










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.51 n.2 San José Jun. 2003
Carlos Díaz-Avalos 1, Juan Francisco Barba-Torres 2 & Mª Teresa Gaspar-Dillanes 3
Recibido 02-XI-2000. Corregido 27-IV-2001. Aceptado 16-IX-2001
Egg abundance of Anchoa mitchilli was studied in Laguna de Tamiahua, Veracruz during three annual cycles (84-85, 85-86 and 86-87). Our goal was to detect areas and seasons with high egg abundance and the possible association of such areas and seasons with covariates such as temperature, salinity, transparency, depth, location, season and year. The association was tested statistically using a Generalized Linear Model (GLM) with Gamma type error. The results suggest that not all the covariates considered have a significant effect on egg abundance (Students T, p>0.005). The significant effects show the presence of a seasonal component and that the effect of salinity on egg abundance is conditional to location. High average egg abundances for the three annual cycles were observed during summer. The presence of significant interactions (Students T, p<0.05) during the last two annual cycles shows that the variability of egg abundance is associated to surface temperature and salinity, conditionally to year.
Key words: Anchoa mitchilli, Tamiahua, generalized linear models, covariate analysis, seasonal abundance.
Una de las metas primordiales de la investigación biológico-pesquera es generar información que mejore el conocimiento sobre las poblaciones animales que impactan al sector pesquero, ya sea porque están bajo explotación o porque son ecológicamente importantes debido a su papel como alimento para las especies comerciales. Anchoa mitchilli (Cuvier y Valenciennes 1848), es una de las presas más abundantes para las especies de importancia comercial en los sistemas estuarinos del Noreste del continente americano (Purcell et al. 1994, Griffith y Bechler 1995, Ayala et al. 1998). Debido a que A. mitchilli no es capturada comercialmente, las estimaciones de abundancia de la población desovante se hacen de manera indirecta a partir del conteo de huevos (Chavance et al. 1984). El efecto que tienen las variaciones espaciales y temporales de la abundancia de huevos de A. mitchilli repercuten en la calidad de las estimaciones de su abundancia. Es importante por lo tanto que dichas variaciones sean consideradas en el proceso de análisis, al igual que otros factores asociados.
En México, A. mitchilli ha sido descrita como una especie típicamente lagunar (Flores Coto et al. 1983). De acuerdo con varios autores, su abundancia fluctúa temporalmente en las lagunas de Pueblo Viejo (Castillo-Rivera et al. 1994), Tamiahua (Barba-Torres y Sánchez-Robles 1981) y Alvarado (Méndez 1980) en el estado de Veracruz, y en la Laguna de Términos en el estado de Campeche (Ocaña-Luna et al. 1987, Flores Coto et al. 1988).
Los ecosistemas lagunares estuarinos son sistemas dinámicos, lo cual dificulta las explicaciones de los cambios en la abundancia de huevos de A. mitchilli. Sin embargo, cuando se considera su posible asociación con factores temporales y otras variables hidrológicas, es posible aproximarse a la comprensión de éstos fenómenos. Esto se puede lograr a través de pruebas de significancia estadística de las asociaciones observadas. Este tipo de análisis no soslaya a las observaciones emanadas de estudios cualitativos, ya que éstos son útiles para sugerir los factores y covariables posiblemente asociados a la abundancia de las especies animales.
Las variaciones espaciales y temporales en la abundancia de huevos de A. mitchilli han sido relacionadas con cambios en la abundancia de alimento (Pebbles et al. 1996, Fulling y Peterson 1999) y de depredadores como las medusas y ctenóforos (Dorsey et al. 1996, Rilling y Houde 1999). Otros autores han encontrado que la cantidad de huevos de A. mitchilli depredados por las medusas está correlacionado negativamente a la cantidad de oxígeno disuelto, temperatura y salinidad (Castillo-Rivera et al. 1994, Breitburg et al. 1997). En este trabajo se describe la variación espacial y temporal de la abundancia de huevos de A. mitchilli y se verifica, mediante la significancia estadística, la asociación de dicha variación con algunos factores y covariables hidrológicas.
Materiales y métodos
Área de estudio: La Laguna de Tamiahua está localizada al Norte del Estado de Veracruz, en la costa Noreste de México (21°1224 y 22°0000 N, 97°1912 y 97°4616 W). Es alargada (93 km por 21.5 km) y somera, con profundidad promedio de 3.0 m hacia su parte central. Una barra arenosa, llamada Cabo Rojo, la separa parcialmente del Golfo de México (Ayala-Castañares et al. 1969). Esta barra se encuentra cortada en su porción Norte por la Boca de Tampachichi de origen artificial y al Sur por la Boca de Corazones de origen natural. El aporte fluvial más importante que recibe la Laguna proviene de varios esteros. Aunque la mayoría de ellos tiene flujo estacional, todos ellos influyen en las condiciones hidrológicas de la Laguna. El clima en la zona es subhúmedo, lluvioso en verano y seco en invierno, correspondiendo al tipo Aw 0 2(e) de acuerdo con García (1973), el cual es modificado por las fuertes corrientes o vientos dominantes del Norte durante otoño e invierno.
La laguna de Tamiahua ha sido descrita como un depósito constante de agua de tipo estuárico, oligohalino, que temporalmente se ve influenciado por los aportes de agua continental y por el influjo de las aguas neríticas que penetran a través de la Boca de Corazones (Villalobos et al. 1968). Estas condiciones han cambiado debido a la apertura de la Boca Tampachichi en el norte de la Laguna, la cual permite la entrada de aguas de tipo ultrahalinas (Gutiérrez y Contreras 1987). A lo largo de cada ciclo anual se distinguen dos épocas climáticas determinantes en el comportamiento hidrológico: una denominada de "secas" que se registra de marzo a agosto, y la otra de "lluvias", la cual abarca de septiembre a febrero. En la Primavera se presentan valores altos de temperatura y salinidad, debido al aumento de la insolación propia de ésta época y a la ausencia de aporte de agua dulce, elevándose la concentración de sales. A partir del mes de septiembre (otoño) se presenta una drástica disminución de temperatura y salinidad como consecuencia del aumento en la precipitación y a la presencia de los fenómenos metereológicos denominados "nortes".
Metodología: Se llevaron a cabo 12 visitas a la zona de estudio, en el período comprendido entre octubre de 1984 y agosto de 1987. En cada una de ellas se realizaron arrastres de ictioplancton utilizando una red cónica tipo trapecio, con diámetro de boca de 0.5 m, luz de malla de 500 micras y manga (longitud de la red) de 1.5 m. Cada arrastre tuvo una duración de 5 minutos en un total de 16 estaciones de recolecta, distribuidas como se muestra en la Fig. 1. Los ejemplares recolectados están depositados en el museo del Instituto de Biología de la Universidad Nacional Autónoma de México con los números de catalogo IBUNA-P2752 e IBUNA-P2759.
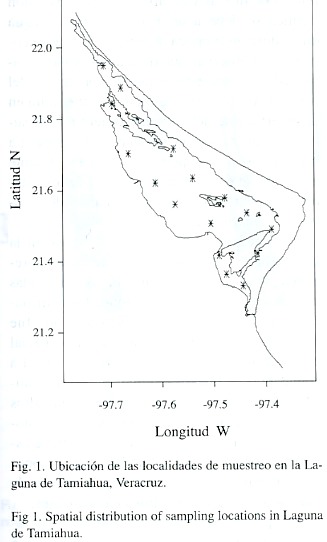
Para cuantificar el agua filtrada por la red, se le adaptó un contador de flujo previamente calibrado. Con los valores obtenidos del número de huevos y la cantidad de agua filtrada se obtuvo una estimación local de la densidad de huevos por metro cúbico de agua. En cada visita a las estaciones de muestreo fueron medidos y registrados los datos de las covariables de temperatura, salinidad y oxígeno disuelto tanto superficial como de fondo. Asimismo, fue medida la transparencia con un disco de Secchi. Además de las covariables anteriores, los factores considerados en el estudio son la época del año, el ciclo anual y la localización geográfica del punto de muestreo.
Con el propósito de caracterizar el tipo de asociación, lineal o no lineal, entre la abundancia de huevos y los factores hidrológicos considerados en este estudio, así como otras propiedades de los datos, se utilizaron diversas técnicas de análisis exploratorio de datos (Tuckey 1977). Los resultados de este análisis sugirieron que la abundancia de huevos y su varianza son proporcionales, así como la existencia de interacciones entre los factores hidrológicos. Para verificar estadísticamente la asociación entre los factores y covariables hidrológicas y la abundancia de huevos de A. mitchilli se ajustó un modelo lineal generalizado (MLG). Los MLG son una extensión de los modelos lineales ordinarios, los cuales permiten considerar diferentes estructuras probabilísticas para el término aleatorio. En los MLG se considera un vector de observaciones o respuestas y, el cual se supone es una muestra aleatoria independiente e idénticamente distribuida de una población cuya distribución de probabilidad pertenece a la familia exponencial (Nelder y Wedderburn, 1972). El valor esperado de y se relaciona con un conjunto de k variables explicativas a través de un predictor lineal 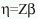 que representa la parte sistemática del modelo, donde Z es una matriz de covariables y factores y
que representa la parte sistemática del modelo, donde Z es una matriz de covariables y factores y 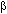 es un vector de coeficientes por estimar. El estimador lineal se relaciona al valor esperado de y por medio de la función liga:
es un vector de coeficientes por estimar. El estimador lineal se relaciona al valor esperado de y por medio de la función liga:
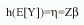
en donde h es una función derivable. En nuestro estudio, la respuesta (abundancia de huevos de A. mitchilli) es una cantidad no negativa, por lo que se escogió una estructura tipo Gamma, es decir,
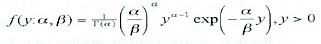
donde es conocida como función Gamma (Abramowitz y Stegun 1974).
El modelo ajustado a los datos incluyó los efectos de temperatura superficial (TESUP), temperatura de fondo (TEFON), salinidad de superficie (SASUP), salinidad de fondo (SAFON), los factores: época y año. Estas dos últimas variables son categóricas (factores), por lo que se utilizaron contrastes de suma a fin de garantizar la identificación de sus efectos. Se consideró también la posible existencia de interacciones de primer orden entre las covariables (Montgomery 1984) y se utilizó una función liga logarítmica. De esta manera, el modelo inicial fue:
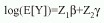
donde es el vector de coeficientes relacionados a los efectos principales y 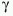 es el vector de coeficientes relacionados a las interacciones de primer orden. Con el propósito de obtener un modelo parsimonioso, se aplicó una rutina de selección de variables en ambos sentidos. Las pruebas de hipótesis sobre la significancia de los parámetros en el modelo se hicieron comparando los cambios en la devianza con una distribución
es el vector de coeficientes relacionados a las interacciones de primer orden. Con el propósito de obtener un modelo parsimonioso, se aplicó una rutina de selección de variables en ambos sentidos. Las pruebas de hipótesis sobre la significancia de los parámetros en el modelo se hicieron comparando los cambios en la devianza con una distribución 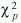 (Mc Cullagh y Nelder 1989), donde los grados de libertad corresponden a la diferencia en el número de parámetros en los modelos comparados.
(Mc Cullagh y Nelder 1989), donde los grados de libertad corresponden a la diferencia en el número de parámetros en los modelos comparados.
Resultados
En el Cuadro 1 se presenta un resumen por época de los valores de temperatura y salinidad, superficial y de fondo. Durante la temporada de secas, la temperatura promedio en la superficie del agua, siempre fue mayor que la del fondo y menor que la del fondo en la temporada de lluvias y en invierno. Esta inversión térmica se debe a un mayor aporte de agua dulce durante la época de lluvias, lo cual se corrobora al observar que durante el otoño la salinidad en la superficie fue menor que la del fondo. En el invierno, la menor temperatura en la superficie se debe al enfriamiento de la superficie que provocan los vientos del norte. La baja salinidad de superficie observada durante el verano se debe al aporte fluvial de los esteros como el Cucharas en las cercanías de la estación número siete.
Los resultados en el Cuadro 1 revelan la presencia de una componente estacional, apreciándose en los valores más bajos para todas las covariables durante el otoño. La distribución espacial de estos factores no fue uniforme, presentándose un patrón espacial bien definido para cada estación del año. La abundancia de huevos también sigue este comportamiento estacional, con valores más altos durante el verano y más bajos durante el otoño. Esto es indicativo de la alta variabilidad espacial y temporal de la abundancia, con zonas en la laguna de alta y baja concentración de huevos a lo largo de los tres ciclos anuales.
El patrón espacial observado de temperatura y salinidad esta asociado al aporte de agua dulce de los ríos y a la entrada de agua marina en las bocas de Tampachichi y de Corazones (Flores-Coto et al. 1983), ya que es cerca de estas áreas donde se localizaron las mayores fluctuaciones. Esto se muestra en la Fig. 2, donde se presentan los mapas que relacionan las temperaturas y abundancias de los huevos promedio para las cuatro estaciones del año. La escala de gris corresponde a la temperatura promedio de la época, mientras que los contornos representan isopletas de abundancia de huevos.
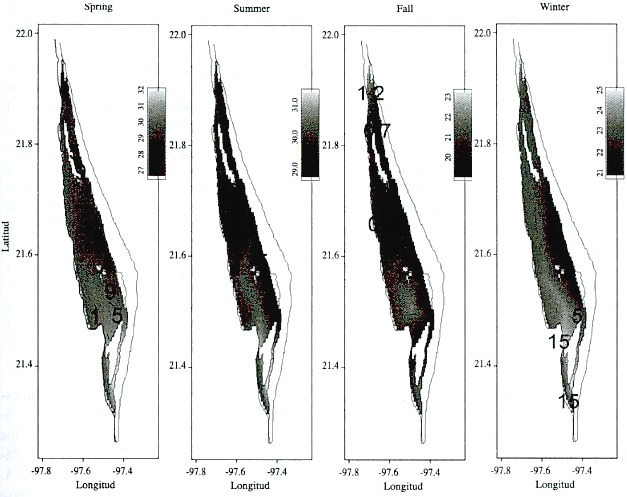
Fig. 2. Mapas promedio de temperatura superficial (escala de gris) y de abundancia de huevos de A. mitchilli (isopletas) para las cuatro estaciones del año en la Laguna de Tamiahua.
Fig. 2. Average temperature (gray scale) and egg abundance (isoplets) maps for the four seasons in Laguna de Tamiahua.
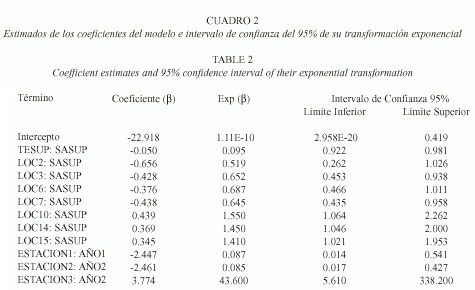
Respecto a la distribución general de la abundancia de los huevos, en la primavera ocurrieron las mayores abundancias de huevos en la parte este de la laguna, mientras que los picos de abundancia se presentaron en la parte central de la laguna durante el verano. En el invierno, las áreas de mayor abundancia de huevos se encontraron en la parte sur de la laguna. Esto implica que las abundancias siempre ocurren asociadas de manera condicional, para cuantificar esta relación se ajustó el modelo lineal general, de acuerdo a estas características. El ajuste del modelo a las condiciones del sistema se presentan en el Cuadro 2, la que contiene a los coeficientes de aquellos términos que resultaron ser significativos (t de Student, p<0.05). El modelo final incluye los efectos principales de temperatura y salinidad de superficie, localidad, época y año, así como las interacciones entre localidad y salinidad de superficie y entre época y año. En este caso, aunque los efectos principales están en el modelo, los coeficientes de sus efectos principales no se pueden interpretar de manera independiente (Montgomery 1984) y no se presentan en el cuadro.
La presencia de interacciones significativas en el modelo, implica que las variables en él incluidas, no están relacionadas con la abundancia de los huevos de la anchoa de manera independiente. La estructura del modelo propuesto implica que las covariables tienen un efecto multiplicativo sobre el valor esperado de la abundancia relativa de los huevos. El signo de los coeficientes de la interacción entre la salinidad superficial y la localidad implica que la asociación entre estas variables es distinta en la parte norte y sur de la laguna. Así, un incremento de una parte por mil en la salinidad en la localidad dos está asociado a una disminución del 51.9% en el valor esperado de la abundancia de huevos con respecto a la media global, mientras que un cambio de la misma magnitud en la localidad diez está asociado a un incremento del 55% en el valor esperado de la abundancia. Los resultados en el Cuadro 2 también confirman la presencia de un efecto estacional en la abundancia de huevos de la anchoa, aunque la magnitud de dicho efecto no fue el mismo en los 3 años debido a la variabilidad del desove y a las condiciones ambientales.
Discusión
Aunque en otros sistemas lagunares estuarinos de México han registrado que la abundancia de A. mitchilli no presenta un patrón espacial definido ni asociación con factores ambientales (Castillo-Rivera et al. 1994), los resultados del modelo lineal ajustado a los datos indican que la abundancia de huevos de A. mitchilli presenta un patrón espacial y temporal significativo, el cual está asociado a la distribución espacial de la temperatura y salinidad superficial. Durante la primavera, las mayores abundancias de huevos se encontraron en áreas al este y noreste de la laguna, mientras que en el verano la abundancia fue mayor en la parte centro y sur de la laguna. En el otoño la abundancia fue baja en general, la cual estuvo asociada a bajas en la salinidad y temperatura superficiales. En esta época se notó una mayor presencia de medusas Chrysaora plocamia, Bougainvillia niobe y Blackfordia virginica, especies registradas como depredadoras de huevos y larvas de A. mitchilli en aguas estuarinas (Barba-Torres y Sánchez-Robles 1981, Breitburg et al. 1997, Rilling y Houde 1999). Con la información obtenida no es posible afirmar si la baja abundancia observada durante el otoño fue debida a una alta tasa de depredación, aunque Govoni y Olney (1991) obtienen que las variaciones estacionales de la abundancia de huevos de A. mitchilli y de sus depredadores gelatinosos son muy similares. Otra posible explicación que deberá ser considerada es que A. mitchilli disminuye su desove ante la presencia de condiciones no propicias para el desarrollo de sus huevos, siendo esta una adaptación de la especie, ajustando la magnitud de sus desoves a las condiciones variables del sistema (Pebles et al. 1996). Finalmente durante el invierno las mayores abundancias se observaron en la parte sur de la laguna. El comportamiento estacional de la abundancia de huevos de A. mitchilli ha sido observado en la mayoría de los estuarios del este de América (Vouglitois et al. 1987, Govoni y Olney 1991, Castillo-Rivera et al. 1994, Griffith y Bechler 1995), aunque la presencia de los picos de abundancia son diferentes a los encontrados en la Laguna de Tamiahua, siendo probable que exista un cambio de abundancia latitudinal.
Aún cuando las fluctuaciones estacionales de A. mitchilli fueron muy grandes, a lo largo de este estudio siempre se encontraron huevos de esta especie, lo que sugiere que es una especie típicamente lagunar, lo que ha sido mencionado por Barba-Torres y Sánchez-Robles (1981), Flores-Coto et al. (1983) y Ayala et al. (1998). Morton y Moran (1989) registran que el desove de A. mitchilli ocurre a temperaturas mayores de 12 ºC y en salinidades superiores a 10 ppm. En la Fig. 2, se aprecia que la mayor abundancia de huevos se encontró entre temperaturas de 24 a 29 ºC.; así mismo, los huevos se registran en un intervalo de salinidades de 7 a 36 ppm, ocurriendo las mayores abundancias entre 14 y 20 ppm (aguas mesohalinas). Con base en estas observaciones, la presencia continua de huevos de A. mitchilli en la Laguna de Tamiahua durante el período de estudio es explicada en parte por las condiciones de temperatura y salinidad, ya que estos dos factores se mantienen todo el año por encima de los valores sugeridos como críticos por Morton y Moran (1989). Con el modelo propuesto es posible determinar que la abundancia es distinta en la parte norte y sur de la laguna, estando la abundancia asociada a los factores y covariables de manera condicional y multiplicativa. Los resultados obtenidos muestran que en la Laguna de Tamiahua, A. mitchilli desova en facies que van desde las oligohalinas, mesohalinas hasta las ultrahalinas.
Agradecimientos
Agradecemos a Samuel Gómez-Aguirre y a Faustino Zavala-García por sus comentarios y criticas al manuscrito. Se agradece el apoyo logístico proporcionado por el Departamento de Biología de la Facultad de Ciencias de la UNAM. El primer autor fue apoyado por el proyecto CONACYT Número 32256E.
Resumen
Se estudió la abundancia de huevos de Anchoa mitchilli (Cuvier y Valenciennes 1848) en la laguna de Tamiahua, Ver. durante tres ciclos anuales (84-85, 85-86 y 86-87), para determinar zonas y épocas de alta concentración de huevos y su posible asociación con variables como temperatura, salinidad, transparencia y profundidad, así como con factores espacio-temporales como posición geográfica, época climática y el ciclo anual de recolecta. Para probar estadísticamente la asociación de la abundancia de huevos con dichos factores y variables se ajusto un Modelo Lineal Generalizado (MLG) con error tipo Gamma. Los resultados del modelo ajustado sugieren que no todas las variables tienen un efecto significativo sobre la abundancia de huevos (t de Student, p >0.05). Las variables que resultaron estadísticamente significativas indican la presencia de un componente estacional y que la salinidad tiene un efecto condicional a la localidad. Las mayores abundancias promedio durante los tres ciclos de recolecta ocurrieron siempre durante el verano. Asimismo, debido a la presencia de interacciones significativas (t de Student, p<0.05) durante los dos últimos ciclos anuales, se puede afirmar que la variabilidad de la abundancia de los huevos de A. mitchilli está asociada a la temperatura y a la salinidad superficiales condicionalmente al año.
Referencias
Abramowitz, M. & Stegun, I. 1974. Handbook of mathematical functions, with formulas, graphs and mathematical tables. Wiley. Nueva York. 1046 p. [ Links ]
Ayala Pérez, L.A., O.A. Alatriste & J. L. Rojas Galaviz, 1998. Estructura de la comunidad de peces en el sistema Candelaria-Panlau, Campeche, México. Rev. Biol. Trop. 46:763-774. [ Links ]
Ayala-Castañares, A., R. Cruz, A. García-Cubas & L.R. Segura. 1969. Síntesis de los conocimientos sobre la Geología Marina de la Laguna de Tamiahua, Veracruz, México, pp. 39-48 In A. Ayala-Castañares & F.B. Phleger (eds.). Lagunas Costeras, Un Simposio. Mem. Simp. Intern. Lagunas Costeras. UNAM, UNESCO. México, D.F. [ Links ]
Barba-Torres, J.F. & J. Sánchez-Robles. 1981. Abundancia, Distribución y Estructura de la Comunidad Ictioplanctónica en la Laguna de Tamiahua, Veracruz, a través de un Ciclo Anual. Tesis de Licenciatura, Universidad Nacional Autónoma de México, México D.F., México. [ Links ]
Breitburg, D.L., T. Loher, C.A. Pacey & A. Gerstein. 1997. Varying effects of low dissolved oxygen on trophic interactions in an estuarine food web. Ecol. Monogr.67: 489-507. [ Links ]
Castillo-Rivera, M., G. Moreno & R. Iniestra. 1994. Spatial, seasonal, and diel variation in abundance of the bay anchovy, Anchoa mitchilli (Teleostei: Engraulidae), in a tropical coastal lagoon of Mexico. Southwest. Nat. 39: 263-268. [ Links ]
Chavance, P., C. Flores-Coto & A. Sánchez-Iturbe. 1984. Early life history and adult biomass of sea bream in the Terminos Lagoon, southern Gulf of Mexico. Trans. Amer. Fish. Soc. 113: 166-177. [ Links ]
Dorsey, S.E., E.D. Houde & J.C. Gamble. 1996. Cohort abundances and daily variability in mortality of eggs and yolk-sac larvae of bay anchovy, Anchoa mitchilli, in Chesapeake Bay. Fish. Bull. 94: 257-267. [ Links ]
Flores-Coto C., F. Barba-Torres & J. Sánchez-Robles. 1983. Seasonal diversity, abundance, and distribution of ichthyoplankton in Tamiahua lagoon, western Gulf of Mexico. Trans. Amer. Fish. Soc. 442: 247-256. [ Links ]
Fulling, G.L. & M.S. Peterson. 1999. Estimation of small scale patchiness of zooplankton and an associated predator, Anchoa mitchilli. Gulf Res. Rep. 11: 72. [ Links ]
García, E. 1973. Modificaciones al Sistema de Clasificación climática de Köppen. Inst. Geografía, Univ. Nal. Autón, México D.F. 252 p. [ Links ]
Govoni, J.J. & J.E. Olney. 1991. Potential predation on fish eggs by the lobate ctenophore Mnemiopsis leidyi within and outside the Chesapeake Bay plume. Fish. Bull. 89: 181-186. [ Links ]
Griffith, S. & D.L. Bechler. 1995. The distribution and abundance of the bay anchovy, Anchoa mitchilli, in as Southeast Texas marsh lake system. Gulf Coast Res. Rep. 9: 117-122. [ Links ]
Gutiérrez, F. & F. Contreras. 1987. Variación estacional de los parámetros hidrológicos y nutrientes en la Laguna de Tamiahua, Veracruz, México, p. 23-37. In Gómez-
Aguirre, S. (ed.). Memoria VII Simposio Latinoamericano sobre Oceanografía Biológica. INP-SEPESCA-ALICMAR. México, D.F.
Mc Cullagh, P. & J.A. Nelder. 1989. Generalized linear models. Chapman . Londres. 511 p. [ Links ]
Méndez Vargas, M.L. 1980. Distribución y abundancia del ictioplancton de la Laguna de Alvarado, Veracruz, a lo largo de un ciclo anual, Tesis Licenciatura, Universidad Nacional Autónoma de México, México D.F., México. [ Links ]
Montgomery, C. 1984. Design and analysis of experiments. Wiley. Nueva York. 538 p. [ Links ]
Morton, T. & D. Moran. 1989. Species profiles: Life histories and environmental requirements of coastal fishes and invertebrates (Mid-Atlantic). Bay anchovy. Biol. Rep. U.S. Fish Wildl. Serv., 1989. 21 p. [ Links ]
Nelder, J.A & R.W.M. Wedderburn. 1972. Generalized linear models. Jour. Royal Stat. Soc. Ser. A, 135: 370- 384. [ Links ]
Ocaña-Luna, J.A., A.S. Luna-Calvo, F. Zavala-García & C. Flores-Coto. 1987. Distribución y abundancia de huevos de algunas especies de Engraulidos (Pisces) en la Laguna de Términos, Campeche, México. Biotica 12: 275-290. [ Links ]
Pebbles, E.B., J.R. Hall & S.G. Tolley. 1996. Egg production by the bay anchovy Anchoa mitchilli in relation to adult and larval prey fields. Mar. Ecol. Prog. Ser. 131: 61-73. [ Links ]
Purcell, J.E., D.A. Nemazie, S.E. Dorsey, E.D. Houde & J.C. Gamble. 1994. Predation mortality of bay anchovy Anchoa mitchilli eggs and larvae due to scyp-homedusae and ctenophores in Chesapeake Bay. Mar. Ecol. Progress Ser. 114: 47. [ Links ]
Rilling, G.C. & E.D. Houde. 1999. Regional and temporal variability in growth and mortality of bay anchovy, Anchoa mitchilli, larvae in Chesapeake Bay. Fish.Bull. 97: 555-569. [ Links ]
Tuckey, J.W. 1977. Exploratory data analysis. Addison-Wesley. Nueva York. 506 p. [ Links ]
Villalobos Figueroa, A., J.A. Cabrera, S. Gómez Aguirre, V. Arenas, F. Manrique, A. Reséndez Medina & G. de La Lanza. 1968. Informe Final de las Investigaciones realizadas en la Laguna de Tamiahua. Inst. Biología, Univ. Nal. Autón. México, México D.F. 76 p. [ Links ]
Vouglitois, J.J., K.W. Able, R.J. Kurtz & K.A. Tighe. 1987. Life history and population dynamics of the bay anchovy in New Jersey. Trans. Amer. Fish. Soc.116: 141-153. [ Links ]
1. Instituto de Investigaciones en Matemáticas Aplicadas y en Sistemas, UNAM, Apdo. Postal 20-726, Delegación Alvaro Obregón, C.P. 01000, México, D. F. Fax: 5622-3621; carlos@sigma.iimas.unam.mx
2. Juan Francisco Barba-Torres. Colegio de Ciencias y Humanidades y Facultad de Ciencias, UNAM. Cataratas y Llanura S/N, Col. Pedregal de San Ángel. México D. F.; fcobarba@servidor.unam.mx
3. Instituto Nacional de la Pesca, Pitágoras 1320 Col. Santa Cruz Atoyac, CP 03310, México, D.F.; tgaspar@inp.semarnap.gob.mx

