










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.52 n.4 San José Dec. 2004
Sexual reproduction of the reef-building coral Diploria labyrinthiformis (Scleractinia:Faviidae),in the Colombian Caribbean
Elvira M.Alvarado Ch.1 ,Rocío García U.2 &Alberto Acosta3
1 Centro de Investigaciones Científicas (Museo del Mar),Universidad de Bogotá Jorge Tadeo Lozano,Santafé de Bogotá,D.C.,Colombia.Fax:57-1-2812840;elvira.alvarado@utadeo.edu.co
2 Department of Marine Sciences,University of Puerto Rico -Mayaguez Campus,P.O Box 908,Lajas P.R 00667; rochy98@hotmail.com
3 Pontificia Universidad Javeriana,K 7 No 43-82,Ed Carlos Ortiz.Oficina 502,Bogotá,Colombia; laacosta55@hotmail.com
Received 26-IV-2002. Corrected 06-I-2003. Accepted 20-II-2003.
Abstract:Sexual reproduction of the coral Diploria labyrinthiformis was studied for the first time.Monthly histological analyses at the Corales del Rosario National Park (Colombian Caribbean) from May 1997 to April 1998 show that D.labyrinthiformis is a hermaphroditic broadcasting species.It presents an annual gametogenic cycle with a 10-11 month period for gonad investment,in which oogenesis begins in August and ends in May- June.Spermiogenesis is short because sperm cysts were only observed in May tissue samples.In histological collected in May,an average of four mature eggs and six spermatic cysts per fertile mesentery were found.The mean diameter of mature eggs was 297 µm (±97 SD)and 90 µm (±33)for spermatic cysts.Rapid maturation of eggs from stage II to stage III coincides with increases in air temperature,high number of solar hours per month,decreases in wind velocity and absence of rainfall.Reproductive effort for D.labyrinthiformis (14.07 mm3/cm2/year)was similar to other Faviidae species.Although gamete release was not observed in the field,the absence of gonads in histological samples in June suggests spawning between May 25 (five days after full moon) and June 24.This event coincides with high air temperature,low number of solar hours per month,low wind velocity,and initiation of the rainy season.The earlier spawning time of this species differs from other species of the same family known for the Caribbean region.Rev.Biol.Trop.52(4):859-868.Epub 2005 Jun 24.
Key words:Diploria labyrinthiformis ,coral reproduction,gametogenesis,Caribbean coral reefs, Colombia.
Studies on sexual reproduction of hermatypic corals have increased during the last two decades,with information of about 30%of the known species currently available.From 91 species of Faviidae within the Indo-Pacific region,information of only 40 species is known (Kojis and Quinn 1982,Harriot 1983, Harrison and Wallace 1990)and from 17 species of the Caribbean region,no more than eight species have been studied (Szmant 1986, Soong 1991,Acosta and Zea 1997).
Faviid species are mainly hermaphroditic and present gonads developing as local thickenings of the mesentery,in contrast with those of Pocilloporidae,which are connected by a short stalk to the mesenteries and projected into the body cavity (Rinkevich and Loya 1979)or those of Siderastrea siderea, which are attached to the mesenterial filaments (Alvarado and Guzman unpublished). In the Colombian Caribbean,information on the reproductive biology of corals is available for the following species:Porites porites (Hernández and Manrique 1991),Acropora palmata (García 1994),Acropora cervicornis (Sanjuan 1995),Tubastrea coccinea (Uscategui 1996)and Montastraea cavernosa (Acosta 1992,Acosta and Zea 1997)as well as on spawning season of Montastraea annularis and M.faveolata (Sánchez et al.1999).
Diploria labyrinthiformis is a common species of the coral community in some areas of the Caribbean region and yet its reproductive biology is,so far,unknown.Previous studies on reproductive characteristics of Diploria strigosa (Wyers 1985,Szmant 1986,Wyers et al.1991,Soong 1991)and Diploria clivosa (Soong 1991)document traits within the genus,such as reproductive patterns,gonadal arrangement,number of eggs per ovary,and size and shape of eggs.Gametogenic cycle of D.strigosa and spawning observations of D.strigosa and D.clivosa indicates a 9-10 month cycle with spawning occurring during the warmest season (July-September)synchronized with other faviid species.The only information we found in the literature is the release of traces of eggs from two colonies of D. labyrinthiformis in laboratory conditions, during late July (Wyers et al.1991).
The present paper describes,for the first time,the reproductive pattern,mode of development, gametogenic cycle,gonadal arrangement,egg size,number of eggs per gonad, and reproductive effort of D.labyrinthiformis . These sexual characteristics were related to abiotic factors as well as compared with other species within the genus.
Materials and methods
The study was carried out at the Isla Grande coral reef (Caribbean coast of Colombia), between May 1997 and April 1998.This island forms part of the Parque Nacional Natural Corales del Rosario,which is located between 10 º08 -10 º26 N and 75 º30 -75 º48 W (Fig. 1).The area lies within the Intertropical Convergence Zone,under trade winds that alternate from northwest to southeast.Three climatic seasons have been described for the area,a dry season,occurring between December and April and dominated by the Alisios winds;an intermediate season (mild rainfall)between May and July;and a rainy season (heavy rainfall),which occurs between August and November (Leble and Cuignon 1987).The annual average for seawater temperature is 27.5 ºC;precipitation is 1 013.1 mm and salinity 35 0/00 (Sánchez 1999).
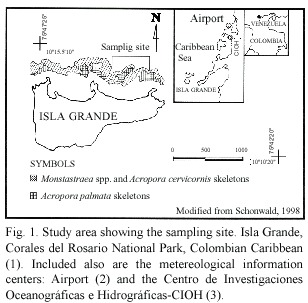
Due to legal restrictions within the Protected Area,only three colonies per month were sampled.Each month,5 days after full moon,two samples of 2.5 cm in diameter, per colony,were extracted from the center of three different colonies,located at depths of 7-12 m.These were fixed in Zenker solution for 24 hours,washed in running tap water for 24 hours and stored in 70%ethanol (Acosta and Zea 1997),until histological procedures. Tissue samples were dehydrated in alcohol and xylene series,embedded in Paraplast and serially sectioned at 6 µm.Sections,taken every 200-250 µm,were stained with Heidenmhain s Azocarmine Aniline Blue (Luna 1968,Acosta and Zea 1997).Seven to nine slides with 4-8 sections each,and 20-56 mesenteries per section,were observed for each colony under light microscope.The slide containing the most reproductive material was used to estimate a) number of fertile mesenteries,b)total number of eggs and sperm cysts (Guzmán and Holts 1993),and c)size and stage of these gametes. Since there is not a clear polyp differentiation in this species,all information is referred to area, where 1 cm2 contains 12 to 15 mesenteries.
A number of 220 complete eggs were observed in order to define different development stages based on descriptions for other species (Wyers 1985,Delvoye 1988,Acosta 1992,Glynn et al. 1994, 1996).Three stages for oogenesis were defined using the following characteristics:1)presence of single mesogleal layer surrounding the egg;2)presence and thickness of basal membrane surrounding the egg;3)presence of cytoplasmatic material inside the egg (scarce or abundant cytoplasmatic material);4)presence of vesicular cytoplasmatic inclusions (few or high -giving the egg a bubbly appearance);5)maximum length of eggs using an ocular micrometer (Olympus, OSM-4,10 X).
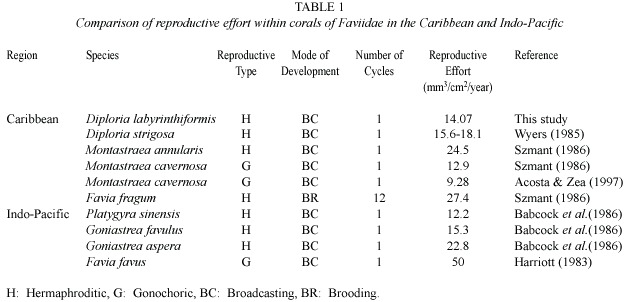
Reproductive effort was calculated with a modification of the Gonadal Index used by Szmant (1986):number of mesenteries per cm2 x number of gonads per mesentery x maximum number of eggs per gonad x volume of eggs per mm 3 x number of cycles per year.The volume was calculated using the formula:V=4/3 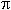 r3.
r3.
Sunlight,air temperature,rainfall and wind velocity data for the study period (May 1997-April 1998)was provided by the Centro de Investigaciones Oceanográficas e Hidrográficas (CIOH)for two near-by sites (airport and Escuela Naval)(Fig.1).
Results
Diploria labyrinthiformis is a hermaphroditic broadcasting species with eggs and spermatic cysts within each mesentery (Fig. 2A).Eggs and spermatic cysts are located towards the aboral part of the mesentery (Fig. 2B).This species displays an annual gametogenic cycle in which oogenesis begins in August and ends between May and June (Fig. 3).Spermiogenesis is short,since sperm cysts were observed only in tissue samples from May.Nevertheless,our observations suggest synchronous maturation between eggs and sperm cysts during May.
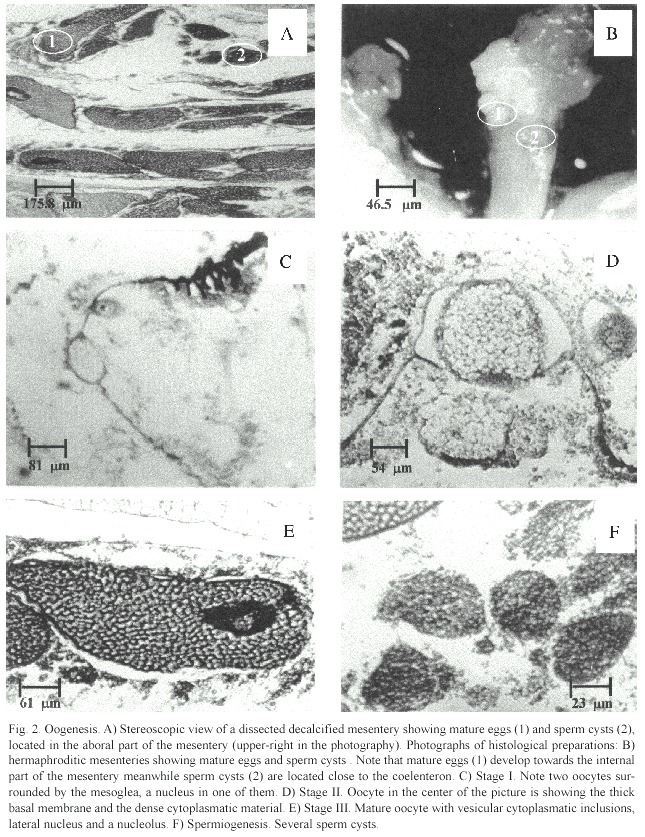
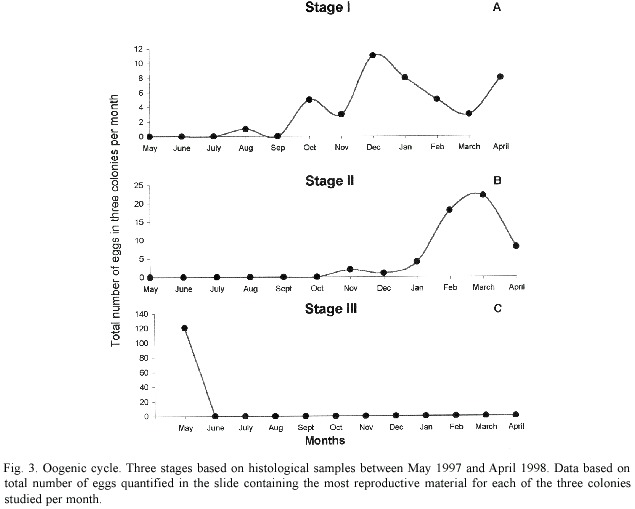
Oogenesis:Stage I oocytes were small, ovoid or rounded and the main characteristic was the presence of a thin,bluish-transparent mesogleal layer surrounding the egg (Fig.2C). Cytoplasmatic material was absent (12%), scarce (86%),or abundant (2%).Mean diameter of oocytes was 116.1 µm (±36.5 S.D.),and range from 53.9 to 213.0 µm (n =44 oocytes). This stage was observed from August to April, with a peak in December (Fig.3A).
Stage II oocytes were also small,ovoid or rounded (Fig.2D),but with a basal membrane that thickened from November (11 µm) to April (33 µm).The majority had scarce cytoplasmatic material (67%),and the rest had either few vesicular cytoplasmatic inclusions or were empty.Mean oocyte diameter was 150.4 µm (±32.6),and range from 57.7 to 209.6 µm (n =55 oocytes).This stage was observed from November to April,with a peak in March (Fig.3B).
Stage III oocytes were enlarged,ovoid or irregular,with a well-defined basal membrane and with high vesicular cytoplasmatic inclusions giving them a bubbly appearance (Fig. 2E).Mean diameter was 297.2 µm (±96.9), and range from 73.5 to 584.3 µm (n =121 oocytes).Mature eggs were only found in May (Fig.3C).
In samples of June and July no eggs were found.This suggests that D.labyrinthiformis may have spawned between May 25 and June 24.Thus,this species has an annual reproductive cycle with a 10-11 months period for gonad investment.
Spermiogenesis:In May,spermatic cysts were only present on 85%of the mesenteries.In no case they were observed without eggs in the same mesentery,which confirms a hermaphroditic pattern.An average of six cysts per fertile mesentery were found,with a mean diameter of 90.1 µm (±32.9),and range from 27.5 to 220.5 µm,(n =160 cysts).Cysts showed either dense (45%)or scarce (55%) cellular material (Fig.2F).
Reproductive effort:On the total mesenteries observed (n =705)for the one-year study period,16%were fertile (n =114),with 379 eggs and 208 spermatic cysts (complete and incomplete).The highest percentage of mature mesenteries was found in May (42%), bearing an average of four eggs per mesentery (range:1-9;21 per cm2)and six spermatic cysts per mesentery (range:1-10;21 cysts per cm2 ).The reproductive effort estimated by the gonad index obtained from mature eggs was 14.07 mm3 /cm2 /year (Table 1).
Discussion
Reproductive type (hermaphroditic), mode of development (broadcaster),gonadal arrangement (oocytes and spermatic cysts in the same mesentery),oocyte mean size and shape,number of eggs per mesentery as well as duration of oogenesis seem to be conservative characteristics within any genus (Soong 1991) (Table 2).The similarity in the reproductive effort between the Faviidae species (Table 1) and particularly for D .labyrinthiformis (this study)and D.strigosa (Wyers 1985)further supports Soong s hypothesis.
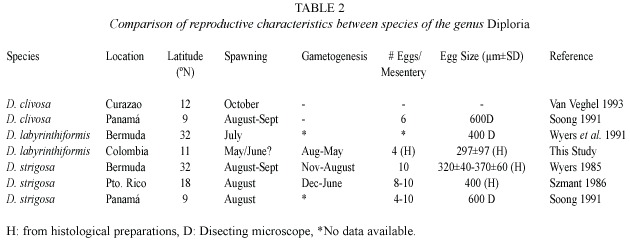
Although the length of the oogenic cycle (10 months)is similar between D.labyrinthiformis and D.strigosa ,the initiation of oogenesis and spawning month differs between the two species.These events occur earlier for D . labyrinthiformis populations in Colombia than for its congeneric species in Bermuda (Wyers 1985)and Puerto Rico (Szmant 1986)(Table 2).Gametogenesis and spawning differences for the same species have been attributed to latitude (Szmant 1986,Soong 1991,Van Veghel 1994, Acosta and Zea 1997,Knowlton et al.1997), temperature,day length or both (Babcock 1995, Fan and Dai 1995,Fadlallah 1996,Ben-David- Zaslow et al.1999),solar energy (Van Veghel 1994),current and winds regimes (Babcock et al.1986)and rainfall (Acosta and Zea 1997).
The first visible egg in D.labyrinthiformis was found during August;while most stage I oocytes were found during October and November,which in Colombia coincides with the highest temperature (Fig.4A).Rapid maturation of eggs (Stage II to Stage III)coincides with highest number of solar hours per month (Fig 4B)and rising temperatures,which happens during the dry season.Spawning occurs during June when wind velocity is lowest (Fig. 4C),and temperature is high.Thus,initiation and maturation of oocytes coincides with environmental conditions reported for species of the same family,by other authors for the Caribbean region (Szmant 1986,Soong 1991, Wyers et al.1991,Van Veghel 1994,Acosta and Zea 1997,Sanchez et al.1999).Nevertheless, gamete release does not occur during the months when most spawning events,known at low latitudes,take place (August-October) (Guzmán and Holts 1993,Van Veghel 1994, Acosta and Zea 1997,Sanchez et al.1999).
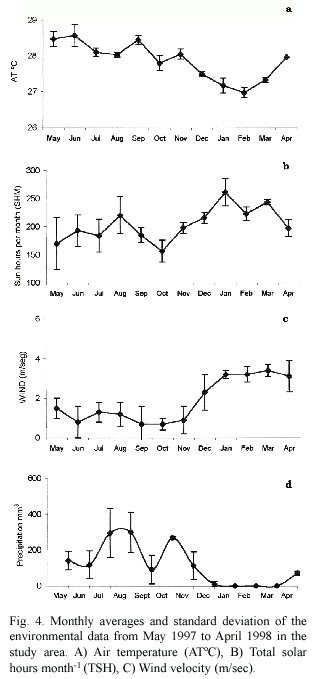
It is worthwhile noting that temperature range for the Caribbean region is 3.2 ºC (Szmant 1986)and for Colombia,during the period studied,the temperature did not vary more than 1 ºC (27.8 ºC ± 0.4;28.4 ºC ± 0.21). As has been noted by other authors,rising temperatures might cue the initiation of gametogenesis in broadcasters,but gamete release is controlled by other factors such as photoperiod (Soong 1991,Szmant 1991),low tides (Kojis and Quinn 1981),low current regimes (Babcock et al.1986),low winds and scarce rainfall (Watson et al.2000).In accordance,Bassim et al.(2002)states that the effects of high seawater temperature do not affect fertilization rates of D.strigosa ;while embryonic development and larval viability decrease dramatically where abnormally high sea surface temperatures occur during the spawning season.
The adaptive value of spawning in late May and early June for D.labyrinthiformis , may be related to appropriate settling space (Fan and Dai 1995),associated with reduction in algae cover caused by local conditions (Soong 1991,pers.obs.in the studied area) or low current regimes and low winds which may ensure that egg clumps will not be carried out by currents and thus,favor high rates of fertilization (Kojis and Quinn 1981),which are the environmental conditions at the study site during these months.Timing of reproduction is of taxonomic significance and may reflect species recognition problems within a group (Johnson 1992).Therefore,the difference in timing of gametogenic cycle in D.labyrinthiformis may represent a mechanism of reproductive isolation to prevent hybridization and maintain species boundaries within the genus (Miller and Babcock 1997),or a strategy for self fertilization,as has been reported for D.strigosa (Hagman et al.1998).Finally,but not least,it is possible that the effects of El Niño event,which was present during 1997, could have affected the cycle of gametogenesis and hence, spawning. But even then,it remains unclear why this event should have affected this species and not others in the same site as M.annularis and M.faveolata that spawned during September and October (Sánchez et al. 1999),as has been reported through out the Tropical Caribbean region.
Acknowledgments
This work is part of the project "Reproduction,growth and transplantation survivorship of the coral species Montastraea annularis ,Diploria labyrinthiformis and Porites astreoides in the National Natural Park Corales del Rosario,Colombian Caribbean", (Universidad Jorge Tadeo Lozano and COLCIENCIAS 1202-09-221-96). Research Permission No.568,15 de octubre de 1996, Ministerio del Medio Ambiente, Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales.We thank J.A.Sánchez,O.Arenas,H.Charry,M.Gil and L.Chasqui,for their assistance during the whole course of this study.Also to Sport Barú, Centro de Investigaciones Oceanográficas e Hidrográficas (CIOH),ECOBUZOS and all those who helped in field or laboratory, as well as those who gave us logistic facilities in different phases of the research.We thank M.Zea and P.Yoshioka who proofread the English version and A.Acero,J.Cortés,H.Guzmán and anonymous reviewers who helped improve the manuscript. Contribution to Marine Science No.10 of Centro de Investigaciones Científicas, Universidad de Bogotá Jorge Tadeo Lozano.
Resumen
La reproducción sexual del coral Diploria labyrinthiformis fue estudiada por primera vez.Análisis histológicos mensuales de muestras de D.labyrinthiformis extraídas en el Parque Nacional Natural Corales del Rosario (Caribe colombiano)entre mayo de 1997 a abril de 1998,muestran que es una especie hermafrodita,liberadora de gametos. Presenta un ciclo gametogénico anual de 10-11 meses para inversión en gónadas,en el cual la oogénesis comienza en agosto y termina en mayo-junio.La espermatogénesis es corta,ya que los paquetes espermáticos solo fueron observados en muestras de tejido del mes de mayo.En las muestras histológicas de mayo,un promedio de cuatro huevos maduros y seis paquetes espermáticos por mesenterio fértil fueron encontrados.El diámetro promedio de los huevos fue de 297 µm (±97 DS)y de 90 µm (±33) para los paquetes espermáticos.La rápida maduración de los huevos del estadío II al III coincidió con aumentos en la temperatura del aire,mayor número de horas de sol por mes,disminución en la velocidad del viento y ausencia de lluvias.El esfuerzo reproductivo de esta especie (14.07 mm3 /cm2/año)fue similar al de otras especies de Faviidae. Aunque la liberación de gametos no fue observada en el campo,la ausencia de gónadas en las muestras histológicas del mes de junio sugieren que la liberación ocurrió entre el 25 de mayo (cinco días después de luna llena)y el 24 de junio.Este evento coincidió con altas temperaturas del aire,bajas velocidades del viento y la iniciación de la época lluviosa.El tiempo de liberación de gametos en esta especie difiere de otras de la misma familia,conocidas para la región del Caribe.
Palabras clave:Diploria labyrinthiformis ,reproducción, gametogénesis,arrecifes coralinos, Caribe, Colombia.
References
Acosta, A.1992.Reproducción sexual del coral Montastraea cavernosa (Scleractinia:Faviidae)en la región de Santa Marta,Caribe Colombiano.M.Sc. Thesis.Universidad Nacional de Colombia.Santa Marta,Colombia.Pp.102. [ Links ]
Acosta,A.&S.Zea.1997.Sexual reproduction of the reef coral Montastraea cavernosa (Scleractinia: Faviidae)in the Santa Marta area,Caribbean coast of Colombia.Mar.Biol.128:141-148. [ Links ]
Babcock,R.C.,G.Bull,P.L.Harrison,A.J.Heyward, J.K.Oliver,C.C.Wallace &B.L.Willis.1986. Synchronous spawning of 105 scleractinian coral species on the Great Barrier Reef.Mar.Biol.90: 379-194. [ Links ]
Babcock,R.1995.Synchronous multispecific spawning on coral reefs:potential for hybridization and roles of gamete recognition.Reprod.Fert.Develop.7: 943-950. [ Links ]
Bassim,K.M.,P.W.Sammarco &T.L.Sneer.2002.Effects of temperature on success of (self and non-self) fertilization and embryogenesis in Diploria strigosa (Cnidaria,Scleractinia).Mar.Biol.140:479-488. [ Links ]
Ben-David-Zaslow,R.G.Henning,D.K.Hofmann & Y.Benayahu.1999.Reproduction in the Red Sea soft coral Heteroxenia fuscescens :seasonality and long-term record (1991 to 1997).Mar.Biol.133: 553-559. [ Links ]
CIOH.1997.Boletín Metereomarino del Caribe Colombiano.Central de Pronósticos Metereológicos y Oceanográficos.Centro de Investigaciones Oceanograficas e Hidrográficas.Armada Nacional de Colombia.No.23. [ Links ]
CIOH.1998.Boletín Metereomarino del Caribe Colombiano.Central de Pronósticos Metereológicos y Oceanográficos.Centro de Investigaciones Oceanograficas e Hidrográficas.Armada Nacional de Colombia.No 35. [ Links ]
Delvoye,L.1988.Gametogenesis and gametogenic cycles in Agaricia agaricites (L.)and Agaricia humilis Verril and notes on gametogenesis in Madracis mirabilis (Duchassaing & Michelotti) (Scleractinia). Foundation for Scientific Research in Surinam and the Netherlands Antilles,Amsterdam 123:101-134. [ Links ]
Fadlallah,Y.H.1996.Synchronous spawning of Acropora clathrata coral colonies from the western Arabian Gulf (Saudi Arabia).Bull.Mar.Sci.59:209-216. [ Links ]
Fan,T.Y.&D.F.Dai.1995.Reproductive ecology of the scleractinian coral Echinopora lamellosa in the northern and southern Taiwan.Mar.Ecol.Prog.Ser. 123:565-572. [ Links ]
García,R.1994.Crecimiento,sobrevivencia,regeneración y reproducción de la especie Acropora palmata (Lamarck,1886)en el Parque Nacional Natural Corales del Rosario.Tesis de Pregrado para optar el título de Biólogo Marino.Universidad de Bogotá Jorge Tadeo Lozano.Bogotá,Colombia.124p [ Links ]
Glynn,P.W.,S.B.Colley,C.M.Eakin,D.B.Smith,J. Cortés,N.J.Gassman,H.M.Guzmán,J.B.Rosario & J.S.Feingold.1994.Reef coral reproduction in the eastern Pacific:Costa Rica,Panama,and Galapagos Islands (Ecuador).II.Poritidae.Mar.Biol.118: 191-208. [ Links ]
Glynn,P.W.,S.B.Colley,N.J.Gassman,K.Black,J. Cortés &J.L.Maté.1996.Reef coral reproduction in the eastern Pacific:Costa Rica,Panama, and Galapagos Islands (Ecuador).III.Agariciidae (Pavona gigantea and Gardineroseris planulata ). Mar.Biol.125:579-601 [ Links ]
Guzmán,H.&I.Holst.1993.Effects of chronic oil-sediment pollution on the reproduction of the Caribbean reef coral Siderastrea siderea .Mar.Pol.Bull.26: 276-282. [ Links ]
Hagman,D.K.,S.R.Gittings &P.D.Vize.1998. Fertilization in broadcast-spawning corals of the Flower Garden Banks National Marine Sanctuary. Gulf Mex.Sci.16:180-187. [ Links ]
Harriot,V.J.1983.Reproductive ecology of four scleractinian species at Lizard Island,Great Barrier Reef. Coral Reefs 2:9-18. [ Links ]
Harrison,P.L.&C.C.Wallace.1990.Reproduction, dispersal and recruitment of scleractinian corals, pp.133-207.In Dubinsky Z.(ed.).Coral Reefs, Ecosystems of the World.25.Elsevier,Amsterdam. [ Links ]
Hernández,S.&M.C.Manrique.1991.Sobrevivencia a la fragmentación,crecimiento y reproducción sexual de la especie coralina Porites porites en la Isla Pavitos. Parque Nacional Natural Corales del Rosario.Tesis de Pregrado para optar al título de Biólogo Marino. Universidad de Bogotá Jorge Tadeo Lozano.Bogotá, Colombia.129 p. [ Links ]
Johnson,K.G.1992.Synchronous planulation of Manicina areolata (Scleractinia)with lunar periodicity.Mar. Ecol. Prog. Ser. 87:265-273. [ Links ]
Kojis,B.L.&N.J.Quinn.1981.Aspects of sexual reproduction and larval development in the shallow water hermatypic coral,Goniastrea australiensis (Edwards and Haime,1857).Bull.Mar.Sci.31:558-573. [ Links ]
Kojis,B.L.&N.J.Quinn.1982.Reproductive ecology of two faviid corals (Coelenterata:Scleractinia).Mar. Ecol.Prog.Ser.:251-255. [ Links ]
Knowlton,N.,J.L.Maté,H.M.Guzmán,R.Rowan & J.Jara.1997.Direct evidence for reproductive isolation among the three species of the Montastraea annularis complex in Central America (Panama and Honduras).Mar.Biol.127:705-711. [ Links ]
Leble,S.&R.Cuignon.1987.El archipiélago de las Islas del Rosario.Estudio hidrodinámico y sedimentológico. Cartagena. Bol.Cient.CIOH.7:37-51. [ Links ]
Luna,J.G.1968.Manual for histologic staining methods of the Armed Forces Institute of Pathology.McGraw Hill.New York. [ Links ]
Miller,K.&R.Babcock.1997.Conflicting morphological and reproductive species boundaries in the coral genus Platygyra. Biol.Bull.192:98-110. [ Links ]
Rinkevich,B.&Y.Loya.1979.The reproduction of the Red Sea coral Stylophora pistillata .I.Gonads and planulae. Mar. Ecol. Prog.Ser.1:133-144. [ Links ]
Sánchez,J.A.,E.M.Alvarado,M.F.Gil,H.Charry, O.L.Arenas,L.H.Chasqui &R.García.1999. Synchronous mass spawning of Montastraea annularis (E&S)and M.faveolata (E&S)at Rosario Islands,Caribbean coast of Colombia.Bull.Mar. Sci.65:873-879. [ Links ]
Sanjuan,A.M.1995.Crecimiento,regeneración,sobrevivencia y reproducción del coral Acropora cervicornis (Scleractinia:Acroporidae)en el Parque Nacional Natural Corales del Rosario,Cartagena, Colombia.Tesis de Pregrado para optar al título de Biólogo Marino.Universidad de Bogotá Jorge Tadeo Lozano.Bogotá,Colombia.182 p. [ Links ]
Schonwald,N.1998.Distribución y composición de los hábitats marinos asociados a las estructuras arrecifales del área de Isla Grande,Archipiélago del Rosario,Caribe Colombiano.Tesis de Pregrado para optar al título de Bióloga.Universidad de los Andes, Bogotá,Colombia.57 p. [ Links ]
Soong,K.1991.Sexual reproductive patterns of shallow-water reef corals in Panama.Bull.Mar.Sci.49: 832-846. [ Links ]
Szmant,A.M.1986.Reproductive ecology of Caribbean reef corals.Coral Reefs 5:43-54. [ Links ]
Uscategui,A.M.1996.Biología reproductiva de Tubastrea coccinea (Scleractinia:Dendrophyllidae)en el Parque Nacional Tayrona.Caribe Colombiano.Tesis de Pregrado para optar al título de Bióloga. Pontificia Universidad Javeriana, Bogotá,Colombia.97 p. [ Links ]
Van Veghel,M.L.J.1994.Reproductive characteristics of the polymorphic Caribbean reef building coral Montastraea annularis .I.Gametogenesis and spawning behavior.Mar.Ecol.Prog.Ser.109:209-219. [ Links ]
Watson,G.J.,M.E.Williams &M.G.Bentley.2000. Can synchronous spawning be predicted from environmental parameters?A case study of the lugworm Arenicola marina.Mar.Biol.136:1003-1017. [ Links ]
Wyers,S.C.1985.Sexual reproduction of the coral Diploria strigosa (Scleractinia,Faviidae)in Bermuda:research in progress.Proceedings of the 5th International Coral Reef Congress.Tahiti.4: 301-306. [ Links ]
Wyers,S.C.,H.S.Barnes &S.R.Smith.1991.Spawning of hermatypic corals in Bermuda:a pilot study. Hydrobiology 216/217:109-116. [ Links ]

