










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.52 n.1 San José Mar. 2004
Description of the previously unknown tadpole
of Hyalinobatrachium pulveratum (Anura: Centrolenidae)
Heinz Hoffmann
Investigación y Asistencia Agrícola, S.A., Apartado Postal 153, Barva-3011, Costa Rica; inasag@racsa.co.cr
Received 06-I-2003. Corrected 18-VIII-2003. Accepted 10-X-2003.
Abstract
Egg clutches of the centrolenid Hyalinobatrachium pulveratum were sampled in four lowland locations of Costa Rica. The ontogenesis of the tadpoles of this species is documented by periodical descriptions of the larval stages. Larvae of H. pulveratum change their shape during development because of the non-equal growth of some body parts. Due to these changes the larvae in early stages of development differ considerably to those from later stages. Young larvae have a nearly circular cross-section, whereas later larval stages change to a typical flat shape. Some further morphological characteristics like width of the interorbital distance and the oral disk width change but not proportional to larvae length. Numerous measurements on different larval sizes help to recognize the larvae of H. pulveratum in all stages. The striking bright red hyobranchial sinus is the conspicuous characteristic feature ventrally of the hyobranchial apparatus. The use of allometric values for taxonomic and ontogenetic studies is discussed.
Key Words: Centrolenidae, Hyalinobatrachium, Tadpole, Larvae, Morphology, Anatomy, Allometry, Ontogenesis, First description.
Seven species of the Hyalinobatrachium centrolenid frogs have been reported from Costa Rica (Savage 2002, Kubicki 2004). Because Savage (2002) accounted that the tadpoles of Hyalinobatrachium pulveratum (Peters, 1873) still remain unknown, a wide range of larvae stages of this species is described in the present study. Whereas some ratios like "Body length: body width" or "Body length: total length", do not change considerably during larvae growth, other ratios change strongly due to the unproportional growth of some body parts. When the size of the measured specimen is not considered, the averages of many measurements of its body parts have obviously a large statistical variation. The ratios of the interorbital distance with other body parts are the most conspicuous examples for this phenomenon. Therefore, only specimens with same size or equal larvae stages are comparable. On the other side the tadpole age is not a reliable parameter because specimens of the same hatching group can show considerable differences in development time. These facts are not unknown, reason for some authors to prefer middle tadpole stages for description (Duellman 2002). Nodzenski et al. (1989) already discuss correlations between general visceral morphology and body size. Nevertheless, other authors of important old and new tadpole descriptions use few days old larvae or do not specify the larvae size at all. This restricts the relevance for other not studied development stages (Starrett 1960, Cadle and McDiarmid 1990, Jaramillo et al.1997, Ibáñez et al.1999). When the body size is not considered, the exclusive use of averages of ratios values and its corresponding variation rates, implicates that real existing differences between species can be mask. The objective of the present study is not only the description of the still undescribed larvae of H. pulveratum – it will also give an example for the dynamic of ontogenesis of some external tadpole characters.
Materials and methods
Eggs of H. pulveratum were collected from oviposition clutches in seven Costa Rican locations: four of them are situated in the cantón Pococí, two in Guácimo (both are cantons of the northeastern Caribbean lowland, Limón Province), and one clutch was sampled in the southwestern opposite region of Golfito (Puntarenas Province). The parent frogs deposited these clutches on tree leaves, which hung at 2-4 m above small or medium sized streams. Each clutch contained about 50 eggs (Table 1). The eggs were enclosed in a transparent jelly. The jelly is voluminous and appears relatively dilute in consistence when rain allowed previously an adequate hydration.
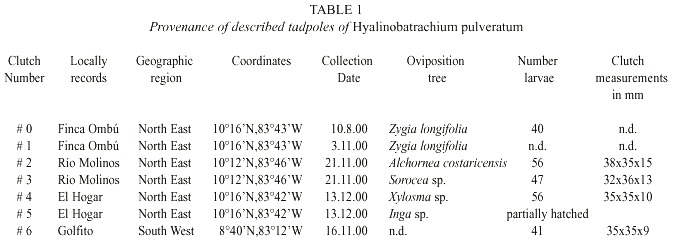
All the clutches were found on the apical third on the leaves upper side. The leaves hung characteristically down by the clutch weight. The embryos were at first bright light-green due to yolk color. All hatchlings included those from Guácimo and Golfito were reared in ambient field conditions in the experimental farm Ombú, Pococí (70 m above sea level). They were held in five to nine liters rainwater buckets with a ground-space of 450 cm2 . The volume of water in the buckets varied, depending on weather conditions such as evaporation and rainfall. River gravel and sand were placed on the bottom of the buckets, and rotting leaf litter and river stones were spread on the top of the sand and gravel layer. The stones were overgrown with algae and small water plants, which gave shelter to a rich microfauna. The stones and leaf litter were changed weekly to provide an abundant food supply for the tad-poles. The larvae developed under the layer of the leaf litter. To prove that the sampled clutches are really such of H. pulveratum,most of the tadpoles of clutches # 1, 2, and 3 were reared to froglets. One frog of these clutches is deposited in the authors collection. The other mentioned tadpole samples did not reach the metamorphic stage.
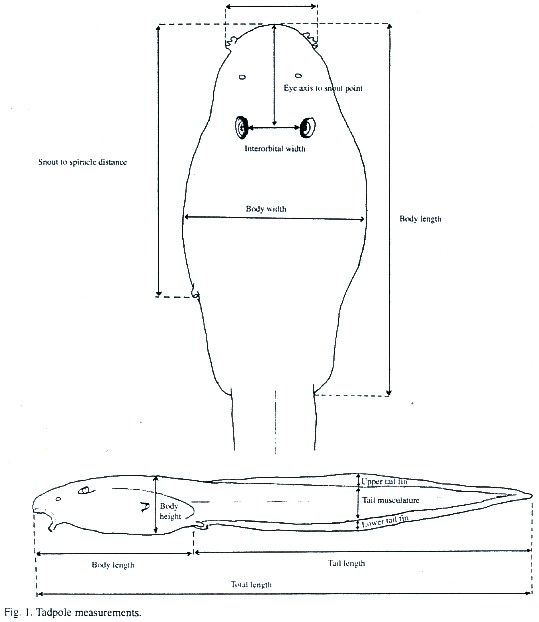
Between August 2000 and April 2001, the tadpoles of the studied populations were sampled for periodical measurements and description. Both were realized on live anesthetic specimens using a dissecting microscope and ocular micrometer at 8 and 25 times augmentation. The tadpoles have been staged according to Gosners (1960) developmental table. After that procedure the tadpoles were immediately preserved in 10% formalin. The measured and preserved specimens are deposited in the authors collection and in the Museum of Zoology of the Universidad de Costa Rica (# 15603). Standard measurements are shown in Fig. 1.
Result
The larvaes Eco Morphological Guild (EMG) may be defined as exotroph, lotic, and burrower, according to the nomenclature of McDiarmid and Altig (1999). Once concluded the embryonic development of between 7 to 12 days the very slender larvae hatch out in stage # 24 to 25 with a total length of 10.0 to 11.5 mm. At this moment they weight about 2.3 mg. In dorsal view the larvae have a dark reddish coloration due to the reddish visceral and muscular system that shine through the hyaline skin. The extreme lateral body parts are completely transparent in dorsal view, because neither muscles nor red colored organs shine through these parts. The reddish color of these Hyalinobatrachium larvae is not conditioned by the skin vascularization as McDiarmid and Altig (1999) described it for this genus. The peripheral vascular blood capillary system that passes through the superficial skin layer gives only space for the flow of a single speedy line of blood corpuscles, and it is therefore unable to influence the general tadpole coloration. The majority of the dorsum is covered with small dark and star-like pigmentation spots. These spots superpose the reddish ground color giving these parts a darker red appearance. The pattern of these spots varies between larvae from different localities, but is fairly symmetrical and reaches in one or more branches down to the ventrolateral body surface (See Fig. 2 for the "Ombú" population). The "Molino" population had more pigmentation than the larvae from Ombú and Golfito. Young Molino larvae had even dorsal, lateral, and ventral tail pigmentation stripes whereas larvae of other populations had only pigmentation spots on the former portion of tail. Although the adults, that are reared from the Molino tadpoles and the adults of the representative Ombú standard population do not show morphological differences, the described larvae differences together with a distinct behavior of the adults of both populations, suggest the possible existence of two different forms. Meanwhile the adult frogs of the other sample places are calmly sleeping during day hours, the Molino frogs have a rather alert diurnal conduct in captivity.
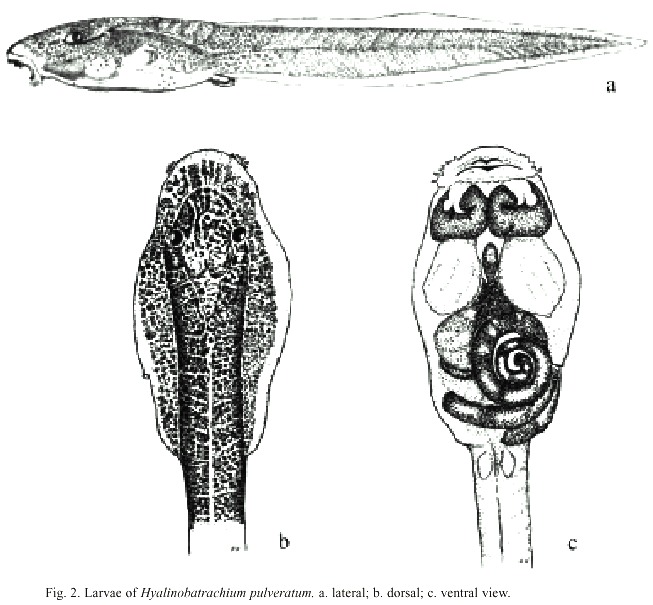
The green yolk determines the color of the caudal part of the larvaes belly during the first days; after about 10 days, the gut spiral is visible and its color varies from grayish to greenish depending on alimentation under the described conditions - now other viscera organs become visible.
In ventral view (Fig. 2) the heart, the sinus venosus and other parts of the central sanguine system are bright red and shine through the hyaline ventral skin. The striking bright red hyobranchial sinus is the conspicuous characteristic feature ventrally of the hyobranchial apparatus. This part of the blood system is small in recently hatched larvae, but later it fills a mayor part of the ventral skull. The reddish liver is accompanied by the dark grayish-green gallbladder. The gills are rose reds.
The bodys width of young larvae (0-4 days) ranges from 1.3 to 1.5 mm and the corresponding measurements of their body height range from 1.0 to 1.3 mm. For that reason, young larvae appear nearly cylindrical in cross sectional view (Ratio "Body width: body height" = 1.06 to 1.30). Although with a wide variation, body grows during developing more quickly wider than vertical (Fig. 3 a). Therefore, body shape of older tadpoles is oval and depressed. The proportion of bodys length to width is equal during larvae growth (Tables 2 and 3, Fig. 3 b), and the percentage of body length in relation to total larvae length increase only in small rates but continuously from about 28 to 33%. Consequently, the percentage of the pointed ochre colored tail decreases from about 72 to 67% (Table 3 and Fig. 3 c). Moreover, larger larvae tend to have a proportionally slender tail than hatchlings. The dorsal fin is low and originates from the tail muscle near tail body junction; dorsal and ventral fins have about the same height, and both fins reach its major height in the central tail part. This part is measured and documented in the tables 2 and 3. Vent tube is medial with slight tendency to dextral position; the sinistral spiracle opens in small larvae on midline at about 3/4 of body length, and at about 2/3 in larger larvae.
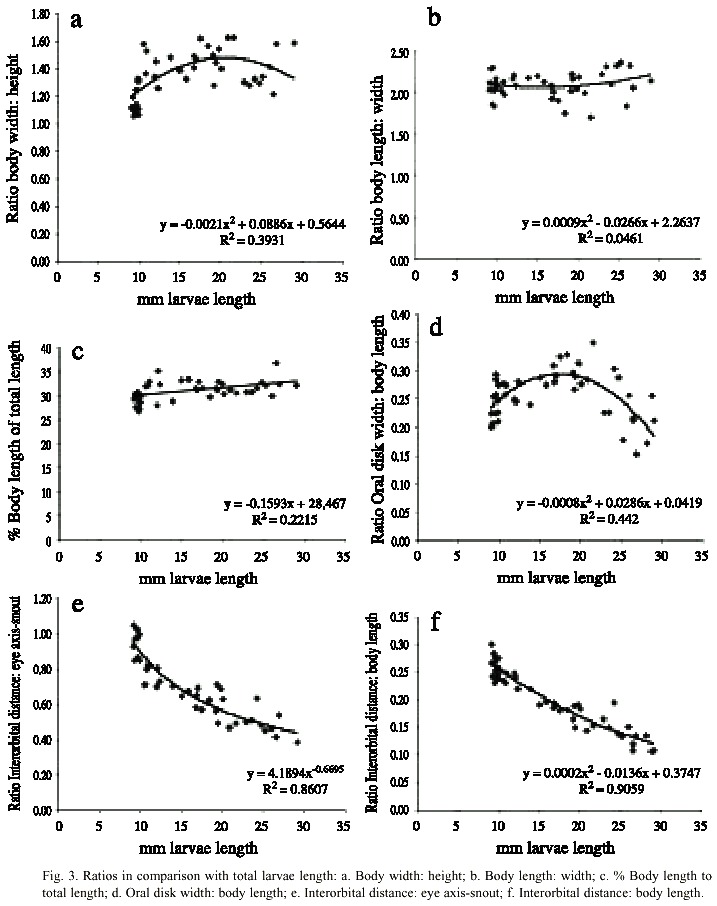
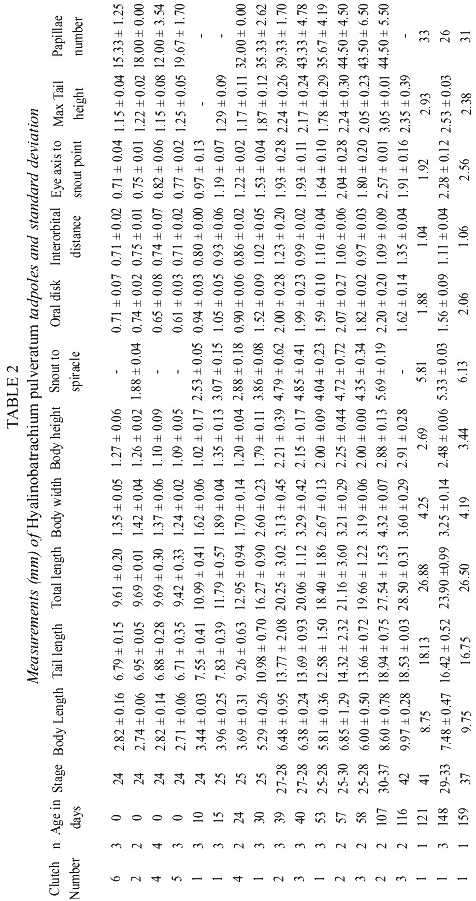
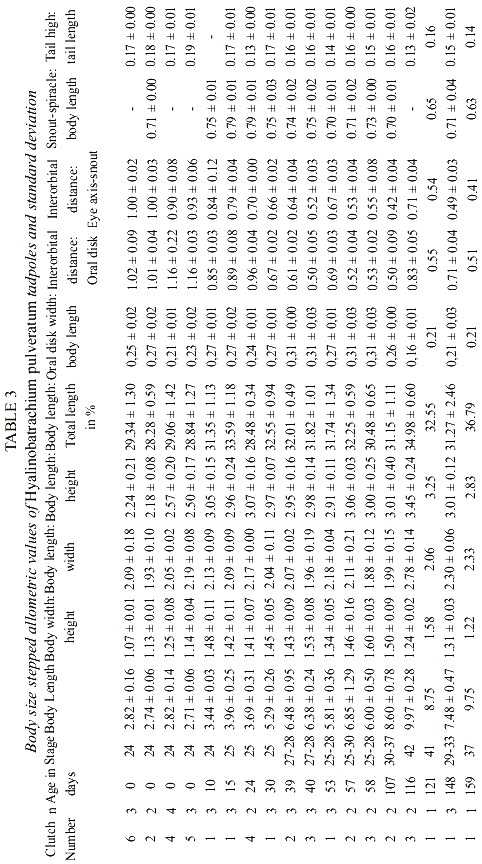
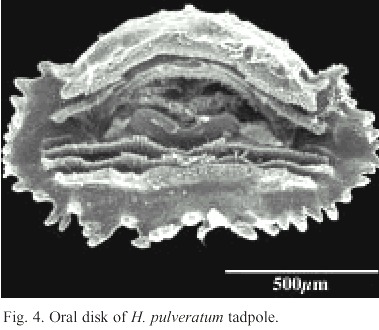
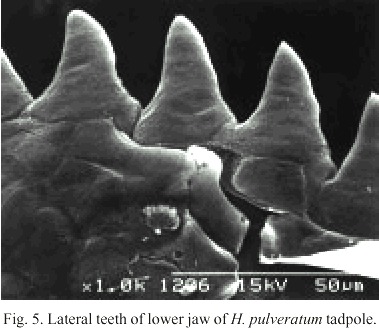
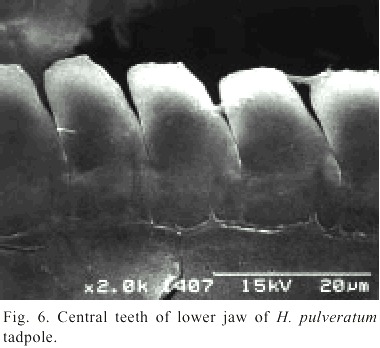
The anteroventral oral disk shows a characteristic ontogenetic change: first, in young larvae, the oral disk grows faster than the body, but later, during the second life period of the larvae, the oral disk does not follow this trend. Therefore, the mouth of hatchlings is relatively small, later stages have a wide mouth, and finally, in still larger larvae it is again relatively small. In Fig. 3 d this phenomenon is visualized with the polynomial curve of the ratio "Oral disk width: body length". This ratio starts from about 0.25 and rises up to 0.30 or more when the larvae reach 15 to 22 mm, but in still larger larvae the oral disk ratio decreases again to values below 0.20. The tooth row formula is 2/3. The A2-tooth-row is broadly separated in the center; P3 is slightly shorter than the equal sized P1 and P2 (Fig. 4). The upper jaw sheath forms a smooth arc and has a pointed serration. It fills half of the total diameter of the oral disk. The lower jaw sheath is smooth and V-shaped. The lateral teeth on the lower jaw are sharp pointed and formed like tiny shark teeth (Fig. 5). The teeth in the center section of the lower jaw sheath are obtuse and melted on the base (Fig. 6). Except for a smaller number of marginal papillae in young larvae, lateral papillae of both sides of the oral disk sum 13 ± 3, the posterior labium has 25 ± 5 papillae, resulting a total number of about 39 ± 7 papillae on the oral disk. The papillae are arranged in a single row but have a wide dorsal gap on the anterior labium (Fig. 4).
The eye position is strongly dorsal, but ratios of interorbital distance to other parameter change markedly with growth of larvae: the eyes of hatchlings are further apart than in older larvae. Consequently, ratio of "Interorbital distance: oral disk" change with growth. Because the snout enlarges, like other body parts, more quickly than the interorbital distance, only the distance between the eyes of hatchlings is for the present nearly as wide as the distance of eye axis to snout tip over the sagittal line. On growing larvae the relative interorbital distance shrinks to the half of the distance of eye axis to snout tip. Consequently, all the other interorbital distance related values show the same tendencies (Fig. 3 e-f). The nares are about equidistant between eye and snout.
Discussion
Some herpetologists might discredit the ratio values used in this study because ratio values have frequently been used incorrectly. Therefore, I emphasized that I am aware that it is statistically incorrect to use allometric methods when the body size or the developing stages of different populations are not comparable. It is misleading for taxonomic studies, when averages of ratio values are taken and summarized from specimens of different size or development stages. The averages of measurements and consequently all its ratio values have too large a statistically variation and lose taxonomic relevance when the stage, size or age of the measured specimen are not considered. That implicates that the alone information of averages of such ratio values and its corresponding standard deviation may mask really existent differences between species. Allometric studies have not only to consider the proportion between two body parts but above all the bodys size or the development stage as the third and most important parameter. Only specimens of the same size or equal stage are comparable. This was considered in this study when allometric values are used for description and visualization of the ontogenesis of the larvae of H. pulveratum. Nevertheless, there are also ratio values of H. pulveratum with few or no changes during growing process, e.g., the ratio of body length to body width and body length to total length. In these special cases, average values and their variation give reliable data for characterizing species. It is recommended to use ratios only for taxonomic description when they are supported by a differentiated body size or stage analysis. For tadpole description, the ontogenetic changes during larvae development require observations and descriptions of more than one single larvae stage. In particular, the description of only a few days old tadpole can not reflect the characteristics of later larvae stages.
Acknowledgments
Coming myself from other fields of natural sciences, I found in Federico Bolaños from the Universidad de Costa Rica a colleague and friend who motivated me to start the present and other herpetological studies. I am also grateful to my assistant Félix Domingo Bonilla who helped me to rear the different frog populations as eggs, larvae and young froglets. Maribelle Vargas Montero from the Department of Electronic Microscopy of the Universidad de Costa Rica has the valuable participation in this study by preparing and scanning the mouth parts of the described larva. Jay Savage helped me to tighten up the text, and Brian Kubicki helped revise the final English manuscript.
Resumen
Masas de huevos de la rana centrolénida Hyalinobatrachium pulveratum fueron recolectados en cuatro localidades de las tierras bajas de Costa Rica. La ontogénesis de los renacuajos de esta especie está documentada por descripciones periódicas de los estadios larvales. La larva de H. pulveratum cambia sus proporciones anatómicas durante el desarrollo debido al crecimiento desigual de algunas partes del cuerpo. Debido a estos cambios las larvas en los estadios tempranos del desarrollo difieren considerablemente de aquellos de los estadios tardíos. Las larvas jóvenes tienen un corte transversal casi circular, mientras que las larvas mayores cambian a la forma plana típica. Algunas otras características morfológicas como el ancho de la distancia interorbital y el ancho del disco oral cambian pero no proporcionalmente con la longitud de la larva. Mediciones numerosas de los diferentes tamaños larvales ayudan a reconocer las larvas de H. pulveratum en todos los estadíos. La característica ventral más conspicua es el sino hiobranquial rojo y brillante que cubre ventralmente el aparato hiobranquial. Se discute el uso de valores alométricos y de los estudios ontogénicos.
References
Cadle, J.E. & R.W. Mcdiarmid. 1990. Two new species of Centrolenella (Anura: Centrolenidae) from north-western Peru. Proc. Biol. Soc. Wash. 103: 746-768. [ Links ]
Duellman, W.E. 2001. The Hylid Frogs of Middle America. Vol.1. Soc. Study Amphibians and Reptiles, ISBN: 0-916984-56-7. [ Links ]
Gosner, K.L. 1960. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 16: 183-190. [ Links ]
Ibañez, R., F.E. Jaramillo & C.A. Jaramillo. 1999. Amplificación del ámbito de distribución y descripción del renacuajo de la rana de cristal Hyalinobatrachium aureoguttatum (Anura: Centrolenidae). Rev. Acad. Colombiana. 22: 293-298. [ Links ]
Kubicki, B. 2004. Rediscovery of Hyalinobatrachium chirripoi (Anura: Centrolenidae) in southeastern Costa Rica. Rev. Biol. Trop. 52: 215-218. [ Links ]
Jaramillo, F.E., C.A. Jaramillo & R. Ibañez. 1997. Renacuajos de la rana de cristal Hyalinobatrachium colymbiphyllum (Anura: Centronelidae). Rev. Biol. Trop. 45(2): 867-870. [ Links ]
McDiarmid, R.W. & R. Altig. 1999. Tadpoles. The Biology of Anuran Larvae. Univ. Chicago, Chicago and London. [ Links ]
Nodzenski, E., R.J. Wassersug & R.F. Inger. 1989. Developmental differences in visceral morphology of megophryine pelobatid tadpoles in relation to their body form and mode of life. Biol. J. Linnean Soc. 38: 369-388. [ Links ]
Savage, J.M. 2002. The Amphibians and Reptiles of Costa Rica. A Herpetofauna Between Two Continents, and Two Seas. University of Chicago, Chicago, USA. 934 p. [ Links ]

