










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.51 n.3-4 San José Sep. 2003
in mouse, rat, and hamster astrocytes
Adriana Troyo & Misael Chinchilla
Centro de Investigación en Enfermedades Tropicales, Departamento de Parasitología, Facultad de Microbiología, Universidad de Costa Rica, 2060 San José, Costa Rica; address: atroyo@cariari.ucr.ac.cr
Received 10-IV-2002. Corrected 26-VIII-2003. Accepted 29-VIII-2003.
Abstract
The infection and multiplication of Toxoplasma gondii and Trypanosoma cruzi were compared in primary cultures of white rat, mouse and hamster astrocytes. These cells were cultured on cover slides and infected with T. gondii tachyzoites or T. cruzi blood trypomastigotes. Results show that hamster astrocytes are more susceptible to the multiplication of both parasites than rat and mouse cells. There was no statistical difference between the T. gondii infection in rat and mouse astrocytes (p<0.05), and this suggests an important role of other mechanisms or cells in the white rat natural resistance to this parasite. Because the hamster astrocytes are less resistant to these parasites multiplication and not necessarily to the invasion, any difference observed could be due to an intracellular effect: hamster brain astrocytes favor survival and multiplication of these parasites.
Key words: Toxoplasma gondii, Trypanosoma cruzi, cell culture, astrocyte, rat, mouse, hamster.
Toxoplasma gondii and Trypanosoma cruzi are parasites of great importance in public health. In immunocompetent hosts, T. gondii infection may cause chorioretinitis, abortion or nervous system problems in the newborn, and T. cruzi infection causes Chagas disease, which involves chronic problems such as cardiomyopathy. However, in the immuno-suppressed host, both parasites may provoke death from severe central nervous system infection (Tanowitz et al. 1992, McCabe and Chirurgi 1993).
T. gondii and T. cruzi are obligate parasites capable of infecting and multiplying in cells of different animals, including humans (Dubey 1987, Tanowitz et al. 1992). This has been demonstrated by in vivo and in vitro experiments using different animal species (Kaufman and Maloney 1962, Braveny et al. 1978, Chinchilla et al. 1981a, Shepp et al. 1985, Jones et al. 1986, Chinchilla et al. 1995, 1996).
For T. cruzi as well as for T. gondii investigations have shown that each animal species has variable susceptibility to the infection. For example, in a study done with white rats, it was demonstrated that this animal is highly resistant to T. gondii infection, and that its susceptibility depends on the age of the rat (Chinchilla et al. 1981a, 1996). The multiplication patterns of these parasites have also been compared in vitro in macrophages of different animals. In these cases also, white rat macrophages were more resistant to infection than mouse, and hamster macrophages (Chinchilla et al. 1981b, 1995).
Both of these parasites are able to infect different cell types, including cells from the nervous system. Research on the in vitro infection of brain cells and specially on the behavior of T. gondii in these cells is relevant, considering the increase in disseminated and central nervous system infections that occur in immunosuppression (Jones et al. 1986, Fagard et al. 1999).
Even though there have been some investigations using brain cells of different animals, the infection of T. gondii and T. cruzi in astrocytes from different host species has not been thoroughly compared. Therefore, we have designed a model in order to study and compare the multiplication pattern of T. gondii and T. cruzi in primary culture of astrocytes from different rodents. This could be used as the basis for future research including immunity studies in mouse, rat and hamster astrocytes.
Materials and methods
Animals: White NGP mice, Sprague Dawley rats, and golden Syrian hamsters, Mesocricetus auratus, were used in the experiments. They were fed with a locally produced concentrate (Harrigan Mix) and water ad libitum. Rodents of less than 3 days old were used for the cell cultures.
Parasites: T. gondii RH strain maintained by twice weekly passages in NGP mice (20- 25g), and T. cruzi TC-2 strain isolated in Costa Rica and maintained by passages once a month in C3 H mice (20g) were used.
Cell cultures: The cell cultures were worked separately using the brains of mice, rats and hamsters according to the method described by Ponten (1973) with some modifications. Briefly, the animals of less than three days old (approximately 8 animals for mice or hamsters, and 4 for rats) were anesthetized until death with ether and soaked with 70º ethyl alcohol. Their heads were separated using sterile scissors and tweezers, and the brains were dissected aseptically. They were placed in Minimal Essential Medium (Sigma) with 10% bovine fetal serum, 100 U/ml of penicillin, and 100 µg/ml of streptomycin (MEM-BFS). Passing the brains through 18, 20 (or 21), and 26 gauge needles, in that order, 3 times each, dissociated the cells. This material was placed in MEM-BFS in 25 cm2 cell culture bottles (or 75 cm 2 if necessary) and incubated at 37º C in a 5% CO2 atmosphere for 8-10 days or until a cell monolayer formed. The medium was replaced whenever necessary.
Infection models: After the primary culture, the cells were washed 3 times with Hanks solution (Sigma) and dispersed by adding 0.25% trypsin and 0.02% EDTA in MEM without bovine fetal serum for 10-15 minutes. The suspension was centrifuged for 8 min at 300 x g, and the resuspended cells in MEM-BFS were subcultured on 22x22 mm cover slides by placing 0.3 ml of the cell suspension on each slide (Chinchilla and Frenkel 1978). By this method, subcultured brain cell monolayers contain more than 95% astrocytes (Fontana et al. 1984, Peterson et al. 1993, Creuzet et al. 1998). The cover slides were incubated at 37ºC in 5% CO2 for 24-48 hours and infected after counting the cells on one of the slides in a Neubauer chamber following trypsinization.
For the infection with T. gondii, the tachyzoites were obtained from peritoneal exudate of an infected mouse, they were counted in a Neubauer chamber, and the inoculum was adjusted to one tachyzoite per cell. The cells were infected and samples were obtained after 1 and 24 hours of incubation at 37ºC in 5% CO2 . The cover slides were dried at room temperature, fixed for 5 minutes with methanol, stained for 30 minutes with Giemsa, dried, and mounted with Permount.
The trypomastigotes used in the infections with T. cruzi were obtained according to the method described by Budzko et al. (1975). Briefly, total heparinized blood of an infected mouse was stratified on Ficoll Histopaque-1077 (Sigma), centrifuged at 400 x g for 40 min at room temperature, and the mononuclear cells and trypanosome layer was separated. The trypomastigotes were counted, the cells infected, and cover slide samples taken at 24, 96, 120 and 144 hours after incubation, treating and staining them as described for T. gondii.
For each cover slide infected with T. gondii, 500-700 astrocytes were counted, and 700-1200 astrocytes for those infected with T. cruzi. The number of parasites per 100 total cells and per 100 infected cells was determined, and multiplication rates were obtained with these data: 24hr/1hr for T. gondii and 96hr/24hr, 120hr/24hr, and 144hr/24hr for T. cruzi.
All the experiments were repeated 5 times and the data was analyzed using the Students t-test to determine the differences between the means (a:0.05) (Hill 1966).
Results
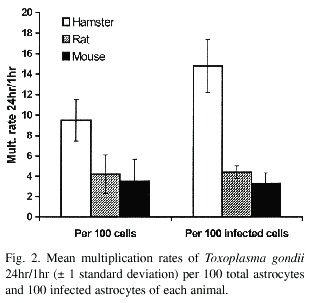
The multiplication curve for T. gondii in the astrocytes of each animal species is shown in Figs. 1 and 2. According to these results, less than 15% of the tachyzoites penetrated the astrocytes of these rodents in an hour or less. In the rat, less than 5% of the cells are infected after 1 hour of incubation. The multiplication rates of T. gondii 24hr/1hr for 100 total cells and 100 infected cells are presented in Fig. 2.
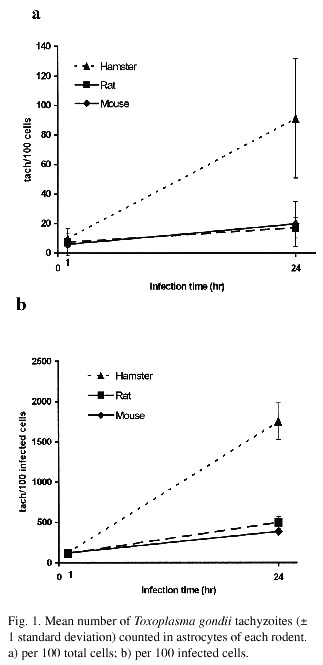
When comparing the three rodents, the T. gondii infection after 1 hour is similar in all of them. However, the parasite multiplication was higher in the hamster astrocytes because the number of tachyzoites after 24 hours of infection is evidently greater for 100 total cells (Fig. 1a) as well as 100 infected cells (Fig. 1b). The statistical analysis indicates a significant difference (p<0.05).
The same result is obtained when the multiplication rates of Toxoplasma (24hr/1hr) between the different animal astrocytes are compared. The multiplication rates in the hamster cells are greater for 100 total and infected cells (p<0.05). The multiplication rates for mouse and rat cells were similar, and the difference between both rodents is not statistically significant (Fig. 2).
The T. cruzi multiplication curves for each rodent are shown in figures 3, and 4. For this flagellate, the infection after 24 hours in most experiments is less than 10 parasites per 100 cells, and in the case of the rat astrocytes, this percentage is much lower.

In all the astrocyte cells infected with T. cruzi, only amastigotes were found after 24 and 96 hours, but other forms of the parasite such as epimastigotes and trypomastigotes were present at 120 hours post-infection (in lower proportion than amastigotes). At 144 hours of infection there were less amastigotes than epimastigotes and trypomastigotes in the cells of the three animals studied.
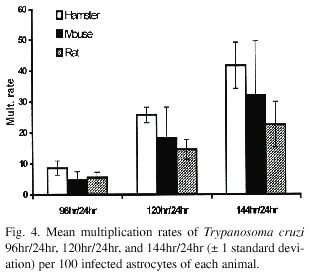
When comparing the multiplication of T. cruzi in the astrocytes, the number of parasites per 100 total cells and 100 infected cells in hamster differs significantly from the rat cells (p<0.05). The hamster astrocytes are more susceptible whereas the rat astrocytes are more resistant to the parasite multiplication (Fig. 3).
The trypanosome 96hr/24hr and 120hr/24hr multiplication rates for hamster and rat astrocytes are also statistically different (p<0.05): they are greater for the hamster cells than for the rat cells. For the mouse astrocytes, the multiplication rate is intermediate (Fig. 4).
Discussion
The multiplication of T. gondii in cell cultures has been studied using cells of different origin. It has been studied in cells such as microglial cells, astrocytes, and even neurons, along with the effect of different immune factors on the infection (Jones et al. 1986, Peterson et al. 1993, Chao et al. 1993, Fagard et al. 1999, Freund et al. 2001).
It has been reported that strains of T. gondii behave differently when infecting the same cell type (Kaufman and Maolney 1962), and it is also known that animals such as the white rat are less susceptible to T. gondii (Chinchilla et al. 1981a). Experiments done in vivo with white rats, wild rats (Sigmodon hispidus), mice, and hamsters show that the white rat is more resistant to a Toxoplasma infection compared to mice and hamsters (Chinchilla et al. 1996). On the other hand, and according to in vivo and in vitro studies, the white rat peritoneal macrophages play a very important role in this animal natural adaptation to the parasite, apparently due to enzymatic factors that prevent the development of T. gondii within the macrophage (Chinchilla et al. 1981a, 1981b).
In this study, the in vitro infection and multiplication of Toxoplasma in rat and mouse astrocytes were similar, further supporting the hypothesis that the natural resistance of the white rat is due to other cells or mechanisms, such as the action of the peritoneal macrophages. However, there is a difference regarding the hamster astrocytes, which are evidently more susceptible to this parasite. In reports of other in vitro (macrophages) and in vivo infections, the similarity was between mice and hamsters (which differ from rats), mostly due to the mentioned activity of the rat macrophages (Chinchilla et al. 1995, 1996).
Also, the number of tachyzoites per 100 infected cells after 1 hour was similar for all the animals studied. Therefore, hamster astrocytes are less resistant to Toxoplasma than mouse and rat astrocytes in vitro, and the difference observed in the parasite multiplication in these cells is due to intracellular phenomena, rather than a deficiency in the organism invasion capacity.
As it has been mentioned, the macrophages of the rat are more resistant to T. gondii than those of other rodents, and it has also been observed that they are more resistant to other intracellular parasites like T. cruzi (Chinchilla et al. 1995). This experiment shows that in vitro, rat astrocytes are also less susceptible to the multiplication of this flagellate than the other rodents, but its entrance into the cell is similar in white rats, mice, and hamsters. Mouse astrocytes present an intermediate resistance, while hamster cells are the least resistant.
When comparing the multiplication rates of T. cruzi and T. gondii in the cells of the different animals, the hamster astrocytes again show to be the most favorable ones for the parasites. Therefore, apparently hamster astrocytes present an intracellular environment that favors their permanence and multiplication more than the rat and mouse cells. This could be due to factors that have a similar intracellular effect for both parasites, as it has been previously suggested for cotton rat (S. hispidus) macrophages (Chinchilla et al. 1995).
With these results, the difference that animals and host cells can show in an infection by an intracellular parasite is even more obvious. The work on intracellular multiplication of parasites that many times is carried out in vitro does not contemplate the differences that can exist between cells of different origin, animals whose characteristics may vary, or differences in human or cell lines. Even cell subpopulations in one organism may possess characteristics and express genes differently, limiting the intracellular parasite multiplication (Moulder 1985, Crocker et al. 1987).
By comparing the multiplication of T. gondii and T. cruzi in astrocytes of different rodents, a base study is established to continue the work comparing these cells. In vitro investigations have been done with astrocytes and microglial cells, protecting the cells from infection with cytokines like interferon g and tumor necrosis factor a (Peterson et al. 1993, Chao et al. 1993, Halonen and Weiss 2000, Freund et al. 2001), but most of these have been done in mice. This study serves as a basis to compare the effect of cytokines obtained from different animals and evaluate their specificity according to parasite and host species, as it has been done with these two parasites in other non-phagocytic cells (Chinchilla and Frenkel 1978, 1984, Reyes and Chinchilla 1987, Chinchilla and Reyes 1990, Chinchilla et al. 2003). Therefore, it presents the possibility of using astrocytes of various rodents, considering the differences in susceptibility, to determine important host factors that influence the parasite multiplication.
Acknowledgements
This work was supported by the Vicerectoría de Investigación, Universidad de Costa Rica, project # 803-97-262.
Resumen
Se comparó la infección y multiplicación de Toxoplasma gondii y Trypanosoma cruzi en cultivos celulares primarios de astrocitos, a partir de células de cerebro de rata blanca, ratón y hámster. Las células fueron cultivadas en cubreobjetos e infectadas con taquizoitos de T. gondii o tripomastigotos sanguíneos de T. cruzi. Los resultados muestran que los astrocitos de hámster son más susceptibles que los de rata y ratón a la multiplicación de ambos parásitos. No se encontró diferencia estadísticamente significativa entre la infección por T. gondii en los astrocitos de ratón y rata (p<0.05), lo que sugiere la importancia de otros mecanismos o células en la resistencia natural de la rata blanca a esta parásito. Al ser los astrositos de hámster menos resistentes a la multiplicación de estos parásitos y no necesariamente a la invasión, cualquier diferencia observada podría deberse a un fenómeno intracelular: los astrocitos de hámster favorecen la permanencia y multiplicación de estos parásitos.
References
Braveny, I., W. Winter & R. Disko. 1978. A method of mass cultivation of Toxoplasma gondii in cell culture. Tropenmed. Parasit. 29: 432-434. [ Links ]
Budzko, D.B., M.C. Pizzimenti & F. Kierszenbaum. 1975. Effects of complement depletion in experimental Chagas disease: immune lysis of virulent blood forms of Trypanosoma cruzi. Infect. Immun. 11: 86- 91. [ Links ]
Chao, C.C., S. Hu, G. Gekker, W.J. Novick Jr., J.S. Remington & P.K. Peterson. 1993. Effects of cytokines on multiplication of Toxoplasma gondii in microglial cells. J. Immunol. 150: 3404-3410. [ Links ]
Chinchilla, M. & J.K. Frenkel. 1978. Mediation of immunity to intracellular infection (Toxoplasma and Besnoitia) within somatic cells. Infect. Immun. 19: 999-1012. [ Links ]
Chinchilla, M., M. Alfaro & O.M. Guerrero. 1981a. Adaptación natural de la rata blanca a Toxoplasma gondii. Rev. Biol. Trop. 29: 273-282. [ Links ]
Chinchilla, M., O.M. Guerrero, L. Reyes & E. Abrahams. 1996. Susceptibilidad de la rata Sigmodon hispidus (Rodentia: Cricetidae) al Toxoplasma gondii (Eucoccidia: Sarcocystidae). Rev. Biol. Trop. 44: 265-268. [ Links ]
Chinchilla, M., O.M. Guerrero & E. Solano. 1981b. Acción de los macrófagos de la rata blanca contra Toxoplasma gondii "in vitro". Rev. Latinoam. Microbiol. 23: 239-243. [ Links ]
Chinchilla, M. & J.K. Frenkel. 1984. Resistencia a la infección por Toxoplasma in vitro inducida por mediadores linfocitarios en células no fagocíticas de ratón y hámster. Inmunología 3: 144-150. [ Links ]
Chinchilla Carmona, M., M. Herrera Corrales, O.M. Guerrero Bermúdez, A. Jiménez Somarribas, G. Tamayo, A. Sittenfeld Appel, V. Nielsen & P. Hurtado. 2003. Efecto de extractos de artrópodos sobre la multiplicación del Toxoplasma gondii dentro de macrófagos peritoneales de ratón. Rev. Biol. Trop. 51: 317-332. [ Links ]
Chinchilla, M. & L. Reyes. 1990. Characteristics of the lymphokine active against multiplication of T. cruzi in mouse heart cells. Infect. Immun. 9: 75-82. [ Links ]
Chinchilla, M., L. Reyes & O.M. Guerrero. 1995. Resistance to intracellular parasites correlates with species differences in ability of macrophages to inhibit parasite replication. Immun. Infect. Dis. 5: 83-87. [ Links ]
Creuzet C., F. Robert, M. P. Roisin, H. Van Tan, C. Benes, J. Dupouy-Camet & R. Fagard. 1998. Neurons in primary culture are less efficiently infected by Toxoplasma gondii than glial cells. Parasitol. Res. 84: 25-30. [ Links ]
Crocker, P.R., E.V. Davies & J.M. Blackwell. 1987. Variable expression of the murine natural resistance gene Lsh in different macrophage populations infected in vitro with Leishmania donovani. Parasite Immunol. 9: 705-719. [ Links ]
Dubey, J.P. 1987. Toxoplasmosis. Vet. Clinics North. Am.: Small Animal Practice. 17: 1389-1404. [ Links ]
Fagard, R., H. Van Tan, C. Creuzet & H. Pelloux. 1999. Differential development of Toxoplasma gondii in neural cells. Parasitology Today. 15: 504-507. [ Links ]
Freund, Y.R., N.T. Zaveri & H.S. Javitz. 2001. In vivo investigation of host resistance to Toxoplasma gondii infection in microglia of BALB/c and CBA/Ca mice. Infect. Immun. 69: 765-772. [ Links ]
Fontana, A., W. Fierz & H. Wekerle. 1984. Astrocytes present myelin basic protein to encephalitogenic T-cell lines. Nature 307: 273-276. [ Links ]
Halonen, S.K. & L.M. Weiss. 2000. Investigation into the mechanism of gamma interferon-mediated inhibition of Toxoplasma gondii in murine astrocytes. Infect. Immun. 68: 3426-3430. [ Links ]
Hill, A.B. 1966. Principles of Medical Statistics. Oxford University, New York. [ Links ]
Jones, T.C., K.A. Bienz & P. Erb. 1986. In vitro cultivation of Toxoplasma gondii cysts in astrocytes in the presence of gamma interferon. Infect. Immun. 51: 147- 156. [ Links ]
Kaufman, H.E. & E.D. Maloney. 1962. Multiplication of three strains of Toxoplasma gondii in tissue culture. J. Parasitol. 48: 358-361. [ Links ]
McCabe, R. & V. Chirurgi. 1993. Issues in Toxoplasmosis. Infect. Dis. Clin. North America. 7: 587-604. [ Links ]
Moulder, J.W. 1985. Comparative biology of intracellular parasitism. Microbiol. Rev. 49: 298-337. [ Links ]
Peterson, P. K., G. Gekker, S. Hu & C. C. Chao. 1993. Intracellular survival and multiplication of Toxoplasma gondii in astrocytes. J. Infect. Dis. 168: 1472-1478. [ Links ]
Ponten, J. 1973. Human Glial Cells. In P. Kruse & M.K. Patterson (eds.). Tissue Culture Methods and Applications. Academic, New York. 868 p. [ Links ]
Reyes, L. & M. Chinchilla. 1987. Growth inhibition of Trypanosoma cruzi in cultured murine myocardial cells mediated by a specifically induced lymphokine. Infect. Immun. 55: 1513-1516. [ Links ]
Shepp, D.H., R.C. Hackman, F.K. Conley, J.B. Anderson & J.D. Meyers. 1985. Toxoplasma gondii reactivation identified by detection of parasitemia in tissue culture. Ann. Intern. Med. 103: 218-221. [ Links ]
Tanowitz, H.B., L.V. Kirchhoff, D. Simon, S.A. Morris, L.M. Weiss & M. Wittner. 1992. Chagas disease. Clin. Microb. Rev. 5: 400-419. [ Links ]

