










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.52 n.4 San José Dec. 2004
Composition of the zooplankton community,with emphasis in copepods, in Punta Morales,Golfo de Nicoya,Costa Rica
Ernesto Brugnoli-Olivera1, Edgardo Díaz-Ferguson2, Mariana Delfino-Machin3, Alvaro Morales- Ramírez3 &Arturo Dominici Arosemena4
1 Sección Oceanología,Facultad de Ciencias,Universidad de la República.Igua 4225. Montevideo, Uruguay; ebo@fcien.edu.uy
2 Laboratorio de Genética y Microbiología,Facultad de Ciencias del Mar,Universidad de Cádiz. Cádiz, España.
3 Escuela de Biología,Universidad de Costa Rica,Apdo.Postal 2060,San José,Costa Rica.
4 Center for Tropical Marine Ecology (ZMT),Fahrenheitstrasse 6-28359.Bremen,Germany.
Received 26-II-2003. Corrected 01-III-2004. Accepted 02-IV-2004.
Abstract:The composition of the mesozooplanktonic community was studied in the Punta Morales estuary, Gulf of Nicoya,Pacific coast of Costa Rica,during 1997.Oblique plankton hauls were performed during high and low tide using a 280 µm mesh screen net equipped with a flowmeter.The community was characterized by holoplanktonic and meroplanktonic organisms.For the holoplanktonic community,the main groups were copepods (80%)and chaetognaths (16%).The most abundant species were the copepods Acartia lilljeborgii and Paracalanus parvus.A.lilljeborgii is a typical estuarine species that maintains high populations in estuarine systems.Meroplankton was represented mainly by crustacean larvae (66%),and icthyoplankton (18%).The dominance of crustacean larvae and icthyoplankton is an evidence of the ecological importance of the Punta Morales zone.Rev.Biol.Trop.52(4):897-902.Epub 2005 Jun 24.
Key words:Abundance,holoplankton,meroplankton,copepods,tropical estuary.
Organisms which are part of zooplankton are either classified as holoplankton or meroplankton on the basis of the time they remain on the planktonic communities;the first are part of the zooplanktonic communities during all their life cycle,have relatively low diversity and are dominated by copepods,in abundance and biomass (Longhurst 1985).Copepods present a high morphological variety (Dudley 1986), include approximately 8500 species, occur in nearly marine,brackish,freshwater or inland salines water (Raymont 1983).On the other side,meroplankton is abundant in neritic environments and is formed by eggs or larval stages of organisms,which presence in the pelagic environment is only temporal (Raymont 1983).Larval stage of estuarine invertebrate species develops a mechanism to maintain their population in their parental stock;this mechanism is called larval retention and is observed in template and tropical estuarine populations (Epifanio and Dittel 1982).
The Gulf of Nicoya is a tropical estuary, located at 10º N and 85º W,in the Pacific coast of Costa Rica (Klemas et al.1981).From an oceanographic point of view,it is possible to divide in an inner and an outer zone according to hydrographic,physical and chemical condition (Voohris et al .1983,Lizano and Alfaro 2004).Zooplankton studies in this system have been concentrated on identification and description separately on the holo and meroplanktonic component.In recent studies holoplankton chaetognaths of the species Sagitta friderici were found showing a seasonal variation of abundance (Hossfel 1996)and Morales and Vargas (1995)give a list of 12 calanoid species for the outer part with species of copepods Acartia lilljeborgii and Paracalanus parvus, being very common.On the other hand,Dittel and Epifanio (1990)described the meroplanktonic organism and behavior of retention larval of crustacean larvae.The goals of this study were determinate the composition of mesozooplankton (holo and meroplankton)community, quantifies the abundance of zooplankton groups and copepods species in the zone of Punta Morales,Gulf of Nicoya,Costa Rica.
The sampling station was located in the area known as Punta Morales (10º 5 N and 84º 57 W),in the inner zone of the Gulf of Nicoya. This area was visited in two consecutive days in high and low tide,during March 1st, 2nd, April 30th, May 1st, 21st, 22nd, September 27th, 28th, October 18th, 19th, November 15th, 16th and December 6th, 7th of 1997.Oblique tows for three minutes at a speed of one knot were realized using a net with a mesh size of 280 µm. The total volume of water filtered was determined using a General Oceanics Inc.flux meter model 2030 R.Samples were fixated in 5% of formaline and carried to the laboratories for counting and identification.Samples were fractionated in a Folson plankton splitter;all the subsamples were placed on a Bogorov chamber, observed on a Wilde M 5 A stereoscope,sorted into major taxa and quantified. The copepods were sorted,identify to species and quantify.Newell and Newell (1975),Smith (1977)and Boltovskoy (1981),keys were used to identify the zooplankton taxa,while Todd &Laverack (1991),Campos y Suarez (1994) and Morales y Vargas (1995)keys were used to identify copepod species.In order to quantify the abundance we selected 18 samples (three in high and low tide during transition and rainy seasons).According to Brugnoli (1999) December and March were determined as dry season,April,May as transition season and September,October and November as rainy season. The abundance of all taxa in these samples were calculated and expressed as relative abundance or annual mean (ind m-3).
During the sampling period the mesozooplankton community was characterized by the presence of different groups,such as copepods, chaetognaths,crustacean larvae,ostracods, cladocerans, icthyoplankton (fish larvae), molluscs, appendicularians, echinoderms and brachiopods larvae (Table 1).
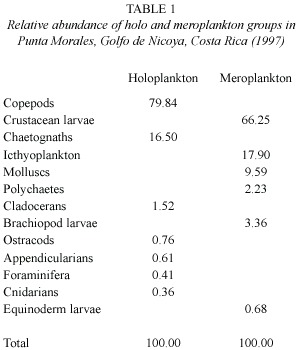
The mesozooplanktonic community during the study show a mean of 4404 ind m-3 (range:1458 ind m-3 -9142 ind m-3).In that community,copepods were the most abundant group with a mean of 2602 ind m-3 (438 ind m-3 -5877 ind m-3),followed by crustacean larva with a mean of 758 ind m-3 (47 ind m-3 a 3249 ind m-3)(Table 2).
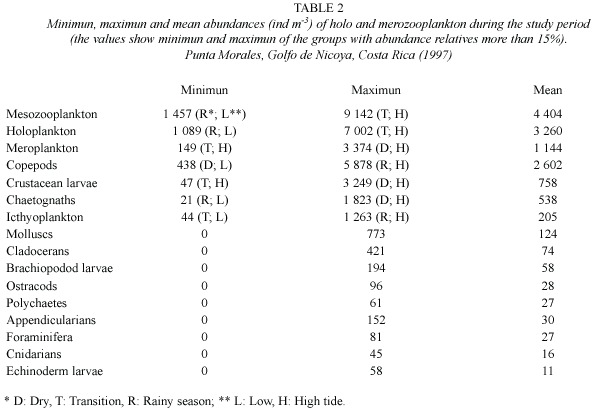
The holoplanktonic organisms shows a mean of 3260 ind m-3 (range:1089 ind m-3 - 7002 ind m-3)and the meroplankton, 1144 ind m-3 (149 ind m-3 -3374 ind m-3 ).Holoplankton shows copepods as the most abundant group followed by chaetognaths;meroplankton was represented principally by crustacean larvae and icthyoplankton (Table 2).
In the copepods,a total of 12 species were identified,with A.lilljeborgii being the dominant species with a mean of 2154 ind m-3 (range:307 ind m-3 of 5242 ind m-3)followed by Pseudiaptomus sp.and Paracalanus parvus (Table 3).
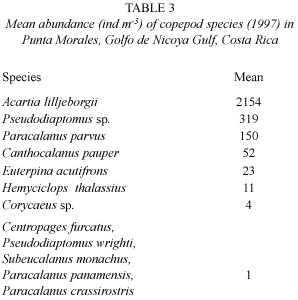
The zooplankton community composition (Table 1)corresponds to Raymont ´s (1983) description of estuarine zooplankton communities and these results fit with the estuarine status suggested for the Gulf of Nicoya by Peterson (1958)and Voohris et al .(1983).In the holoplankton,copepods were the more abundant group (Table 2),according to Raymont (1983). The number (12)of observed copepod species (Table 3)was in accordance to Morales and Vargas (1995) and agrees to the low diversity suggested for estuarine zones (Hopcroft et al . 1998).However,Hopcroft et al .(1998) pointed the presence of small sized species,in tropical zones,which are not collected effectively by nets with mesh sizes over 64 µm (Chisholm and Roff 1990), suggesting that the present research may has underestimated the number of copepod species due to the mesh size of the net used.Morales-Ramírez (2001)suggests the same problem of mesh size in the determination of copepod diversity.
The more abundant species of copepods during the sampling period were A.lilljeborgii, Pseudiaptomus sp.and P.parvus (Table 3).According to Björnberg (1981),such species are typically estuarine which confirms the characteristics of the Gulf of Nicoya.At the same time,the presence of Centropages furcatus and Canthocalanus pauper (Table 3) has being reported by Suárez-Morales y Gasca (1989)for the Costa Rican dome and suggested an oceanic influence in the Punta Morales region during the sampling period.In our study we found more mean abundance if compared with other copepods (Pontellidae)studies in Tropical Oriental Ocean Pacific (Álvarez-Silva et al .2003)however,the copepod abundance can vary betwen 100-70 000 ind m-3 in different oceans (Suárez-Morales 1995).
The species A.lilljeborgi (Table 3)were the most abundant.These results are in accordance with Raymont (1983),who states that holozooplankton tends to be more abundant and less diverse in estuarine areas,due to the dominance of certain species.The dominance of the genus Acartia agrees with others studies in coastal zone (Palomares-García and Gómez-Gutierrez 1996).Acartia is a typically estuarine genus,generally dominating in such system (Kimmerer and McKinnon 1987a)and its presence is probably due to its availability to maintain high clearance rates (Paffenhöfer and Stearns 1988),to its high reproductive rates and it s omnivorous condition (Palomares-García and Gómez-Gutierrez 1996).
Zooplankton maintains their populations in the estuary zone through a vertical migration behaviour in response to tidal direction (Kimmerer and Mc Kinnon 1987b).This behaviour was more relevant in larvae (Cronin 1982). However,Wooldridge and Erasmus (1980),suggest that this form of migration may also be widespread among Acartia species in tidal bays. This kind of behaviour has been observed in vertical profiles of zooplankton which includes bottom and surface samples when we consider a complete tidal cycle (Wooldridge and Erasmus 1980,Kimmerer and Mc Kinnon 1987b).
On the other hand,we have found differences in some groups of the zooplankton in terms of the retention mechanism. Some groups have shown more abundance during the high tide than others and some may not (Table 2).This fact can be explained due to the sort of sampling method (oblique tows without complete the tidal cycle).These results are not enough to confirm the existence of the cited behaviour in the copepod species that we have identified.In spite of this,some authors have found differences in abundance of crustaceans between tides in the Punta Morales zone (Dittel et al.1991,Díaz-Ferguson y Vargas 2001).
The dominance of crustacean larvae in the meroplankton (Table 1)is an evidence of its ecological importance for the zone,in benthic (Dittel et al .1985)or planktonic (Epifanio and Dittel 1984)communities and its relationship with zones located close to the sampling site. There is a tendence of crustaceans to use estuarine zones for breading and spawning (Dittel and Epifanio 1990).On the other hand,the high percent of icthyoplankton (Table 1)with the suggest for Ramirez et al.(1989),who mention the Punta Morales zone as a breeding and hutching area for different fish species,confirm the importance of this zone.
Acknowledgments
We thank the suggestion for the editor and two anonymous referee to improve the article.The financial support was provided by Vicerrectoría de Investigación,Universidad de Costa Rica, project N º 808-97-272,and the supervision of Alvaro Morales.The study is part of the first author s MSc.Thesis and received and AECI- Mutis fellowship.We also thank Marco Quesada for the translating the manuscript.
Resumen
Se estudió la composición de la comunidad mezoplanctónica en Punta Morales,Golfo de Nicoya, Pacífico de Costa Rica,en 1997,arrastrando oblicuamente una red de plancton (280 µm,con flujómetro)durante marea alta y baja.La comunidad estuvo caracterizada por la presencia de organismos holo y meroplanctónicos.Los principales grupos holoplanctónicos fueron copépodos (80%)y quetognatos (16%).Las especies más abundantes fueron los copépodos Acartia lilljeborgii y Paracalanus parvus;A. lilljeborgii es una especie estuarina típica que mantiene altas poblaciones en sistemas estuarinos.El meroplancton estuvo representado principalmente por larvas de crustáceos (66%),e ictioplancton (18%).La dominancia de larvas de crustáceos e ictioplancton evidencian la importancia ecológica de la zona de Punta Morales.
Palabras clave:Abundancia,holoplancton,meroplancton, copépodos,estuario tropical.
References
Álvarez-Silva,C.,M.G.Guadalupe Miranda Arce &G. De Lara-Isassi.2003.Familia Pontellidae (Crustacea: Copepoda)en la Bahía La Ventosa,Oaxaca,México: Sistemática y ecología básica.Rev.Biol.Trop.51: 737-742. [ Links ]
Björnberg,T.K.1981.Copepoda,p.578-679.In D. Boltovskoy (ed.).Atlas de Zooplancton del Atlántico Sudoccidental y métodos de trabajo con el zooplancton marino.INIDEP,Mar del Plata,Argentina. [ Links ]
Boltovskoy,D.1981.Atlas del zooplancton del Atlántico Sudoccidental y metodología de trabajo con el zooplancton marino. INIDEP,Mar del Plata,Argentina.933 p. [ Links ]
Brugnoli,E.1999.Factores abióticos y bióticos que regulan la estructura y dinámica de la comunidad fitoplánctica en la zona de Punta Morales,Golfo de Nicoya.Tesis de maestría,Universidad de Costa Rica,San José,Costa Rica. [ Links ]
Campos,A.&E.Suarez.1994.Copépodos pelágicos del Golfo de México y Mar Caribe.I.Biología y sistemática. Centro de Investigaciones de Quintana Roo, México.353 p. [ Links ]
Cronin,T.1982.Estuarine retention of larva of the crab Rhithropanopeus harrisii .Estuar.Coast.Shelf.Sci. 15:207-220. [ Links ]
Chisholm,L.&J.C.Roff.1990.Abundances,growth rates,and production of tropical neritic copepods of Kingston, Jamaica. Mar.Biol.106:79-89. [ Links ]
Díaz-Ferguson,E &J.A.Vargas.2001.Abundance of Petrolisthes armatus (Crustacea:Porcellanidae)in a tropical estuarine intertidal rocky beach,Gulf of Nicoya estuary,Costa Rica.Rev.Biol.Trop.49:97-101. [ Links ]
Dittel,A.,C.E.Epifanio &J.B.Chavarría.1985.Population biology of the Portunid crab Callinectes arcuatus Orway in the Gulf of Nicoya,Costa Rica,Central America.Estuar.Coast.Shelf Sci.20:593-602. [ Links ]
Dittel,A.&C.E.Epifanio.1990.Seasonal and tidal abundance of crab larvae in a tropical mangrove system, Gulf of Nicoya, Costa Rica.Mar.Ecol.Progr.Ser. 65:25-34. [ Links ]
Dittel,A.,C.E.Epifanio &O.Lizano.1991.Flux of Crab Larvae in Mangrove Creek in the Gulf of Nicoya, Costa Rica. Estuar. Coast. Shelf Sci.32:129-140. [ Links ]
Dudley,P.C.1986.Aspects of general body shape and development in Copepoda.Syllogues 59:7-25. [ Links ]
Epifanio,C.E.&A.Dittel.1982.Comparison of dispersal of crab larvae in Delaware Bay (USA),and the Gulf of Nicoya,Central America,p.447-487.In V. Kennedy (ed.)Estuarine Comparisons.Academic. New York. [ Links ]
Hopcroft,R.,J.C.Roff &D.Lombard.1998.Production of tropical copepods in Kingston Harbour,Jamaica: the importance of small species.Mar.Biol.130: 593-604. [ Links ]
Hossfel,B.1996.Distribution and biomass of arrow worms (Chaetognata)in Golfo de Nicoya and Golfo Dulce, Costa Rica.Rev.Biol.Trop.44:157-172. [ Links ]
Kimmerer,W.&A.D.McKinnon.1987a.Zooplankton in a marine bay.I.Horizontal distribution used to estimate net population growth rate.Mar.Ecol.Prog. Ser.41:43-52. [ Links ]
Kimmerer,W.&A.D.McKinnon.1987b.Zooplankton in marine bay.II.Vertical migration to maintain horizontal distributions. Mar. Ecol.Progr.Ser.41:53-60. [ Links ]
Klemas,V.,S.Ackleson,M.Murillo &J.A.Vargas.1981. Water quality assement of Golfo de Nicoya,Costa Rica.Progress report of the 1980-1981.International Sea Grant Program,University of Delaware, College of Marine Studies,Newark.1096 p. [ Links ]
Longhurst,A.1985.The structure and evolution of plankton communities.Progr.Oceanogr.15:1-35. [ Links ]
Lizano O.G.&E.J.Alfaro.2004.Algunas características de las corrientes marinas en el Golfo de Nicoya, Costa Rica.Rev.Biol.Trop.52 (Suppl.2):77-94. [ Links ]
Morales,A.&J.A.Vargas.1995.Especies comunes de copépodos (Crustacea:Copepoda)pelágicos del Golfo de Nicoya, Costa Rica.Rev.Biol.Trop.43: 207-218. [ Links ]
Morales,A.1996.Checklist of copepods from Gulf of Nicoya,Coronado Bay and Golfo Dulce,Pacific coast of Costa Rica,with comments on their distribution.Rev.Biol.Trop.44:103-113. [ Links ]
Morales-Ramírez,A.2001.Bi odi versi dad marina en Costa Rica,los microcrustáceos:Subclase Copepoda (Crustacea:Maxillopoda).Rev.Biol. Trop.49:115-133. [ Links ]
Newell,G.&R.Newell.1975.Marine Plankton.A practical guide.Hutchinson,London.221 p. [ Links ]
Paffenhöfer,G.&D.Stearns.1988.Why is Acartia tonsa (Copepoda:Calanoida)restricted to nearshore environments? Mar. Ecol. Progr.Ser.42:33-38. [ Links ]
Palomares-García,R.&J.Gómez-Gutierrez.1996. Copepod community structure at Bahia Magdalena, Mexico during El Niño 1983-1984.Estuar.Coast. Shelf Sci.43:583-595. [ Links ]
Peterson,C.L.1958.The physical oceanography of the Gulf of Nicoya,Costa Rica,a tropical estuary.Bull. Inter. Ame. Trop. Tuna Comm.3:139-216. [ Links ]
Ramírez,A.,W.A.Szelistowski &M.López.1989. Spawning pattern and larval recruitment in Gulf of Nicoya anchovies (Pisces:Engrulidae).Rev.Biol. Trop.37:55-62. [ Links ]
Raymont,J.1983.Plankton and productivity in the oceans. Vol.2.Zooplankton.Pergamont.Oxford.824 p. [ Links ]
Suárez-Morales,E.&R.Gasca.1989.Copépodos calanoides epidemiológicos del Domo de Costa Rica (Junio- Agosto,1982).Cien.Mar.15:89-102. [ Links ]
Suárez-Morales,E.1995.Copépodos.Con qué se comen? Inf.Cient.Tecn.México.17:18-21. [ Links ]
Smith,D.1977.A guide to marine coastal plankton and marine invertebrate larvae.Kendall &Hunt. EUA.140 p. [ Links ]
Todd,C.&M.Laverack.1991.Coastal marine zooplankton.A practical manual for students.Cambridge. 106 p. [ Links ]
Voohris ,A.,D.Epifanio,C.E.Maurer,A.Dittel & J .Vargas .1983.The estuarine character of the Gul f of Nicoya,an embayment on t he Pacific Coast of Central America.Hydrobiology 99: 225-237. [ Links ]
Wooldridge,T.&T.Erasmus.1980.Utilization of tidal currents by estuarine zooplankton. Estuar. Coast. Mar. Sci.2:107-114. [ Links ]

