










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.51 n.2 San José Jun. 2003
How do mangrove forests induce sedimentation?
K. Kathiresan 1
Received 07-V-2002. Corrected 27-VI-2003. Accepted 30-VI-2003.
Abstract
The mangrove forests play a significant role as sediment traps. They reduce tidal flows and induce sedimentation of soil particles at low tide. However, there are no such processes taking place in the non-mangrove areas. Site of Avicennia–Rhizophora interphase is more efficient than Avicennia and or Rhizophora zones, in trapping the sediment by 30, 25 and 20% respectively at low tide as compared to high tide.
Key words: Rhizophora, Avicennia, sedimentation, mangrove.
Mangrove forests are one among the most productive ecosystems, lying between the land and sea in the tropical and sub-tropical latitudes (Kathiresan and Bingham 2001, Kathiresan 2002). One of the important functions of mangroves to environment is to provide a mechanism for trapping sediment, and thus the mangrove forests are believed to be an important sink of suspended sediment (Woodroffe 1992, Wolanski et al. 1992, Wolanski 1994, 1995, Furukawa et al. 1997). The mangrove trees catch sediment by their complex aerial root structure, thus functioning as land builders. In numerous cases, there has been proof of annual sedimentation rate in mangrove areas, ranging between 1 and 8 mm (Bird and Barson 1977). Woodroffe (1992) has a different view that the mangrove forests are a result, and not the cause of sedimentation in protected coastal areas, and that they only accelerate the sedimentation process.
The suspended sediment is introduced into coastal areas by river discharge, dumping of dredged material and resuspension of bottom sediment by waves and ships (Wolanski 1994). The mechanisms of sediment transport in mangrove waters are mostly based on the mechanism of hydrodynamic process rather than biological process (Ayukai and Wolanski 1997), and the hydrodynamic process includes the asymmetry of the tidal currents, the baroclinic circulation and shear-induced destruction of flocs (Gibbs 1985, Woodroffe 1985, Kjerfve 1990, Mazda et al. 1995, Wolanski and Gibbs 1995, Wolanski 1995, Wolanski et al. 1980, 1995). The mangroves trap the suspended sediments during their transport based on tidal flows (Wolanski et al. 1980). In India, there have been no previous studies, available on the tidal flows around mangrove vegetation and their role in the process of sedimentation, which forms a basis of this study.
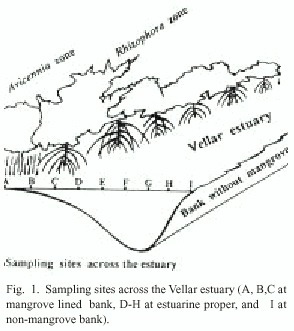
The study area is located along the Vellar estuary (11°29 N; 79°46 E), south east coast of India. One bank of the estuary has a stretch of 9-year old mangrove vegetation, artificially developed by the author, and another bank of the estuary is devoid of mangrove vegetation. Across the estuary, nine sampling points were fixed with a distance of 5 m between sampling points at the mangrove zones (A, B, C) and with a distance of 20 m between sampling points (D - H) at the estuarine proper. The sampling point - A refers to landward Avicennia zone, B to Avicennia-Rhizophora interphase, C to water-front Rhizophora zone, along the intertidal area. The sampling points from D to H are in the estuarine proper, and I denotes the opposite side of estuarine bank devoid of mangroves (Fig. 1).
Water column was measured for heights in cm using a scale. Velocity of water flows was measured in cm s -1 using a current meter (Emcon, Cochin). Suspended sediment concentration was measured in grams after filtering a known quantity of water through a Millipore filter unit and subsequent drying in an oven. The samples were made in triplicate from each sampling point, every 1 h for 12 h from 6 a.m. to 6 p.m. Average values for low and high tides are given in the figures.
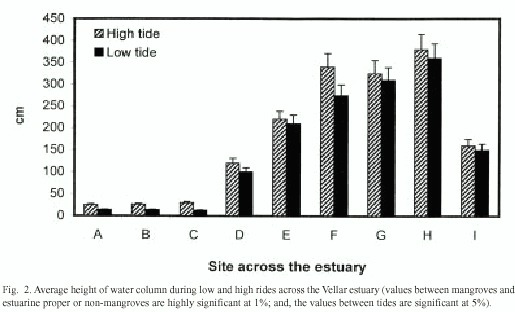
The present study analyzed water column height in different sampling sites across an estuarine system, at a mangrove-lined bank, estuarine proper, and a non-mangrove bank. The water column height ranged from 25 to 48 cm in the mangrove-lined bank, from 128 to 390 cm in the estuarine proper, and it varied from 128 to 150 cm. in the non-mangrove bank (Fig. 2). This indicated that the change in water column height due to tidal water was greater in the relatively deeper estuarine proper, than the shallower mangrove areas and the change in water level was little in the non-mangrove bank. Among the mangrove sites, the water level was low in the landward Avicennia zone and high in the low-lying Rhizophora zone of the intertidal slope.
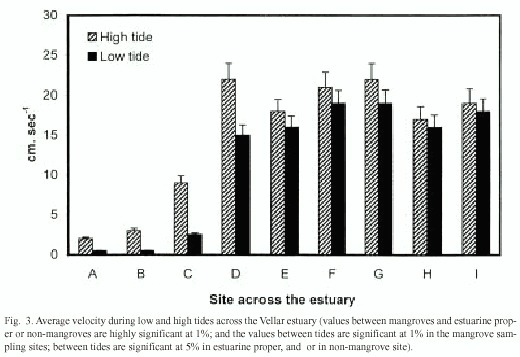
The velocity of water flow was found high in the estuarine proper, ranging from 16-23 cm/sec in the estuarine proper, and 18-20 cm/sec in the non-mangrove bank, whereas the velocity was low in the mangrove-lined bank, varying from 0-9 cm/sec (Fig. 3). The velocity was significantly greater in lowlying area than that in landward areas of the intertidal region. The difference in velocity between low and high tides was more significant in the mangrove areas than the others.
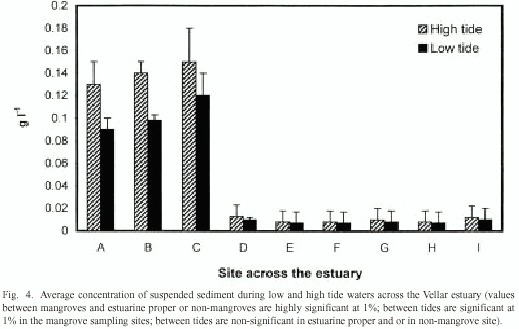
The concentration of particles suspended in water column ranged from 0.09 to 0.15 g/l in the mangrove-lined bank. This value was greater than that of other two areas varying from 0.008 to 0.01 g/l in both the estuarine proper and the non-mangrove area (Fig. 4). The change in suspended sediments between low and high tides was significantly greater in the mangrove-lined bank than that in other two areas.
The water-flows in mangrove forests are controlled by the tides. Water spills over from the estuary to inundate the forests at high tide. The water drains back to the estuary at low tide. The driving force for this flow is the slop-piness of the surface from the estuary into the forests (Furukawa et al. 1997). The height of water column is significantly lower in the mangrove sites than that in estuarine proper and in non-mangrove site, at both high and low tides (Fig. 2). A similar trend is seen with the velocity of water-flows (Fig. 3). The mangrove structures inhibit the tidal flow, probably due to friction force as suggested by previous workers (Woodroffe 1992, Wolanski et al. 1992, Wolanski 1994, 1995, Furukawa et al. 1997).
There is a significant difference in the concentration of suspended sediment between high and low tide waters in the mangrove zones. However, such situation does not occur in the non-mangrove areas and in the estuarine proper (Fig. 4). Sediment particles are carried in suspension into mangrove forests at high tide, and they are maintained in suspension (Fig. 4). This maintenance is due to the turbulence caused by mangrove structures as suggested by previous workers (Woodroffe 1992, Wolanski et al. 1992, Wolanski 1994, 1995, Furukawa et al. 1997). Concentration of suspended sediment is less at low tides in the mangrove forests (Fig. 4). This is due to the settlement of the suspended particles during low tide, as the velocity of water-flows that is small and sluggish (Fig. 3) to carry the sediment back to the estuary. However, there is only very low concentration of suspended sediment during both the tides in the estuarine proper and non-mangrove bank (Fig. 4). This is a clear evidence that mangroves trap the suspended sediment at the low tide, as also has been observed in Cairns mangroves, Australia (Furukawa et al. 1997).
The efficiency of sediment trapping varies with mangrove zones. Avicennia-Rhizophora interphase is much efficient by trapping at low tide 30% of total suspended sediment received at high tide. Avicennia zone is efficient by removing 25%, but Rhizophora can trap only 20% of sediment. The high efficiency of trapping suspended sediment in Avicennia-Rhizophora interphase may be attributed to wide spread occurrence of numerous pneumatophores (aerial respiratory roots) in Avicennia and to compactly arching, stilt roots of Rhizophora. The complex aerial roots of increased surface area of physical barrier might have caused turbulence facilitating sedimentation. The difference in trapping sediments between the two species may be due to the fact that Rhizophora experiences more of tidal inundations and relatively higher water-flow velocity as it grows in water-front areas, whereas Avicennia does less of tidal inundations and lower water velocity as it occurs widely towards the upland of intertidal slope. The density of the mangrove species and their complexity of root structures may also constitute important parameters, determining the sedimentation.
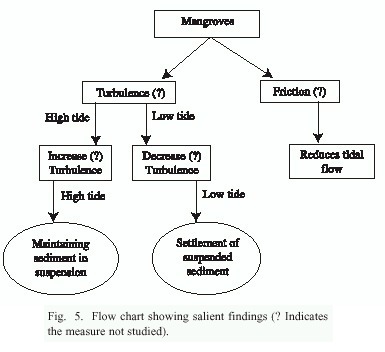
The salient findings of the study are depicted in fig. 5. The mangrove structures inhibit tidal flows, probably due to the friction force. The soil particles are carried in suspension into mangrove forest from estuarine sea-water at high tide, and the soil particles are maintained, probably due to turbulence caused by mangrove structures. The particles settle in the forests around low tide, probably when the turbulence reduces and also the water velocity at low tide is sluggish and small to carry the particles back to the estuary. Woodroffe (1992) believed that the mangroves are not the cause of sedimentation in protected coastal areas, and that they only accelerate the sedimentation process, which depends on the complexity of exchange process, which takes place between mangroves and adjoining coastal areas.
Acknowledgement
The author is thankful to the authorities of Annamalai University for providing facilities, and Dr. N. Rajendran for his help.
Resumen
Los bosques de manglar juegan un papel significativo como trampas de sedimentos. Ellos reducen los flujos de mareas e inducen las sedimentación de las partículas del suelo en la marea baja. Sin embargo, tal proceso no ocurre en areas sin manglares. Los sitios de interfase Avicennia–Rhizophora son más eficientes que las zonas monoespecíficas de Avicennia o de Rhizophora para atrapar sedimentos, en un 30, 25 and 20% respectivament, durante la marea baja en comparación con la marea alta.
References
Ayukai, T. & E. Wolanski. 1997. Importance of biologically medicated removal of the sediments from the Fly River plume. Papua New Guinea. Estuar. Coast. Shelf Sci. 44: 629-639. [ Links ]
Bird, E.C.F. & M.M. Barson. 1977. Measurement of physiographic changes on mangrove-fringed estuaries coastlines. Mar. Res. Indonesia. 18: 73-80. [ Links ]
Furukawa, K., E. Wolanski & H. Mueller. 1997. Currents and sediment transport in mangrove forests. Estuar. Coast. Shelf Sci. 44: 301-310. [ Links ]
Gibbs, R. J. 1985. Estuarine flocs: their size, settling velocity and density. J. Geophysical Res. 90: 3249-3251. [ Links ]
Kathiresan, K. 2002. Greening the blue mud! Rev. Biol. Trop. 50: 869-874. [ Links ]
Kathiresan, K. & B.L. Bingham. 2001. Biology of mangroves and mangrove ecosystems. Advance. Mar. Biol. 40: 81-251. [ Links ]
Kjerfve, B. 1990. Manual for investigation of hydrological processes in mangrove ecosystems. UNESCO/ UNDP Regional Project. l990, 79 p. [ Links ]
Mazda, Y., N. Kanazawa & E. Wolanski. 1995. Tidal asymmetry in mangrove creeks. Hydrobiologia 295: 51-58. [ Links ]
Wolanski, E. 1994. In Physical oceanography processes of the Great Barrier Reef. CRC, Boca Raton, Florida. [ Links ]
Wolanski, E. 1995. Transport of sediment in mangrove swamps. Hydrobiologia 295: 31-42. [ Links ]
Wolanski, E. & R.J. Gibbs. 1995. Flocculation of suspended sediment in the Fly River estuary, Papua New Guinea. J. Coast. Res. 40: 321-337. [ Links ]
Wolanski, E. M., M. Jones & J.S. Bunt. 1980. Hydrodynamics of a tidal creek - mangrove swamp ecosystem. Aust. J. Mar. Freshwat. Res. 31: 431-450. [ Links ]
Wolanski, E., Y. Mazda & P. Ridd. 1992. Mangrove hydrodynamics. pp. 436-462. In A.I. Robertson & D.M. Alongi (eds.). Tropical mangrove ecosystem, American Geophysical Union, Washington D.C. [ Links ]
Wolanski, E., B. King & D. Galloway. 1995. Dynamics of the turbidity maximum in the Fly River estuary, Papua New Guinea. Estuar. Coast. Shelf Sci. 40: 321-337. [ Links ]
Woodroffe, C.D. 1985. Studies of a mangrove basin, Tuff Crater, New Zealand: II. Comparison of volumetric and velocity - area methods of estimating tidal flux. Estuar. Coast. Shelf Sci. 20: 431-445. [ Links ]
Woodroffe, C. 1992. Mangrove sediments and geomorphology. pp. 7-41. In A. I. Robertson & D.M. Alongi (eds.). Tropical mangrove ecosystem, American Geophysical Union, Washington D.C. [ Links ]
1 Centre of Advanced Study in Marine Biology, (Annamalai University), Parangipettai 608 502. Tamil Nadu, India. Phone: +91 4144 243533 ext. 210 (Office) +91 4144 238419 (Home). Fax: +91 4144 238419 / 243555; cdl_aucasmb@sancharnet.in ; cdl_mangrove@sancharnet.in

