










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.51 n.1 San José Mar. 2003
Abstract
The effects of sex, injuries, season and site on the growth of the spiny lobsters Panulirus gracilis, and P. inflatus, were studied through mark-recapture techniques in two sites with different ecological characteristics on the coast of Guerrero, México. Panulirus gracilis occurred in both sites, whereas P. inflatus occurred only in one site. All recaptured individuals were adults. Both species had similar intermolt periods, but P. gracilis had significantly higher growth rates (mm carapace length week -1 ) than P. inflatus as a result of a larger molt increment. Growth rates of males were higher than those of females in both species owing to larger molt increments and shorter intermolt periods in males. Injuries had no effect on growth rates in either species. Individuals of P. gracilis grew faster in site 1 than in site 2. Therefore, the effect of season on growth of P. gracilis was analyzed separately in each site. In site 2, growth rates of P. gracilis were similar in summer and in winter, whereas insite 1 both species had higher growth rates in winter than in summer. This could be due to spatial differences in processes related to changes in population density and food resources, which were documented in previous works. The overall results show that P. gracilis grows faster than P. inflatus, and that growth rates of both species are highly variable and are affected by environmental factors such as site and season, which should be taken into account when attempting to produce population growth curves for each species.
Key words: Growth rates, growth variability, intermolt period, molt increments, Panulirus gracilis, Panulirus inflatus, spiny lobsters.
Growth in crustaceans occurs by a series of molts or ecdyses. The rate of growth is determined by two factors: the molt increment, i.e. the increase in size at each molt, and the intermolt period, i.e. the interval between successive molts. These factors do not remain constant with growth, but generally as size increases the percentage molt increment decreases whereas the intermolt period lengthens (Hartnoll 1985). Information on both growth factors is necessary to provide a complete description of growth for populations of spiny lobsters (Morgan 1980).
The spiny lobster Panulirus inflatus (Bouvier, 1895) is endemic to the Pacific coast of Mexico, from Baja California Sur to Oaxaca, whereas Panulirus gracilis Streets, 1871, occurs from Baja California Sur to the west coast of Peru and the Galapagos Islands in South America (Holthuis 1991). Hence, both species co-occur throughout most of the Pacific coast of Mexico. However, each species has different habitat requirements: P. inflatus occurs solely in areas with rocky bot-toms and relatively clear waters, whereas P. gracilis inhabits both rocky and gravel-sand bottoms, and tolerates a wider range of water turbidity (Briones et al. 1981, Lozano et al. 1982, Pérez González et al. 1992).
Both species are fished throughout their geographic range, but in relatively low numbers. In Mexico, the combined catch of both species comprised 10–20% of the total lobster catch of the Pacific coast between 1985 and 1997 (Briones-Fourzán and Lozano-Álvarez 2000). Because of their limited fishery importance, both species have remained poorly studied. In the late 1970s and early 1980s, our research group conducted a tagging program to study the population dynamics of P. inflatus and P. gracilis in Guerrero, Mexico. Results on population density (Lozano et al. 1982), feeding ecology (Aramoni-Serrano 1982, Lozano-Álvarez and Aramoni-Serrano 1996), and reproductive dynamics (Briones-Fourzán and Lozano-Álvarez 1992) were published, but the data on growth remained unprocessed. To our knowledge, no estimates on growth of any of these two species have been produced to date.
Because of the scarcity of scientific literature on both P. inflatus and P. gracilis, we present the growth results of both species in this paper. We hypothesized that: (a) both species would have similar growth rates because they reach similar sizes; (b) males would grow faster than females in both species; (c) small individuals would grow faster than large individuals; (d) injured individuals (those missing appendages) would have lower growth rates than uninjured (whole) individuals because of the energy required to regenerate appendages; and (e) both species would have either similar growth rates in summer or winter, or alternatively, higher growth rates in summer than in winter.
Materials and methods
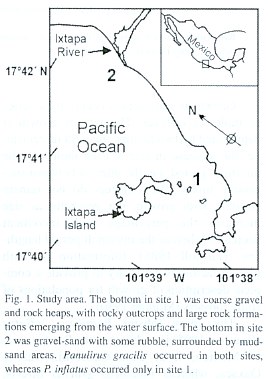
The study was conducted in two sites with different ecological characteristics along the coast of the state of Guerrero, on the Pacific coast of Mexico (Fig. 1). Site 1 was located between the continental coast and Ixtapa Island (17º42.5 N, 101º38.8 W; ~1.2 km to the west of the coast), and had a maximum depth of 10 m. The bottom in site 1 consisted of rubble (coarse gravel with rock heaps), rocky outcrops and large rock formations emerging from the water surface. Both P. inflatus and P. gracilis occurred in site 1, with a predominance of P. inflatu s. Site 2 was located 2.5 km north of site 1, in front of the mouth of the Ixtapa River (17º 42.0 N, 101º 37.0º W). The mouth of the river remained closed during the dry season (November-May) and opened during the rainy season (June-October). The bottom in site 2 was mostly gravel-sand with some rubble, surrounded by sand-mud areas, and had a maximum depth of 8 m. Water was murkier in site 2 than in site 1 because of the river discharge. Hence, only P. gracilis occurred in site 2. Sea surface temperatures in the study area range from an average minimum of 24ºC in April, to a maximum of 30.5ºC in August (Baqueiro and Stuardo 1977, Briones et al. 1981, Aramoni-Serrano 1982).
Lobsters were caught in both sites from February 1979 to May 1980 with bottom-set nets measuring 80–150 m long and 2 m in height. Mesh size ranged from 11.5 to 15.0 cm. The nets were deployed on gravel-sand or sand-mud bottoms. In site 1, the nets were set close to the rock heaps and formations, but not directly on these, and 20 wire-mesh lobster traps (mesh size: 5 x 2.5 cm) were also used. The traps were 1.5 m long, 0.8 m wide, and 0.6 m high, and were baited with assorted fish heads. Both sites were sampled at least three times per week during the whole study period, unless precluded by bad weather. This allowed for multiple recaptures of many individuals. Lobsters were tagged with individually numbered spaghetti-type tags (Floy Tag FD–68B), inserted into the dorsolateral extensor muscle between the cephalothorax and the abdomen. Once tagged, the lobsters were immediately released in the same place of capture.
Data collected for each lobster were species, tag number, capture date, capture site, sex, injuries (e.g. number and type of missing appendages), carapace length (CL, measured from between the rostral horns to the posterior edge of the carapace, ±0.1 mm), and carapace fouling stage (from 1: completely clean, to 4: heavily fouled).
Growth data were analyzed as mean weekly growth rate (increase in mm CL week -1), intermolt period (the time elapsed between two molts, in weeks), and molt increment (estimated by substracting initial CL from CL at recapture). These three growth variables were estimated for each species and for the following factors: sex, injury (injured or uninjured individuals), and season ("summer" and "winter"). "Summer" encompassed the warmest months, from June to December (sea surface temperature range: 27.5–30.5ºC), whereas "winter" included the coolest months, from January to May (24.0–26.5ºC). In P. gracilis, an additional factor was site (site 1 or site 2).
Mean weekly growth rate reflects the combined effects of molt increment and intermolt period, allowing comparisons over a wide range of sizes and conditions. Intermolt period was calculated by means of a technique pro-posed by Munro (1974) and further utilized by Davis (1981) and Hunt and Lyons (1986), which appeared reasonable because molting activity in both species occurred throughout the year. The technique assumes that at the time of tagging the lobsters were randomly distributed throughout their molting cycle, and that 50% had therefore molted when half of their intermolt period had elapsed. Therefore, the intermolt period is estimated as twice the time interval (in weeks) over which half of all lobsters for a given factor molted. Based on this rationale, molt increment was calculated only from lobsters that remained at large <50% of the intermolt period obtained for each factor (sex, injury, season, and site). Because of possible measurement errors, only changes >2 mm CL were considered as growth (Hunt and Lyons 1986), unless individuals regenerated appendages or changed from carapace fouling stages 3–4 to 1–2 between subsequent captures.
Differences between means were evaluated using Students t-tests for unequal sample size (Zar 1984) for each factor combination. When necessary, data were log-transformed to homogenize variances between groups.
Results
In total, 1 768 lobsters were tagged, of which 1 307 were P. gracilis and 461 P. inflatus. Sex ratio was heavily skewed toward males in both species (3.6:1 in P. gracilis; 4.2:1 in P. inflatus). Nets and traps catch more males than females because breeding females are more reclusive than non-breeding females or males, and females of both species can produce up to four or five broods per year (Lozano et al. 1982; Briones-Fourzán and Lozano-Álvarez 1992). Hence, most recaptured individuals were also males. In total, we recaptured 220 males (CL range: 48.0–107.5 mm CL) and 21 females (56.3–88.9 mm CL) of P. gracili s, and 34 males (58.1–111.7 mm CL) and 10 females (68.0–82.5 mm CL) of P. inflatus. Both species attain sexual maturity between 45 and 50 mm CL (Briones et al. 1981, Briones-Fourzán and Lozano-Álvarez 1992), so virtually all the recaptured lobsters were adults. Individuals of P. gracilis remained at large from 12 to 253 days (median: 53 days), and individuals of P. inflatus from 22 to 185 days (median: 64 days).
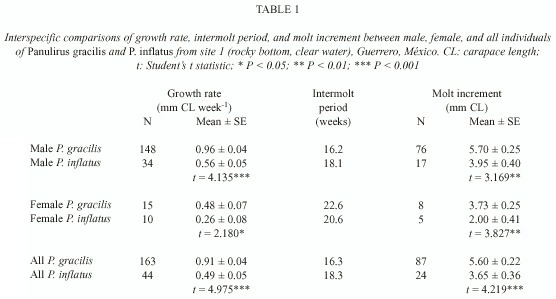
A comparison of growth between the two species revealed that males and females of P. gracilis grew faster than males and females of P. inflatus (Table 1). This significant difference was not related to differences in intermolt periods, which were similar for individuals of the same sex of both species, but to a larger molt increment in individuals of P. gracilis. When combining data of both sexes for each species, individuals of P. gracilis had a mean growth rate of 0.91 mm CL week -1 , and in P. inflatus the overall average growth rate was 0.49 mm CL week -1 . The intermolt period was similar in both species, but P. gracilis had a significantly larger molt increment than P. inflatus (Table 1).
The relatively small number of recaptures of P. inflatus precluded an analysis of growth rate by size class for this species. This was true also for females of P. gracilis. In P. gracilis males, data allowed for an analysis of weekly growth rate (increase in CL week -1 ) by 5-mm size class on a seasonal basis (Fig. 2). Males in size classes below 75 mm CL had higher growth rates than males above 75 mm CL, but in general, the weekly growth rate by size class was higher and more variable in winter than in summer. The wider variability of growth rates during winter was due to the additional effects of site, as further analyzed below.

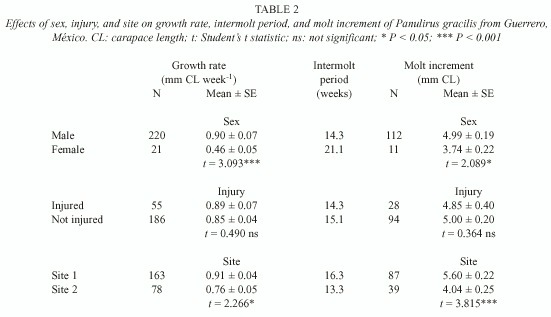
Overall, males had significantly higher weekly growth rates than females in P. gracilis, as a result of both a shorter intermolt period and a larger molt increment in males (Table 2). Injuries had no effect on growth rate, intermolt period or molt increment (Table 2). Individual P. gracilis had significantly higher growth rates in site 1 than in site 2. This difference was not due to the intermolt period, which was slightly longer in site 1, but to a larger molt increment in lobsters from site 1 (Table 2). The differences in growth between individual P. gracilis from site 1 and site 2 warranted a separate analysis of season effects on each site. In site 2, although intermolt period was slightly longer in winter, growth rate and molt increment were similar in both seasons. In contrast, growth rate and molt increment were significantly higher in winter in site 1, whereas intermolt period was similar in both seasons (Table 3).
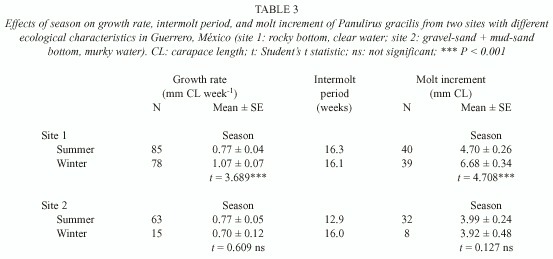
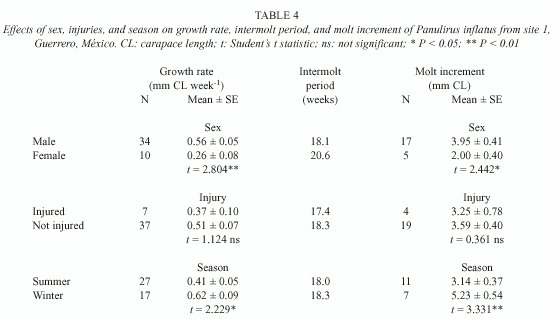
The effects of sex, injury and season on growth of P. inflatus were similar to those on P. gracilis. Female P. inflatus had significantly lower growth rates than males, as a combined effect of a longer intermolt period and a smaller molt increment in females (Table 4). Injuries had no effect on growth in P. inflatus, but individuals had significantly higher growth rates and molt increments in winter than in summer (Table 4).
Discussion
Our results show that P. gracilis grows faster than P. inflatus. Weinborn (1977) reached a similar conclusion through CL-frequencies analysis. This result was intriguing given the many similarities between the two species. Both reach similar CL sizes (largest P. inflatus recorded in Guerrero: 130 mm CL; largest P. gracili s: 124 mm CL, Briones et al. 1981), and females of both species attain sexual maturity at a similar CL: 47.5 mm in P. gracilis (Weinborn 1977), and 45.6 mm CL in P. inflatus (Gracia 1985), with 50% of ovigerous females below 74 mm CL in P. gracilis, and 70 mm CL in P. inflatus (Briones et al. 1981). However, the two species also have a number of differences that may be related, at least par-tially, to their differential growth rates. Although both species have a similar diet in terms of gross food items (Lozano-Álvarez and Aramoni-Serrano 1996), the greater flexibility in habitat use of P. gracilis may allow this species to successfully exploit more resources than P. inflatus. Also, P. inflatus has a disproportionately large carapace, such that at a same total length, this species has a larger CL than P. gracilis (Chapa 1964), and at a same CL, females of P. gracilis have a larger brood size than females of P. inflatus (Fernández-Lomelín 1992).
Despite its variability, the growth rate of P. gracilis was lower in the largest size classes, which is a common pattern in decapods (Hartnoll 1985). In both P. gracilis and P. inflatus, males grew more than females. This is a general characteristic of spiny lobsters (Morgan 1980). Females grow smaller than males because ovarian maturation and breeding suppress growth and prolong the intermolt period, especially in repetitive-breeding species that do not necessarily molt between broods (Quackenbush 1994), such as P. gracilis and P. inflatus, which can produce up to four or five broods in a year (Briones-Fourzán and Lozano-Álvarez 1992).
Injuries affect growth of different species of lobsters in different ways. In some cases, injured lobsters had shorter intermolt periods but smaller growth rates than uninjured lobsters (Chittleborough 1975). In other cases, injuries affected growth rates of immature but not of mature individuals (Davis 1981, Hunt and Lyons 1986), or had no effect at all (Forcucci et al. 1994). Injuries had no effect on growth either in P. gracilis or in P. inflatus. However, most of our injured individuals were missing only one leg or one antenna. The lack of more appendages might have a greater effect on growth rates in these two species, as was found in P. cygnus (Brown and Caputi 1985).
In general, growth rates of spiny lobsters are related to temperature and photoperiod. In temperate or relatively cold areas, spiny lobsters molt once a year (Pollock 1986). In contrast, most tropical species do not have a specific molting season, although their growth rates still show some seasonality. For example, in Jamaica and Florida (USA), P. argus grows faster in summer than in winter (Munro 1974, Davis 1981, Hunt and Lyons 1986, Forcucci et al. 1994) due to longer intermolt periods during the winter. Given their tropical nature, we expected the growth rates of P. gracilis and P. inflatus to be either similar in winter and summer, or higher in summer. In site 2, P. gracilis had no seasonal differences in growth, but in site 1, counter to our expectations, both species grew significantly faster in winter. We believe that this seeming contradictory result might have been caused by processes related to variations in lobster density and food resources in site 1, as explained below.
Growth rates of several species of palin-urids vary widely among different areas. In some cases, these variations are related to temperature, but in other cases they have been ascribed to differences either in food availability or in the densities of lobsters in relation to food supplies (Newman and Pollock 1974, Barkai and Branch 1988, Edgar 1990, Melville-Smith and Goosen 1995). Although spiny lobsters are communal in nature, at very high densities their growth can be depressed (Chittleborough 1976, Pollock 1986, 1991, Barkai and Branch 1988, Jernakoff et al. 1994, Lozano-Álvarez 1996; Booth and Kittaka 2000), suggesting a density-dependent regulation of growth rate.
Aramoni-Serrano (1982), and Lozano-Álvarez and Aramoni-Serrano (1996) studied the feeding ecology of P. gracilis and P. inflatus in the same area and at the same time as our study. They found that during September and October (i.e. within our "summer" season), there was a significant increase in organic matter and biomass of mollusks (the main food item in both lobster species) in site 1 compared to other adjacent sites. Moreover, Lozano et al. (1982) reported that in September-October there was an important immigration of individuals of both species to site 1, apparently as a result of the higher food availability in that site, which rapidly reached a crowding situation because lobster density increased up to four times its average values during the rest of the year. This sudden increase in density may have offset the effects of the large food supply, resulting in a slower growth rate of both species during the summer. Further on, in November-December, the density of both species in site 1 decreased to its average values (Lozano et al. 1982). In early January, individuals of P. inflatus caught in site 1 had a low condition factor and their hepatopancreas were in a poor condition, but by May they were fully recovered (Lozano-Álvarez and Aramoni-Serrano 1996). Therefore, the return to average levels of population density may have allowed individual lobsters to use the existing food resources more efficiently, increasing their growth rates during our "winter" season (i.e. January-May). This hypothesis conforms to Breens (1994) observation that different species of lobsters might be regulated in different ways, and that the same population might be regulated by different mechanisms as abundance varies.
In conclusion, our results show that P. gracilis grows faster than P. inflatus, males grow faster than females; large individuals have lower growth rates than small individuals, but growth of both species is highly variable and is affected by environmental factors such as site and season. However, many issues remain inconclusive and warrant more studies. Additional data on growth of females are clearly needed. Other biotic and abiotic factors not addressed in our study, such as salinity and primary production, may have also accounted for the different seasonal growth rates of lobsters in site 1. Interannual variations in growth rate should also be explored, as well as the implications for the fishery of the different and variable growth rates of these two species.
General growth models commonly used in fisheries assessments, such as von Bertalanffys growth function and Mauchlines linear relationships (Mauchline 1977), assume that the growth rate is constant throughout any one year, which is clearly not the case of P. gracilis and P. inflatus. Their variability in growth should be taken into account when attempting to produce population growth curves for each species.
Acknowledgments
We thank Adolfo Gracia, Leonardo Santarelli, Carlos Illescas and Gabriela Aramoni for their help in field and laboratory activities. This project was funded by Instituto Nacional de la Pesca, México, and Universidad Nacional Autónoma de México.
Resumen
Se analizaron, por medio de marcado-recaptura, los efectos del sexo, heridas, estación del año y localidad sobre el crecimiento de las langostas espinosas Panulirus gracilis Streets, 1871, y Panulirus inflatus (Bouvier, 1895) en dos localidades con diferentes características ecológicas en la costa de Guerrero, México. Panulirus gracilis se presentó en ambas localidades, mientras que P. inflatus sólo se encontró en una de ellas. Todos los individuos recapturados eran adultos. Ambas especies presentaron períodos de intermuda similares, pero P. gracilis tuvo tasas de crecimiento (mm de longitud cefalotorácica semana -1 ) mayores que las de P. inflatus, debido a un mayor incremento por muda. Los machos crecieron más rápidamente que las hembras en ambas especies, ya que presentaron incrementos por muda mayores y períodos de intermuda más cortos.Las heridas no afectaron las tasas de crecimiento en ninguna de las dos especies. Los individuos de P. gracilis crecieron más rápidamente en la localidad 1 que en la localidad 2. Por tanto, se analizaron los efectos estacionales sobre el crecimiento de P. gracilis en cada localidad por separado. En la localidad 2, las tasas de crecimiento de P. gracilis fueron similares en verano e invierno, mientras que en la localidad 1 la tasa de crecimiento de ambas especies fue mayor en invierno que en verano. Esto podría ser resultado de diferencias espaciales en procesos relacionados con cambios en la densidad poblacional y en los recursos alimentarios, documentados en trabajos anteriores. Los resultados globales muestran que P. gracilis crece más rápidamente que P. inflatus, y que ambas especies tienen tasas de crecimiento altamente variables, las cuales son afectadas por factores ambientales tales como localidad y estación del año, lo que debe ser tomado en consideración cuando se pretenda producir curvas de recimiento poblacional para cada una de estas especies.
References
Aramoni-Serrano, G. 1982. Alimentación de las langostas Panulirus inflatus (Bouvier) y P. gracilis Streets en Zihuatanejo, Guerrero y su relación con el bentos. Tesis Profesional, Universidad Nacional Autónoma de México, México. 66 p. [ Links ]
Baqueiro, E. & J. Stuardo. 1977. Observaciones sobre la biología, ecología y explotación de Megapitaria aurantiaca (Sow., 1931), M. squalida (Sow., 1835) y Dosinia ponderosa (Gray, 1838) (Bivalvia: Veneridae) de la Bahía de Zihuatanejo e Isla Ixtapa, Gro., México. An. Centro Cienc. Mar Limnol. Univ. Nal. Autón. México 4: 161–208.
Barkai, A. & G. M. Branch. 1988. Energy requirements for a dense population of rock lobsters Jasus lalandii : novel importance of unorthodox food resources. Mar. Ecol. Prog. Ser. 50: 83–96. [ Links ]
Booth, J.D. & J. Kittaka. 2000. Spiny lobster growout, p. 556–585. In B.F. Phillips & J. Kittaka (eds.) Spiny lobsters: Fisheries and culture. Fishing News Books, Oxford. [ Links ]
Breen, P. 1994. Population dynamics and stock assessment of lobsters: a review. Crustaceana 67: 239–255. [ Links ]
Briones-Fourzán, P. & E. Lozano-Álvarez. 1992. Aspects of the reproduction of Panulirus inflatus (Bouvier) and P. gracilis Streets (Decapoda: Palinuridae) from the Pacific coast of Mexico, J. Crustacean Biol. 12: 41–50.
Briones-Fourzán, P. & E. Lozano-Álvarez. 2000. The spiny lobster fisheries in Mexico, p. 169–188. In B.F. Phillips & J. Kittaka (eds.) Spiny lobsters: Fisheries and culture. Fishing News Books, Oxford.
Briones, P., E. Lozano, A. Martínez & S. Cortés. 1981. Aspectos generales de la biología y pesca de las langostas en Zihuatanejo, Gro., México (Crustacea: Palinuridae). An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México 8: 79–102.
Brown, R. S. & N. Caputi. 1985. Factors affecting the growth of undersize western rock lobster, Panulirus cygnus George, returned by fishermen to the sea. Fish. Bull. 83: 567–574. [ Links ]
Chapa, H. 1964. Contribución al conocimiento de las langostas del Pacífico mexicano y sus pesquerías. Inst. Nal. Inv. Biol. Pesq, Sec. Ind. Com. México, Publ. 6:1–68. [ Links ]
Chittleborough, R.G. 1975. Environmental factors affecting growth and survival of juvenile Western rock lobsters Panulirus longipes (Milne-Edwards). Aust. J. Mar. Freshwater Res. 26: 177–196. [ Links ]
Chittleborough, R. G. 1976. Growth of juvenile western rock lobsters Panulirus longipes cygnus George on coastal reefs compared with those reared under optimal environmental conditions. Aust. J. Mar. Freshwater Res. 27: 279–295.
Davis, G. E. 1981. Effect of injuries on spiny lobster,Panulirus argus, and implications for fishery man-agement. Fish. Bull. 78: 979–984. [ Links ]
Edgar, G. J. 1990. Predator-prey interaction in seagrass beds. I. The influence of macrofaunal abundance and size-structure on the diet and growth of the western rock lobster Panulirus cygnus George. J. Exp. Mar. Biol. Ecol. 139: 1–22. [ Links ]
Fernández-Lomelín, M. P. 1992. Potencial reproductivo de las langostas Panulirus gracilis Streets, 1871, y Panulirus inflatus (Bouvier, 1895). Tesis Profesional, Universidad Nacional Autónoma de México, México. 52 p.
Forcucci, D., M. J. Butler IV & J. H. Hunt. 1994. Population dynamics of juvenile Caribbean spiny lobster, Panulirus argus, in Florida Bay, Florida. Bull. Mar. Sci. 54: 805–818.
Gracia, A. 1985. Variación estacional en la fecundidad de la langosta Panulirus inflatus (Bouvier, 1895) (Crustacea: Decapoda: Palinuridae). Ciencias Marinas 11: 7-27.
Hartnoll, R.G. 1985. Growth, sexual maturity and reproductive output, pp. 101–128. In A.M. Wenner (ed.) Factors in adult growth. Crustacean Issues 3. Balkema, Rotterdam.
Holthuis, L.B. 1991. Marine lobsters of the world: An annotated an illustrated catalogue of species of interest to fisheries known to date. FAO Fisheries Synopsis 125, Vol. 13. Rome. 292 p. [ Links ]
Hunt, J.H. & W.G. Lyons. 1986. Factors affecting growth and maturation of spiny lobsters, Panulirus argus, in the Florida Keys. Can. J. Fish. Aquat. Sci. 43:2243–2247. [ Links ]
Jernakoff, P., J. Fitzpatrick, B.F. Phillips & E. de Boer. 1994. Density and growth in a population of juvenile western rock lobsters, Panulirus cygnus (George). Aust. J. Mar. Freshwater Res. 45: 69–81.
Lozano-Álvarez, E. 1996. Ongrowing of juvenile spiny lobster, Panulirus argus (Latreille, 1804) (Decapoda, Palinuridae), in portable sea enclosures. Crustaceana 69: 958–973. [ Links ]
Lozano-Álvarez, E. & G. Aramoni-Serrano. 1996. Alimentación y estado nutricional de las langostas Panulirus inflatus y Panulirus gracilis (Decapoda: Palinuridae) en Guerrero, México. Rev. Biol. Trop. 44: 453–461.
Lozano, E., P. Briones, L. Santarelli & A. Gracia. 1982. Densidad poblacional de Panulirus gracilis Streets y P. inflatus (Bouvier) (Crustacea: Palinuridae) en dos áreas cercanas a Zihuatanejo, Gro., México. Ciencia Pesquera 3: 61–73.
Mauchline, J. 1977. Growth of shrimps, crabs, and lobsters: an assessment. J. Cons. Int. Explor. Mer 37:162–169.
Melville-Smith, R. & P. C. Goosen. 1995. The spiny lobster Jasus lalandii (H. Milne.Edwards, 1837) off the South African coast: inter-annual variability in male growth and female fecundity. Crustaceana 68:174–183.
Morgan, G.R. 1980. Population dynamics of spiny lobsters, p. 189–217. In J.S. Cobb & B.F. Phillips (eds.) The biology and management of lobsters, Vol 2. Academic, New York.
Munro, J.L. 1974. The biology, ecology, exploitation, and management of Caribbean reef fishes. Part V. 1. The biology, ecology and bionomics of Caribbean reef fishes: Crustaceans (spiny lobsters and crabs). Univ. West Indies Zool. Dep. Res. Rep. 3: 1–57.
Newman, G. G. & D. E. Pollock. 1974. Growth of the rock lobster Jasus lalandii and its relationship to benthos. Mar. Biol. 24: 339–346. [ Links ]
Pérez-González, R., L.M. Flores-Campaña & A. Núñez-Pastén. 1992. Análisis de la distribución de tallas, captura y esfuerzo en la pesquería de langostas Panulirus inflatus (Bouvier, 1895) y P. gracilis Streets, 1871 (Decapoda: Palinuridae) en las costas de Sinaloa, México. Proc. San Diego Soc. Nat. Hist. 15: 1–5.
Pollock, D.E. 1986. Review of the fishery for and biology of the Cape rock lobster Jasus lalandii with notes on larval recruitment. Can. J. Fish. Aquat. Sci. 43: 2107–2117. [ Links ]
Pollock, D.E. 1991. Spiny lobsters at Tristan da Cunha, South Atlantic: inter-island variations in growth and population structure. S. Afr. J. Mar. Sci. 10: 1–12. [ Links ]
Quackenbush, L.S. 1994. Lobster reproduction: a review. Crustaceana 67: 82–94. [ Links ]
Weinborn, J.A. 1977. Estudio prelimiar de la biología, ecología y semicultivo de los Palinúridos de Zihuatanejo, Gro., México, Panulirus gracilis Streets y Panulirus inflatus (Bouvier). An. Centro Cienc. Mar Limnol. Univ. Nal. Autón. México 4: 27-78.
Zar, J.H. 1984. Biostatistical analysis. Prentice-Hall, New Jersey. 718 p. [ Links ]
1 Universidad Nacional Autónoma de México, Instituto de Ciencias del Mar y Limnología, Unidad Académica Puerto Morelos. P. O. Box 1152, Cancún, Q. R. 77500 México. Fax: +52 (998) 871-0138; briones@mar.icmyl.unam.mx

