










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.52 suppl.1 San José Sep. 2004
Paralytic Shellfish Poisoning (PSP) in Margarita Island, Venezuela
Amelia La Barbera-Sánchez1 *, Jose Franco Soler2 , Luisa Rojas de Astudillo3 & Ivan Chang-Yen4
1 Centro de Investigaciones Agropecuarias de Sucre/Nva. Esparta, INIA, MCT. Av. Carúpano, sector Caiguire, Cumaná, Estado Sucre, Venezuela. P.O. Box 236, Cumaná 6101. Fax: 58 (93)432-5385; alabarbe@hotmail.com
2 Instituto de Investigaciones Marinas, (CSIC), Vigo, España; jose.franco@vi.ieo.es
3 Department of Chemistry, Universidad de Oriente, Cumaná, Estado Sucre, Venezuela; lrojas40@yahoo.com 4 Department of Chemistry, The University of the West Indies, St. Augustine, WI. Fax: 868-663-9685; icyen@carib-link.net
Recibido 31-X-2002. Corregido 08-IX-2003. Aceptado 11-XII-2003.
Abstract
A severe outbreak of Paralytic Shellfish Poisoning (PSP) occurred in Manzanillo and Guayacán, northwestern coast of Margarita Island, Venezuela, between August and October 1991. A bloom of dinoflagellates including Prorocentrum gracile, Gymnodinium catenatum and Alexandrium tamarense seemed to be responsible for this outbreak. Levels of PSP toxins in mussels (Perna perna) exceeded the international safety limit of saxitoxin, 80 µg STX/100 g meat. PSP toxin values varied between 2 548 and 115 µg STX/100 g meat in Manzanillo, and between 1 422 and 86 µg STX/100 g meat in Guayacán. At both locations, the highest levels were detected in August, when 24 patients exhibited typical symptoms of PSP toxicity after consuming cooked mussels (16 required hospitalization). A high pressure liquid chromatographic (HPLC) procedure was recently used on the 1991 samples. The major toxin detected in samples of both locations was decarbamoyl saxitoxin (dcSTX), but low concentrations of saxitoxin were also found in Manzanillo samples. Gonyautoxins GTX1, GTX2 and GTX3 were detected only at Guayacán, while in both locations, decarbamoylgonyatouxin (dcGTX2,3) toxins were detected. These findings represent the first time that causative toxins of PSP in Venezuela have been chemically identified, and confirm the presence of dcSTX and dcGTX in mussels from the Caribbean Sea. The presence of dcSTX and dcGTX in shellfish is indicative that Gymnodinium catenatum was a causative organism for outbreak of PSP.
Key words: Dinoflagellate, Paralytic Shellfish Poisoning, saxitoxin, Prorocentrum, Gymnodinium catenatum, Alexandrium tamarense.
Palabras clave: Dinoflagelado, intoxicación paralizante por moluscos, saxitoxina, Prorocentrum, Gymnodinium catenatum, Alexandrium tamarense.
Margarita Island is located in northeastern Venezuela in the southern Caribbean. Upwellings occur around the island and its waters are characterized by a high productivity that supports a thriving fishing industry, which supplies seafood to the tourism sector of the island. The fishing villages in Manzanillo and Guayacán are located on the northwestern side of Margarita Island, on which natural banks of the Perna perna mussels thrive. Many villagers depend exclusively on mussel exploitation for a living.
Unfortunately, toxic dinoflagellate blooms occur in the northern marine environment of Margarita. They represent a hazard to public health and cause significant economic losses to the shellfish industry. The first toxic outbreak in Manzanillo was recorded in April 1977 caused by Gonyaulax tamarense var. excavata (Reyes- Vásquez et al. 1979). During this event, twelve patients exhibited symptoms of PSP, including a two-years-old child who died after consuming contaminated mussels. Subsequently, in early August 1991, the inhabitants of Manzanillo and Guayacán reported water discoloration that covered a large area. About the same time, 24 patients showing severe symptoms of PSP were hospitalized in Porlamar, capital city of Nueva Esparta state.
Dinoflagellate toxins are accumulated in shellfish, such as oysters, mussels and scallops, exposed to dinoflagellate blooms, and become toxic to humans and other organisms that consume such shellfish (Oshima et al. 1987). PSP causes muscular paralysis, respiratory difficulties, and can lead to death (Thibault et al. 1991). The neurotoxins belonging to the PSP toxin group are among the most notorious poisons found in the marine environment in many parts of the world (Genenah and Shimizu 1981, Park et al. 1986). There are more than 20 different PSP toxins (Oshima 1995), with others being reported (Quilliam et al. 2001). Collectively, these PSP are termed saxitoxins, deriving the name from the butter clam, Saxidomus giganteus, from which saxitoxins were originally extracted and identified (Bates and Rapoport 1975).
Saxitoxins are neurotoxins that retard the movement of sodium ions through nerve cell membranes, and may block the flow of nerve impulses to cause the symptoms of PSP toxicity, which include paralysis and disorientation (Mosher et al. 1964). There is no antidote for PSP, and all cases require immediate medical attention that may include application of life support equipment to save a victims life. (Kao 1993). Thus, the toxic event of 1991, in Margarita Island, resulted in the activation of an emergency monitoring plan to determine its causes.
The number of different PSP toxins and their tendency for chemical transformation into other toxic products are major factors hindering development of a simple field test kit for measuring PSP toxins (Sullivan and Wekell 1988). Currently, only the mouse bioassay method (Horwitz 1980), adopted as the official method by the Association of Official Analytical Chemists (AOAC) in 1965, is approved by US Food and Drug Administration (USFDA), because it simultaneously measures the total toxicity of all the saxitoxins in a sample of shellfish tissue. However, mouse bioassays have a number of drawbacks, namely the inability to efficiently detect the low toxicity analogues of STX (toxins B and C) (Casais 1991). HPLC analyses of shellfish have thus been developed to confirm the causative toxins of PSP and to determine the toxin profiles of PSP-contaminated organisms (Franco and Fernández 1993, Oshima 1995, Lawrence and Niedzwiadek 2001, Vale and Sampayo 2001).
The most commonly used techniques for the analysis of PSP involve the conversion of non-fluorescent PSP toxins to fluorescent compounds by oxidation reactions. The fluorescent derivatives are then detected and quantified by fluorometry.
The method of HPLC described by Sullivan et al. (1985), which incorporates post- column derivatization to carry out the chemical reaction and originally proposed by Bates and Rapoport (1975), was used to determine PSP in mussels collected from Manzanillo and Guayacán on Margarita Island.
Materials and methods
Mussel and phytoplankton samples were collected at Manzanillo and Guayacán, Margarita Island, during August to December 1991 (Fig. 1). Phytoplankton samples were taken with Niskin bottles and with nets (30 µm), and fixed with lugol solution, and formalin (4%), respectively. The phytoplankton abundance was obtained following the technique described by Utermöhl (1958). All shellfish were externally washed, with a brush and distilled water, to minimize contamination of their tissues and then frozen until analysis. Sampling frequency in the bivalve banks was increased until no PSP was detected by mouse bioassay.
Extracts of the mussels were prepared according to the Official Method of the AOAC (Horwitz 1980). After boiling for 5 minutes and adjust pH between 2-4, the extract was centrifugated at 14 000 rpm for 20 min at 20ºC; 10 µl of the supernatant of each sample were used for HPLC analysis.
Certified calibration solutions (STX; NEOSTX; GTX1; GTX2; GTX3; GTX4) from the National Research Council of Canada were used as diluted to prepare the calibration standards, using Milli-Q water as diluent. The dcSTX was provided from a BCR (European Union) program of PSP intercalibration. Two mixtures of toxin standards were prepared, one mixture contained, NeoSTX, dcSTX and STX, and another one contained GTX1, GTX2, GTX3 and GTX4.
Analysis of the principal toxins was carried out on a HPLC with fluorescence detection using, with minor modifications, the post-column oxidative fluorescence method of Franco and Fernández (1993).
The LC system used was composed of a high-pressure pump (Hitachi L-6000) with an intelligent autosampler (Hitachi AS-4000). The HPLC column used was a 5 µm Lichrospher 100 RP-18 cartridge (12.5 cm x 4 mm i.d), held in a LichroCART 125-4 (Merck) cartridge holder, enclosed in a column heater unit (Jones Chromatography, 7971) at 30°C. Two A-30- SW-2 Eldex pumps with inline degasser (Waters ILD) were used for both oxidizing reagent and acid. The postcolumn reaction was performed in a teflon coil (10 m x 0.5 mm i.d.) held in glass tube (at 65°C) and immersed in a water bath connected to a circulating temperature control unit (Ultratherm 6000383). The detector was a spectrofluorimeter (Jasco FP-920), set at 330 nm excitation and 390 nm emission. Millennium 2010 (v. 215.01) software was used for recording and integrating peaks.
The first isocratic phase (for separation of neoSTX, dcsTX and STX) was 94% of 1 mmol/l sodium octanesulphonate in 10 mmol/l ammonium phosphate (pH 7.2) and 6% acetonitrile, at 1.0 ml/min. The second isocratic phase (for separation of GTX toxins) was 1.5 mmol/l sodium octansulphonate in 10 mmol/l ammoniumphosphate (pH 7.0), at 0.8 ml/min.
The presence of toxins C, GTX5 and GTX6 were confirmed by boiling the sample extract with an equal volume of 0.4 mol/l HCl for 15 minutes. This procedure hydrolyses the sulphocarbamoyl groups. The hydrolysis of C toxins produces the corresponding GTX1 through 4, and GTX5 and GTX6 give rise to STX and neoSTX (Sullivan et al. 1985, Oshima et al. 1990).
The post-column oxidant was 7 mmol/l periodic acid in 50 mmol/l sodium phosphate buffer, adjusted with 1 mol/l ammonium hydroxide to pH 9. Acetic acid (0.5 mol/l) was used as acidifier. The flow rate of both oxidant and acid was 0.4 ml/min.
Post-column oxidative fluorescence method was evaluated by injection of uncontaminated mussel samples with PSP toxin standards and by calculating the recovery values for these experiments.
Toxin concentrations in samples extracts were determined by comparing the peak areas of each toxin with those of the prepared calibration standards.
Toxicities were determined and compared with those obtained by mouse bioassay. To compare the toxicity calculated from the HPLC to that measured by bioassay, the individual toxicity of each toxin were added and converted to µg STX equivalents per 100 g shellfish meat by assuming a conversion factor (CF) (Horwitz 1980) of 0.2 µg STX per mouse unit. All HPLC analyses were run in duplicates and results presented are means of these duplicate determinations.
Results
Microscopic analysis of the water in the vicinity of the mussel banks showed the presence of Gymnodinium catenatum and Alexandrium tamarense; at concentrations of £ 20 cell/ml. A member of the genus Cochlodinium sp. was also detected at very low densities. The most abundant diatoms were Cyclotella sp., Navicula sp., Nitzchia seriata and Rhizosolenia delicatula, while the most abundant dinoflagellates were Ceratium furca, Dinophysis cf. acuminata, Gonyaulax polygramma, Prorocentrum gracilis, and Polykrikos polikrikoides. At both locations, Prorocentrum gracile was the dominant species, and its density indicated that it was the major bloom-forming species. Mesodinium rubrum, a bloom-forming species in the Gulf of Cariaco, was also found at low abundance.
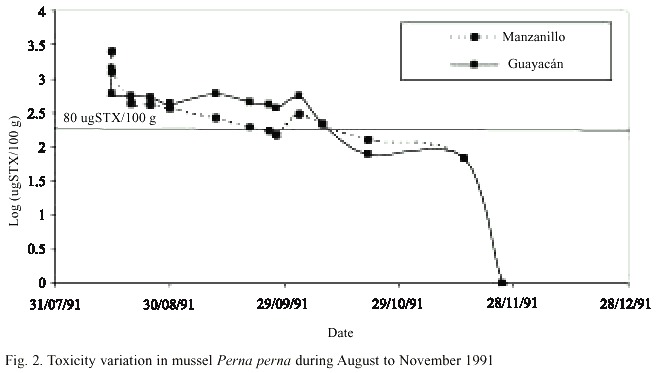
Concentrations of PSP, analyzed by mouse bioassay, in 1991, were higher than the Maximum Permissible Level (MPL) established by USFDA for human consumption, this is 80 µg STX/100 g meat of shellfish. The ranges found were from 115 to 2 548 µg STX/100 g meat in Manzanillo and 86 to 1 422 µg STX/100 g meat in Guayacán. In both areas, toxin concentrations increased during the period August-September; and remained above the MPL until October 15th (Fig. 2). After that date, values decreased and became non-detectable in November and December 1991.
Due to the unavailability of chemical confirmatory technology in Venezuela at the time of the PSP outbreak, samples of shellfish tested by standard mouse bioassay were held at -15ºC until recent acquisition of training in PSP toxins analysis by HPLC. No significant changes in the values of PSP were obtained by analyzing the samples once more by standard mouse bioassay, at the same time that the HPLC analyses were performed.
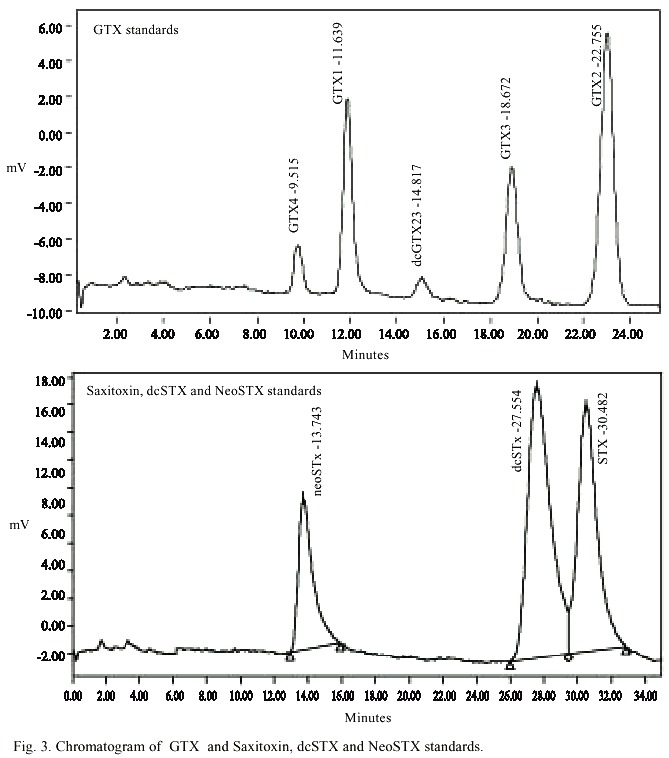
HPLC analyses with post-column derivatization was carried out to identify and determine the toxins responsible for the outbreak of PSP in Manzanillo and Guayacán. This enabled the separation of the epimers GTX1,4 and GTX2,3 (Fig. 3), as well as enabled toxin dc-STX to be distinguished from STX. The mean value of triplicate injection and the fluorescent intensity of each toxin was plotted against its concentrations.
Recoveries from injected samples were between 84-103%. Significant correlation (r = .90; p<.05) between mouse bioassay and HPLC with fluorescence detection was obtained. However, the bioassay results were, in general, lower than those from HPLC, considering no quantified toxins (dcGTX) for lacking of quantitative standards (Table 1). The cause of these differences is unclear, although the selection of toxicity factors (FC) for LC quantification may be implicated (Lawrence et al. 1995). It is also possible that the mouse bioassay underestimates the amount of toxicity present (Ledoux and Hall 2000).
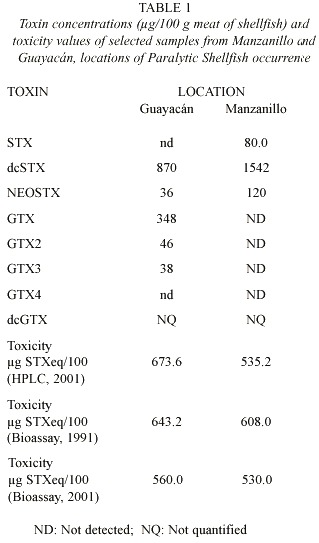
Shellfish samples analyzed using HPLC were lethal to mice. Samples from Manzanillo contained STX, but dcSTX was found the highest concentration (1 542 µg/100 g meat) (Table 1). The PSP toxin profiles of P. perna samples collected from Manzanillo (Margarita Island) showed that the major toxins present were dcGTX and dcSTX groups (Fig. 4).
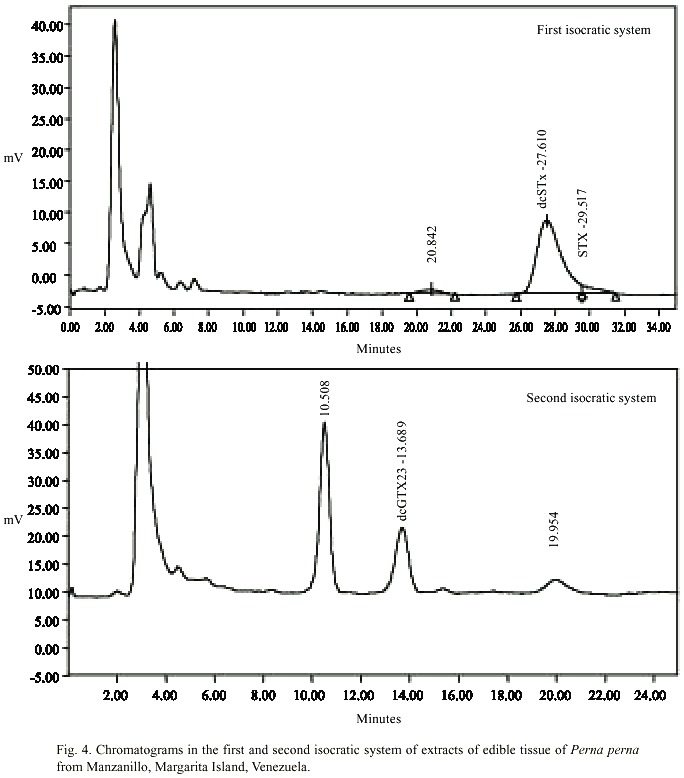
The toxins dcGTXs were identified using a qualitative standard obtained from a collaborative study, but their presence could not be quantified because of the lack of calibrants. The availability of the calibrants for dcGTX is essential for wide use of the method, and attention should be given to projects aimed at the development of calibrants for decarbamoyl toxins (Van den Top et al. 2000).
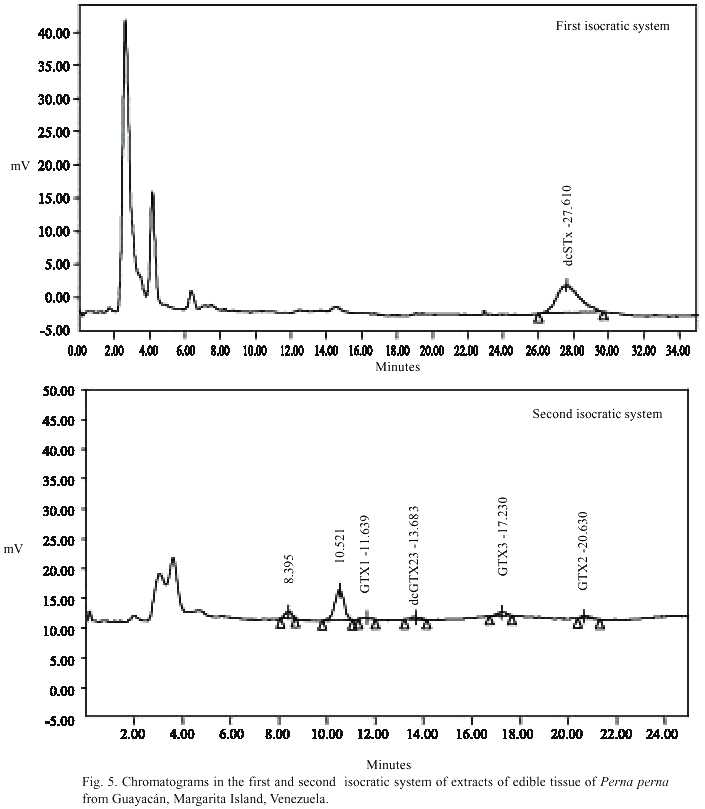
From Guayacán, dcSTX and dcGTX were also detected in samples of P. perna, but at lower concentration when compared with Manzanillo samples (Fig. 5). Samples of mussels from Guayacán also contained GTX1, GTX2 and GTX3 at low concentrations (Fig. 5). A large unidentified peak with retention time 10.5 min., near GTX6 and GTX4, was observed in samples from both locations (Figs. 5 and 4), similar to that reported by Oshima et al. 1995.
Discussion
These findings agree with previous reports of toxins other than STX being dominant in contaminated shellfish (Noguchi et al. 1983, Lassus et al. 1989, Carreto et al. 1996, Amzil et al. 1999, Jiang et al. 2000).
Our analyses have confirmed the presence of dcSTX in our mussel samples, following the outbreak of PSP toxicity in Manzanillo and Guayacán (Margarita Island). Harada et al. 1983 also found this toxin among tropical specimens.
G. catenatum and A. tamarense have been reported in Venezuela as toxigenic and have been associated with other toxic outbreaks (La Barbera-Sánchez et al. 1993). A characteristic feature of G. catenatum toxins is their production of decarbamoyl toxins (Oshima et al.1990, Donker et al. 1997, Negri et al. 2000). Mussels and oysters showed similar toxin profiles to those of the causative dinoflagellate, with a slightly higher amount of decarbamoyl and carbamoyl toxins (Oshima et al. 1990). Thus G. catenatum appears to be one of the organisms responsible for the incidence of PSP toxins initially investigated by standard mouse bioassay. The compilation of toxin profiles, as chemical signatures of G. catenatum populations, is the subject of continued investigation (Hallegraeff and Fraga 1998).
Analysis of A. tamarense by Hummert et al. 1997 showed only N-sulfocarbamoyl toxins in this species; it is likely that this species was also another contaminant dinoflagellate.
With the results of this investigation, toxic profiles of shellfish from Venezuela coast are available and can be compared in detail with those obtained from other countries.
Acknowledgments
We acknowledge the collaboration of Rommel Delgado from the Ministry of Health in Porlamar for providing the data on number of patients affected by the toxic outbreak in that locality during 1991. We also thank the cooperation of Simón Silva, Oswaldo Gallardo and Luis Gómez from the Toxicology Laboratory, Instituto Nacional de Investigaciones Agricolas (INIA-Sucre). We acknowledge the support of Technological Development Program, phase II, subscribed between Venezuelan Government and the Interamerican Development Bank, Instituto de Oceanografia, Spain, Intergovernmental Oceanographic Commision (IOC) of UNESCO, Consejo de Investigación of Universidad de Oriente, Venezuela, and The University of the West Indies, Trinidad.
Resumen
Un severo brote de intoxicación paralizante por moluscos (PSP en inglés) ocurrió en Manzanillo y Guayacán en la costa noroeste de la Isla de Margarita, Venezuela entre agosto y octubre de 1991. Una proliferación de Prorocentrum gracile, Gymnodinium catenatum y Alexandrium tamarense causó el brote. Los niveles de PSP en mejillón (Perna perna) superaron los niveles máximos permisibles de saxitoxina, 80 µg STX/100g carne. Los niveles de toxinas variaron entre 2 548 y 115 µg STX/100 g carne en Manzanillo y entre 1 422 y 86 µg STX/100g carne en Guayacán. En ambas localidades, los máximos niveles se detectaron en agosto, cuando 24 personas presentaron síntomas típicos de PSP después de consumir mejillones cocidos (16 fueron hospitalizados). Se aplicó recientemente cromatografía líquida de alta presión (HPLC) a muestras del año 1991 y la toxina más detectada fue decarbamoyl saxitoxina (dcSTX), pero también se encontró saxitoxinas en muestras de Manzanillo. Las gonyautoxinas GTX1, GTX2 y GTX3 solo se encontraron en Guayacán; en ambas localidades se detectó decarbamoylgonyatouxin (dcGTX2,3). Estos hallazgos representan la primera vez que las toxinas causantes de un brote de PSP en Venezuela han sido químicamente identificadas, confirmando la presencia de dcSTX y dcGTX en mejillones del mar Caribe. La presencia de dcSTX y dcGTX en moluscos, indica que G. catenatum fue el organismo responsable de la intoxicación.
References
Amzil, Z., M. Quilliam, T. Hu & J.L.C. Wright. 1999. Winter accumulation of paralytic shellfish toxins in digestive glands of mussels from Arcachon and Toulon (France) without detectable toxic plankton species revealed by interference in the mouse bioas say for lipophilic toxins. Nat. Toxins 7: 271-277. [ Links ]
Bates, H.A. & H. Rapoport. 1975. A chemical assay for saxitoxin, the paralytic shellfish poison. J. Agric. Food Chem. 23: 237-239. [ Links ]
Carreto, J., C. Elbusto, H. Sancho, M. Carignan, T. Yasumoto & Y. Oshima. 1996. Comparative studies on paralytic shellfish toxin profiles of marine snails, mussels and an Alexandrium tamarense isolate from the Mar del Plata coast (Argentina). Rev. Invest. Des. Pesq. 10: 101-107. [ Links ]
Casais, C. 1991. Gymnodinium catenatum toxins from mussels (Mytilus galloprovincialis). Environ. Technol. 12: 33-40. [ Links ]
Donker, S., M.I. Reyero, B. Reguera & J.M. Franco. 1997. Perfil de toxinas PSP de seis cepas de Gymnodinium catenatum de Galicia, pp. 69-76. In J.M Wyeites & F. Leira (eds.). V Reunión Ibérica sobre Fitoplancton Tóxico y Biotoxinas, Vigo, España. [ Links ]
Franco, J.M. & P. Fernández-Villa. 1993. Separation of paralytic shellfish toxins by reversed phase high performance liquid chromatography with postcolumn reaction and fluorimetric detection. Chromatographia 35: 613-620. [ Links ]
Genenah, A.A. & Y. Shimizu. 1981. Specific toxicity of paralytic shellfish poisons. J. Agric. Food Chem. 29: 1281-1291. [ Links ]
Hallegraeff, G.M. & S. Fraga. 1998. Bloom dynamics of the toxic dinoflagellate Gymnodinium catenatum, with emphasis on Tasmanian and Spanish coastal waters, pp. 125-152. In D.M. Anderson, A.D. Cembella & G.M. Hallegraeff (eds.). Physiological ecology of harmful algal blooms, Berlin. [ Links ]
Harada, T., Y. Oshima & T. Yasumoto. 1983. Natural occurence of decarbamoylsaxitoxin in Tropical dinoflagellate and bivalves. Agric. Biol. Chem. 47: 191-193. [ Links ]
Horwitz, W. 1980. Official methods of Analysis of the Association of Official Analytical Chemists. Washington D.C. pp. 298-299. [ Links ]
Hummert, C., M. Ritscher, K. Reinhardt & B. Luckas. 1997. Analysis of the characteristic PSP profiles of Pyrodinium bahamense and several strain of Alexandrium sp. by HPLC based on ion-pair chromatographic separation, post-column oxidation and fluorescence detection. Chromatographia 45: 312-316. [ Links ]
Jiang, T., Y. Yin, Z. Chen, Y. Luo, W. Huang & Y. Qi. 2000. HPLC analysis of paralytic shellfish toxins in shellfish from Daya Bay. Mar. Environ. Sci. 19: 16-19. [ Links ]
Kao, D.Y. 1993. Paralytic shellfish poisoning, pp. 75-86. In I.R. Falconer (ed.). Algal toxins in seafood and drinking water. San Diego, CA. [ Links ]
Koehn, F.E., V.E Ghazarossian, E.J. Schantz, H.K Schnoes & F.M. Strong. 1981. Derivatives of saxitoxin. Bioorg. Chem. 10: 412-428. [ Links ]
La Barbera-Sánchez, A., S. Hall & E. Ferraz-Reyes. 1993. Alexandrium sp., Gymnodinium catenatum and PSP in Venezuela, pp. 281-285. In T.J. Smayda & Y. Shimizu (eds.). Toxic Phytoplankton Blooms in the Sea, Amsterdam. [ Links ]
Lassus, P., J.M. Fremy, M. Ledoux, M. Bardouil & M. Bohec. 1989. Patterns of experimental contamination by Protogonyaulax tamarensis in some french commercial shellfish. Toxicon 27: 1313-1321. [ Links ]
Lawrence, J.F. & B. Niedzwiadek. 2001. Quantitative determination of paralytic shellfish poisoning toxins in shellfish by using prechromatographic oxidation and liquid chromatography with fluorescence detection. J. AOAC Int. 84: 1099-1108. [ Links ]
Lawrence, J.F., C. Ménard & Ch. Cleroux. 1995. Evaluation of prechromatographic oxidation for liquid chromatography determination of Paralytic Shellfish Poisons in shellfish. J. AOAC Int. 78: 514-520. [ Links ]
LeDoux, M. & S. Hall. 2000. Proficiency testing of eight french laboratories in using the AOAC mouse bioassay for paralytic shellfish poisoning: Interlaboratory collaborative study. J. AOAC Int. 83: 305-310. [ Links ]
Mosher, H.S., E.A. Fuhrman, H.S. Buchwald & H.G. Fischer. 1964. Tarichatoxin-Tetrodoxin: A potent neurotoxin. Science 144: 1100. [ Links ]
Noguchi, T., J. Maruyama, Y. Onoue, K. Hashimoto & T. Ikeda. 1983. Toxins of mussels infested with Protogonyaulax catenella isolated from Senzaki bay. Yamaguchi Prefecture. Bull. Japan. Soc. Sci. Fish. 49: 499. [ Links ]
Oshima, Y. 1995. Post column derivatization liquid chromatographic method for paralytic shellfish toxins. J. AOAC Int. 78: 528-532. [ Links ]
Oshima, Y., K. Sugino, H. Itakura, M. Hirota & T. Yasumoto. 1990. Comparative studies on paralytic shellfish toxin profile of dinoflagellates and bivalves, pp. 391-396. In E. Graneli, B. Sundstrom, L. Edler & D.M. Anderson (eds.). Toxic marine Phytoplankton, New York. [ Links ]
Oshima, Y., M. Hasegawa, T. Yasumoto, G. Hallegraeff & S. Blackburn. 1987. Dinoflagellate Gymnodinium catenatum as the source of paralytic shellfish toxins in Tasmanian shellfish. Toxicon 25: 1105-1111. [ Links ]
Park, D., W.N. Adams, S. Graham & R. Jackson. 1986. Variability of mouse bioassay for determination of paralytic shellfish poisoning toxins. J. AOAC Int. 69: 547-549. [ Links ]
Quilliam, M.A., C. Dell' Aversaro & P. Hess. 2001. Analysis of PSP toxins by liquid chromatographymass spectrometry. 15th AOAC International Annual Meeting. September 9th -13th , Kansas City, Missouri. [ Links ]
Reyes-Vásquez, E., E. Ferraz-Reyes & E. Vásquez. 1979. Toxic dinoflagellate blooms in northeastern Venezuela during 1977, pp. 191. In D.L. Taylor, H.H. Seliger (eds.). Toxic Dinoflagellate Blooms, New York. [ Links ]
Sullivan, J., & M. Wekell. 1988. Detection of paralytic shellfish toxins, pp. 87-106. In T. Anthony (ed.). Handbook of Natural Toxins, New York. [ Links ]
Sullivan, J., M.M. Wekell & L. Kentala. 1985. Application of HPLC for the Determination of PSP toxins in shellfish. J. Food. Sci. 50: 26-29. [ Links ]
Thibault, P., S. Pleasance & M.V. Laycock. 1991. Analysis of paralytic shellfish poisons by capillary electrophoresis. J. Chromatogr. 542: 483-501. [ Links ]
Utermöhl, H. 1958. Zur vervollkommung der quantitativen phytoplankton-methodic. Mitt. Int. Verein. Theor. Angrew. Limnol. 9: 1-38. [ Links ]
Vale, P. & M.A.M. Sampayo. 2001. Determination of paralytic shellfish toxins in Portuguese shellfish by automated pre-column oxidation. Toxicon 39: 561-571. [ Links ]
Van den Top, H.J., A. Boenke, P.A. Burdaspal, J. Bustos, H.P. Van Egmond, T. Legards, A. Mesego, A. Mouriño, W.E. Paulsch & C. Salgado. 2000. The development of reference materials for paralytic shellfish poisoning toxins in lyophilized mussel. I: Interlaboratory studies of methods of analysis. Food Add. Contam. 17: 419-433. [ Links ]
Internet References
Negri A.P., C.J.S. Bolch, S.I. Blackburn, M. Dickman, L.E. Lewellyn & S. Mendez. 2000. Paralityc shellfish toxins in Gymnodinium catenatum strains from six countries. Procceedings of the 9th International Conference on Harmful Algal Blooms (Tasmania). (Downloaded: September 8th , 2003, http://www.utas.edu.au/docs/plant_science/HAB200 0/poster_abstracts/docs/Negri_Andrew_P.html) [ Links ]

