproducen sincrónicamente y crecen isométricamente, al igual que en los canales, estos últimos alcanzan mayores tallas y pesos que los primeros.
T. areolatus parece presentar una extraordinaria capacidad colonizadora, siendo la primera especie nativa en ocupar hábitat ritrales fuertemente perturbados, como en el caso del río Laja (cuenca del Biobío) después del desvío de su cauce (datos no publicados). Es probable que T. areolatus presenta una buena capacidad locomotora, tal como la especie congenérica, T. itacarambiensis, la cual alcanza desplazamientos activos continuos de hasta 600 m y desplazamientos por efecto de deriva (aguas abajo) de hasta 1500 m (Trajano 1997b).
Los resultados de este estudio permiten concluir que la conducta oportunista en la estrategia alimentaria de T. areolatus ,sus hábitos bentónicos y adaptación a ambientes de rápidos, les permite una efectiva colonización de los canales de riego, los que dada su alta heterogeneidad ambiental (velocidad de la corriente, tipo de sustrato, vegetación), resultante de una escasa mantención y su construcción sin revestimiento, permite la formación de hábitat propicios para la colonización de una abundante fauna de macroinvertebrados que sustentan estas poblaciones de peces nativos.
Agradecimientos
Agradecemos a Raúl Arriagada, Cesar Cid y Pedro Sánchez por su valiosa ayuda en actividades de terreno. A Viviana Olmos por sus observaciones al manuscrito. Este proyecto
fue financiado por el Proyecto FONDECYT 2940040 y parcialmente por el proyecto DIUC 200.113.054-1.3.
Resumen
Doce de las dieciséis especies de peces presentes en la cuenca del río Itata ingresan al menos en una época del año a los canales de riego, constituyendo un hábitat frecuentemente utilizado por la ictiofauna nativa. Sin embargo sólo el siluriforme Trichomycterus areolatus se encuentra en altas abundancias todo el año, siendo la especie dominante de estos ambientes fluviales artificiales. Entre los factores que pueden dar cuenta de su éxito relativo dentro de los canales están sus hábitos bentónicos y adaptación a ambientes ritrales, una oferta de recursos tróficos concordante con sus hábitos alimentarios y una menor abundancia de depredadores. En este estudio se analiza comparativamente la ecología trófica de esta especie en 174 individuos que habitan en el río Itata (Octava Región de Chile) y 231 de canales de riego que nacen de este sistema fluvial. Además se analizan las relaciones de talla-peso y frecuencia estacional de estados reproductivos, con el fin de inferir acerca del estado de las poblaciones y su condición de residentes en canales de riego. La dieta de las poblaciones de canales incluye una mayor riqueza de táxones y una mayor cantidad de presas consumidas respecto a las del curso natural. El análisis de selectividad muestra que T. areolatus prefiere, tanto en el río como en los canales de riego,presas tales como Chironomidae, Baetidae, Elmidae, Plecoptera y Hyallela. Además, en los canales de riego prefiere presas de mayor biomasa como los lumbricúlidos. Si bien las poblaciones locales de canales presentan, al igual que los ejemplares de río, un crecimiento de tipo isométrico, éstos alcanzan mayores tallas y pesos corporales que los individuos de los cauces naturales. De acuerdo al análisis de estados reproductivos de 345 ejemplares en diferentes períodos del año, las poblaciones de ambientes de canal se reproducen sincrónicamente con las del río, aunque con un leve retardo en la actividad reproductiva.
Palabras clave: Trichomycterus, ecología de alimentación, reproducción, Itata river, canales de irrigación, Chile.
Referencias
Araoye. P. A. 2002. Man-made lakes, ecological studies and conservation needs in Nigeria. Rev. Biol. Trop. 50:857-864. [ Links ]
Arellano, M., L. Huaquín & A. Manríquez. 1983. Aspectos comparativos de dos especies de Siluriformes Chilenos de agua dulce (Pisces, Trichomycteridae). Arch. Biol. Med. Exp. 16:136. [ Links ]
Arellano, M., L. Huaquín, A. Manríquez & G. Arratia. 1980. Aspectos reproductivos en Trichomycterus aerolatus. Arch. Biol. Med. Exp. 13:43. [ Links ]
Arratia, G. 1983. Preferencias de hábitat de peces siluriformes de aguas continentales de Chile (Familia Diplomystidae y Trichomycteridae). Stud. Neotr. Fauna Environ. 18:217-237. [ Links ]
Arratia, G. 1990. The south american Trichomycterinae (Teleostei:Siluriformes), a problematic group. p. 395- 403. In G. Peters & R. Hutterer (eds.). Vertebrates in the tropics. Museum Alexander Koenig, Bonn. [ Links ]
Arratia, G., G. Rojas & A. Chang. 1981. Géneros de Peces de Aguas Continentales de Chile. Publ. Ocas. Mus. Nac. Hist. Nat. 34:3-108. [ Links ]
Baskin, J. N., T. M. Zaret & F. Mago-Leccia. 1980. Feeding of reportedly parasitic catfiches (Trichomycteridae and Cetopsidae) in the Rio Portuguesa Basin, Venezuela. Biotropica 12:182-186. [ Links ]
Beckmann, W. 1948. The lenght -weight relationship, factors for conversions between standard and total lengths, and coefficient of condition for seven Michigan fishes. Trans. Am. Fish. Soc. 75:237-256. [ Links ]
Brookes, A. 1994. River channel change, p. 55-75. In P. Callow & G. Petts (eds). The River Handbook: Hydrological and Ecological Principles. Vol II. Blackwell Science Oxford. [ Links ]
Campos, H. 1985. Distribution of the fishes in the Andean rivers in the South of Chile. Arch. Hydrobiol. 104: 169-191. [ Links ]
Campos, H., V. H. Ruiz, J. F. Gavilán & F. Alay. 1993a. Peces del Río Biobío. Serie Publ. Divulgación EULA 5:1-100. [ Links ]
Campos, H., F. Alay, V. H. Ruíz & J. F. Gavilán. 1993b. Antecedentes biológicos de la fauna íctica presente en la hoya hidrográfica del río Bíobio, p. 70-72. In O. Parra & F. Faranda (eds). Seminario Limnología y Evaluación de Impacto Ambiental. Centro EULA –Chile, Concepción, Chile. [ Links ]
Campos, H., G. Dazarola, B. Dyer, L. Fuentes, J. F. Gavilán, L. Huaquín, G. Martínez, R. Meléndez, G. Pequeño, F. Ponce, V. H. Ruiz, W. Siefeld, D. Soto, R. Vega & I. Vila. 1998. Categorías de Conservación de peces nativos de aguas continentales de Chile. Bol. Mus. Nac. Hist. Nat. 47:101-122. [ Links ]
de Pinna, M. C. 1998. Phylogenetic relationships of neotropical siluriforms (Teleostei: Ostariophysi): historical overview and synthesis of hypotheses. p. 279-330. In L.R. Malabarba, R. E. Vari, Z. M. Lucena & C. A. Lucena (eds.). Phylogeny and classification of neotropical fishes. Porto Alegre, Edipucrs. [ Links ]
Dyer, B. 2000. Systematic review and biogeography of the freshwater fishes of Chile. Estudios Oceanológicos 19:77-98. [ Links ]
Del Mar Torralva, M., M. A. Puig & C. Fernández-Delgado. 1997. Effect of river regulation on the life-history patterns of Barbus sclateri in the Segura river basin (south-east Spain). J. Fish Biol. 51:300-311. [ Links ]
Duarte, W., R. Feito, C. Jara, C. Moreno & A. E. Orellana. 1971. Ictiofauna del sistema hidrográfico del río Maipo. Bol. Mus. Nac. Hist. Nat. Chile 32: 227-268. [ Links ]
Figueroa, R. 1998. Calidad de agua y macroinvertebrados bentónicos: Río Damas, Osorno, un caso de estudio. Seminario Experimental Magister en Ciencias Zoología Universidad de Concepción, Concepción. 70 p. [ Links ]
Gavilán, J. F. 1993. Estudio biológico de poblaciones de Percichthys trucha (Percichthyidae)y Oncorhynchus mykiss (Salmonidae) presentes en la hoya hidrográfica del río Biobío, Chile. Tesis de doctorado, Universidad de Concepción, Concepción. 285 p. [ Links ]
Habit, E. 1994a. Ictiofauna en canales de riego de la cuenca del río Itata durante la época de otoño -invierno. Com. Mus. Hist. Nat. Concepción 8:7-12. [ Links ]
Habit, E. 1994b. Contribución al conocimiento de la fauna íctica del río Itata. Bol. Soc. Biol. Concepción 65: 143-147. [ Links ]
Habit, E. 1997. Ictiofauna de un sistema fluvial modificado por canales de riego. Comparación de las comunidades de peces asociadas al cauce natural y canales artificiales. Tesis de doctorado, Universidad de Concepción, Concepción. 193 p. [ Links ]
Habit, E. 1998. Peces, p. 79 -86. In O. Parra & E, Habit (eds.).Documento de Síntesis Estudio de Línea de Base para la Evaluación de Impacto Ambiental del Complejo Forestal Industrial Itata. Centro EULA - Chile Universidad de Concepción. 172 p. [ Links ]
Habit, E., C. Bertrán, S. Arévalo & P. Victoriano. 1998. Benthonic fauna of Itata River and irrigation canals (Chile).Irrigation Science 18:91-99. [ Links ]
Habit, E. & O. Parra. 2001. Impacto ambiental de los canales de riego sobre la fauna de peces. Ambiente y Desarrollo XVII:50-56. [ Links ]
Katano, O., J. Toi, K. Maekawa & K. Iguchi. 1998. Colonization of an artificial stream by fishes and aquatic macro-invertebrates. Ecological Research 13:83 -96. [ Links ]
Magurran, A. E. 1988. Ecological Diversity and its Measurement. Croom Helm Londres. 167 p. [ Links ]
Manríquez, A. & M. Arellano. 1981. Ciclo reproductivo y ovárico en Trichomycterus areolatus. Arch. Biol. Méd. Exp. 14:276. [ Links ]
Manríquez, A., L. Huaquín, M. Arellano & G. Arratia. 1988. Aspectos reproductivos de Trichomycterus areolatus Valenciennes, 1846 (Pisces: Teleostei: Siluriformes) en río Angostura, Chile. Stud. Neot. Fauna Environ. 23:89-102. [ Links ]
Meyer, C. & P. Molnár. 1994. Environmental Impacts of Channelisation Projects. Seminario CE 624 Control de Crecidas e Inundaciones. Universidad de Michigan: 1-15 p. [ Links ]
Parker, M. 1994. Feeding ecology of stream –dwelling pacific giant salamander larvae (Dicamptodon tenebrosus). Copeia 3:705-718. [ Links ]
Redding, T. A. & A. Midlen. 1992. Estudio de la producción piscícola en los canales de riego. FAO Doc. Téc. Pesca N º317.114 p. [ Links ]
Ricker, W. E. (ed.). 1968. Methods for Assessment of Fish Production in Fresh Waters. IBP Handbook N °3 Int. Biol. Prog.7 Marylebone Road London NW1 Blackwell Scientific Publications. 313 p. [ Links ]
Roman-Valencia, C. 2001. Ecología trófica y reproductiva de Trichomycterus caliense y Astroblepus cyclops (Pisces:Siluriformes) en el río Quindio,Alto Cauca, Colombia. Rev. Biol. Trop. 49:657-666. [ Links ]
Ruiz, V. H. 1994. Ictiofauna del río Andalién (Concepción, Chile). Gayana Zool. 57:109-278. [ Links ]
Ruiz, V. H. 1996. Ictiofauna del río Laja (VIII Región, Chile): una evaluación preliminar. Bol. Soc. Biol. Concepción 67:15-21. [ Links ]
Ruiz, V. H., M. T. López, H. I. Moyano & M. Marchant. 1993. Ictiología del Alto Biobío: aspectos taxonómicos, alimentarios, reproductivos y ecológicos con una discusión sobre la hoya. Gayana Zool. 57:77-88. [ Links ]
Ruiz, V. H. & T. Berra. 1994. Fishes of the high Biobío river of south-central Chile with notes on diet and speculations on the origin of the ichthyofauna. Ichthyol. Exploration Freshwaters 5:5-18. [ Links ]
Smith, W. S., M. Petrere Jr. & W. Barrella. 2003. The fish fauna in tropical rivers: The case of the Sorocaba river basin, SP, Brazil. Rev. Biol. Trop. 51:769-782. [ Links ]
Trajano, E. 1997a. Food and reproduction of Trichomycterus itacarambiensis cave catfish from south –eastern Brazil. J. Fish Biol. 51:53-63. [ Links ]
Trajano, E. 1997b. Population ecology of Trichomycterus itacarambiensis, a cave catfish from easter Brazil (Siluriformes, Trichomycteridae). Environ. Biol. Fishes. 50:357-369. [ Links ]
Travnichek, V. & M. Maceina. 1994. Comparison of flow regulation effects on fish assemblages in shallow and deep water habitats in the Tallapoosa river, Alabama. J. Fresh. Ecol. 9:207-216. [ Links ]
Welcomme, R. 1979. Fisheries ecology of floodplain rivers. Longman Londres y Nueva York 317 p. [ Links ]
Williams, D. 1987. The ecology of temporary waters. Croom Helm, Londres. 205 p. [ Links ]
Windell, J. T. 1968. Food analysis and rate of digestion. En Ricker WE, ed. Methods for assessment of fish production in freshwaters: 197-203. Ed. IBP Handbook Nº 3:197-203 p. [ Links ]
Zar, J. 1984. Bioestatistical analysis. Prentice Hall International. USA. 718 p. [ Links ]

