










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.3-4 San José Dec. 2002
Abstract
Population sizes of six mammal species were estimated using the King method during the late dry season (March) of 1996 in the Punta Leona Private Wildlife Refuge, Costa Rica. The white-faced monkey (Cebus capucinus), coati (Nasua narica) and nine-banded armadillo (Dasypus novemcinctus) with 148, 46, and 8 sighted individuals, respectively, demonstrated the largest populations in the refuge. The Central American spider monkey (Ateles geoffroyi), tamandua anteater (Tamandua mexicana), and variegated squirrel (Sciurus variegatoides) were also included in the census. Population densities are calculated, habitat types are describes, and habitat use and activity periods are discussed.
Key words
Costa Rica, dry season, King method, mammals, primates.
Accurate estimates of wildlife population densities are an important initial step in the creation of effective conservation and management plans. Accordingly, scientists, conservationists, and policy makers seek reliable, accurate and feasible population estimation methods (Chapman et al. 1988 ). The large diversity of animal species in Costa Rica, coupled with the general lack of information regarding animal population dynamics, requires development of sampling techniques to quantify species numbers and arrive at the most accurate density estimates possible. For arboreal primates and highly mobile mammals, repeatedly sampling strip transects is the most common method of estimating population density (Cant 1978 , Freeze et al. 1982 , Wilson and Johns 1982 , Defler and Pintor 1985 ). However, transect methodology is highly variable regarding number and placement of transects and degree of habitat stratification and species behavior. Information must be collected on the appropriateness of different methods for particular situations. The National System of Conservation Areas (SINAC) of Costa Rica and adjacent private reserves protect over 27% of Costa Rica's 51,000 km 2 and offer an ideal protected environment for the realization of population size studies (Vaughan and Rodriguez 1997 , Vaughan and McCoy 1984 ). The present work describes the censusing of six mammal species inhabiting a Costa Rican private wildlife refuge, with calculation of population densities and discussion of habitat use and activity periods. This is the first manunalian census within this refuge and is thus important for the development of a more effective wildlife management strategy in Punta Leona.
Materials and methods
Study site: The study was conducted at the Punta Leona Private Wildlife Refuge located on the southern shore of the mouth of the Gulf of Nicoya near Jacó, Puntarenas Province, Costa Rica. The refuge was created nearly 25 years ago and is privately owned by the Punta Leona Hotel and Club. lt consists of approximately 300 ha and although designated as predominately tropical-wet forest (Janzen 1983 ), the area includes patches of mixed primary and secondary, wet and dry forest adjacent to the entrance road and surrounding the main lodging area. Geographically, the refuge is located on the Central Pacific coast and is characterized by a small central valley (containing the hotel and associated buildings) and severas sandy beaches flanked by rocky escarpments. Surrounding forested ridges are inhabited by appreciable bird and mammal populations and encompass a mosaic of distinct habitat types resulting from varying local environmental conditions and land manipulation in the past. The region experiences a defined dry season (December-May), and wet season (June-November).
Transect lines: Census of animal populations in heterogeneous habitats leads to difficulties in achieving sufficient sampling of all habitats. Particularly in tropical regions, the variation in habitat use exhibited by specialized or selective animals, and problems of visibility and accessibility resulting from dense vegetation growth, often lead to only the partial sampling of habitats (Chapman et al. 1988 ). To include representative habitats in this study, the entire refuge was initially explored and habitat types were defined and describes prior to the establishment of transects. Habitat classification was based on vegetation and largely defined by the height of the canopy and percentage of leaves retained by the trees during the latter part of the dry season. Six habitat types were describes in the study area (Table 1 ): 1) primary-wet, defined by evergreen vegetation and a high canopy, with a relatively thin understory, little fallen leaf matter on the forest floor, and large diameter primary growth trees present; 2) secondary-wet, similar to primary-wet with evergreen foliage, but marked by a significantly lower canopy, developed understory growth, and the presence of palms (Scheelea rostrata) and small diameter trees; 3) primary-dry, dominated by large diameter deciduous trees retaining less than 50% of their leaves and characterized by a forest floor with considerable accumulated dry leaf matter; 4) secondary-dry, sharing the vegetation type of its primary counterpart but with much fewer and smaller tree species; 5) wet-dry transition, characterized by both wet and dry secondary forest species on habitat fronts; and 6) scrub growth, defined as a low and tangled mixture of shrubs, vines and small trees characteristic of highly disturbed areas. In total, six transects were established, primarily through the re-clearing of existing trails, and were used for the duration of the census (Fig. 1 ). In addition, all transect distances were paced and converted to km, timed, and mapped prior to censusing.
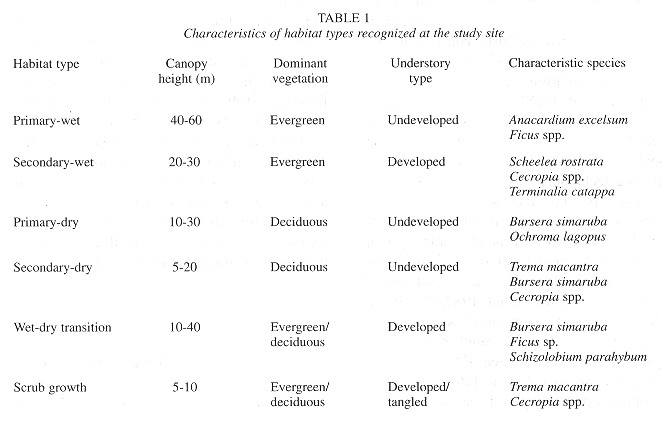
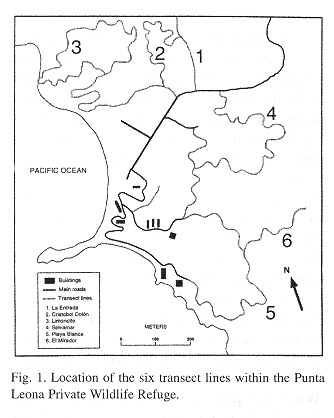
Established transects measured a total of 17.2 km and routed through all major areas of the refuge. Transects were stratified in an attempt to incorporase samples of each habitat type into the study in proportion to their representation within the refuge ( Table 2 ). Transect #1 (La Entrada) consisted of a 2.55 km section of the entrance road leading to the hotel grounds. This transect passed almost exclusively through primary-wet forest and bordered only small areas of secondary-wet forest and scrub habitat. Transect #2 (Cristóbal Colón) was 2.01 km in length and dissected a region of predominately secondary-wet forest parallel to transect # 1; it contained one relict patch of primary-wet forest and various areas of scrub growth. Transect #3 (Limoncito) consisted of large areas of secondary-dry and secondary and primary-wet forest habitats. Its 4.69 km route was used as a tourist horseback riding trail and included a small beach, later passing through the largest area of undisturbed forest in the refuge. Transect #4 (Selvamar) was located along the inside of a semi-circular ridge top encompassing a small valley of evergreen forest. Its beginning and end ran through areas of secondary-dry growth, but the majority of the 2.89 km trail passed through rich primary and secondary-wet forest. Lastly, Transects 5 and 6 (Playa Blanca, el Mirador) measuring 3.94 km and 1.11 km, respectively, were located in the southernmost extreme of the refuge ( Fig. 1 ). Transect 5 dissected all habitat types and ascended and descended the steepest local ridge; transect 6 branched off transect 5 and was the highest point in the refuge, dominated by primary and secondary-dry forest, with limited secondary-wet forest habitat.
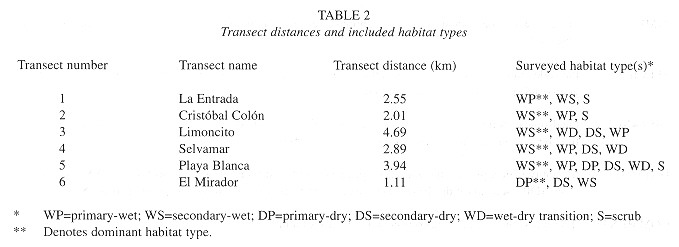
Censusing: Six mammal species were observed during the study: the white-faced monkey (Cebus capucinus), Central American spider monkey (Ateles geoffroyi), tamandua anteater (Tamandua mexicana), coati (Nasua narica), nine-banded armadillo (Dasypus novemcinctus), and variegated squirrel (Sciurus variegatoides). Although small rodents and bats were often sighted, they were not recorded and no other large mammals were sighted in the refuge during this census. Data for this study were collected from 4 to 16 March 1996. Transect walking began shortly after sunrise (approximately at 0630 hrs), when animals, particularly primates, were leaving their sleeping sites, and continued until shortly before sunset (approximately at 1800 hrs). Because animal activity sharply declined in mid-afternoon heat (1100- 1400 hrs), (Moscow and Vaughan 1987 ), transects were not walked at that time. Thus, transect lines were walked nearly 8 hrs each day for a total of 10 days. All 6 transect lines were covered daily and walking order was rotated for equal morning and afternoon coverage of each transect.
For each species the King method was applied for collecting census data (Giles 1974 , McCoy 1980 , Vaughan and McCoy 1984 ). As transects were walked, animal vocalizations and/or movements in the vegetation were carefully listened and watched for and all contacts with animals were recorded. Upon visual observation of an individual or group of animals, the date, time of day, transect number and name, species name and number of individuals, sighting number, habitat type, and sighting distance in m were recorded (Table 3 ). Here, a variation of sighting distance was used and the closest distance (diagonal or otherwise) between the observer and the animal at first sight was estimated visually. In the case that a group of individuals was sighted, the center of their occupied space was used to calculate sighting distance. Elapsed observation time and vertical location in the canopy were also documented. Animals were observed as long as necessary to ensure count accuracy and transect lines were sometimes left to identify animals. Whenever possible, group composition and individual sex and age were describes. All animal contacts were also mapped according to sighting number and species. Estimations of population density for each species were calculated according to sighting distances (x) , transect lengths (y), and numbers of individuals sighted (n) according to the King method with the following algorithm:



Results
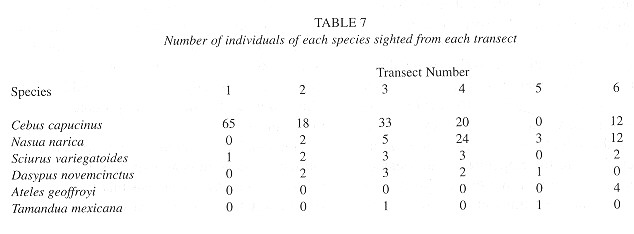
During 77 hours and 172 km of censusing, a total of 219 animals were sighted which belonged to 66 sighted groups ( Table 4 ). Sighting distances ranged from 1 to 90 m, and mean sighting distances varied considerably between species and transects.
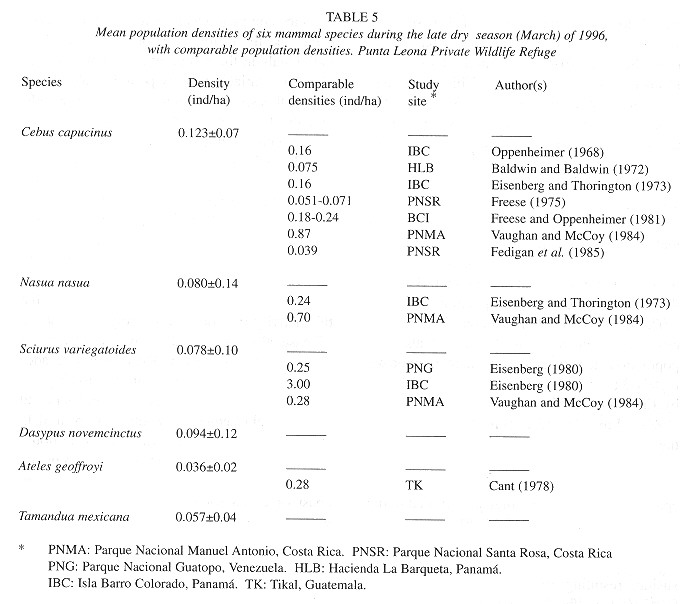
Population densities: White-faced mon keys (Cebus capucinus) and coatis (Nasua narica) were the most abundant species during this study, with 148 and 46 sighted individuals, respectively. Variegated squirrels (Sciurus variegatoides) and nine-banded armadillos (Dasypus novemcinctus) were sighted less frequently, with 12 and 8 individuals, respectively, and 4 spider monkeys (Ateles geoffroyi) and 2 tamandua anteaters (Tamandua mexicana) were also recorded. At 0.123 ind/ha, whitefaced monkeys had the largest density within the refuge, followed by nine-banded armadillos at 0.094 ind/ha and coatis at 0.080 ind/ha (Table 5 ). The population densities of variegated squirrels and tamandua anteaters within the refuge were noticeably less, at 0.078 ind/ha and 0.057 ind/ha, respectively. Spider monkeys had the smallest population density, at 0.036 ind/ha. Estimated population density values and a known study area of 300 ha were used to arrive at estimations of population size within the refuge for each species (Fig. 2 ).
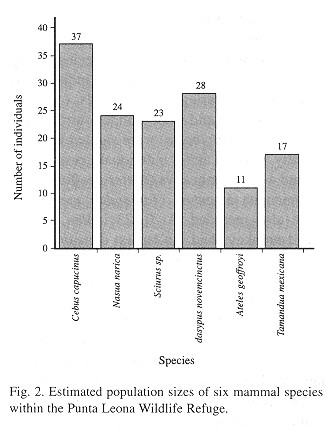
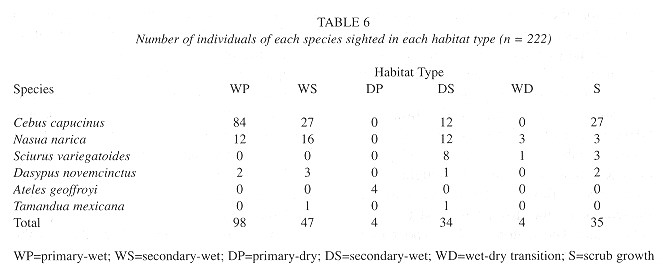
Habitat use. Mammals were observed in all six describes habitat types. The majority of white-faced monkeys were sighted in primary-wet forest (56%), while 18% of sightings occurted in secondary-wet forest, 18% in scrub growth, and 8% were observed in secondary-dry forest (Table 6 ). Similarly, a considerable percentage of coati sightings were recorded in the primary-wet (26%) and secondary-wet forest (34%). A total of 67% of variegated squirrel 34%). A total of 67% of variegated squirrel (34%). A total of 67% of variegated squirrel sightings ocurred were in secondary-dry habitat, with lesser numbers being found in scrub growth (25%) and wet-dry transition forest (8%). Small numbers of nine-banded armadillos were found in a variety of habitat types. Of the two tamandua anteaters sighted during this census, one inhabited the secondary-wet forest and the other was observed in the secondary-dry forest. The four recorded spider monkeys were all observed in areas of primary-dry forest.
Activity periods. White-faced monkeys were significantly more active during the early morning (0500-0900 hrs) and late afternoon (1600-1800 hrs) than during the other two time periods of the day (Kruskal-Wallis test, H=9.0, d.f.=9, p=0.437). Coatis were also sighted more between 0500-0900 hrs than later in the day (Kruskal-Wallis test, H=9.0, d.L=9, p=0.236). The majority (56%) of variegated squirrel sightings occurred from 0900-1200 hrs, 25% from 1600-1800 hrs, and 17% from 0500-0900 hrs. No variegated squirrels were sighted between 1400-1600 hrs. Of all nine-banded armadillo sightings, 50% were recorded during the late morning (0900-1200 hrs), 37% in the early morning (0500-0900 hrs), and 13% during the early afternoon period (1400-1600 hrs). Lastly, the two recorded tamanduas were observed in the early morning and late afternoon, respectively, and all four spider monkeys were seen soon before dusk, between 1600-1800 hrs. As previously stated, transects were not walked from 1100-1400 hrs, when the environmental temperature of the refuge was higher and general animal activity was lower.
Discussion
Cebus capucinus: The calculation of 0.123+-0.07 ind/ha is mid-range or low for C. capucinus when compared to other studies (Table 5 ). This could be a result of the relatively scarce undisturbed habitat within the refuge or a reflection of troop structure. Here, the calculation of population density does not consider C. capucinus home range nor the average number of individuals contained in a troop. However, based on sightings and sighting times, five troops of 15-30 individuals are believed to inhabit the refuge, leading to a crude estimated population size of near 100 individuals. This number contrasts sharply with the estimated population size of 37 individuals based solely on species density, but is more reliable. Two C. capucinus troops are thought to occupy territories adjacent to transect 1, while a large troop of approximately 30 individuals occupies an area near transect 3, and two more troops may overlap territories off transects 4 and 5.
Ateles geoffroyi: Only four spider monkeys were observed during this census, resulting in a population density of 0.036±0.02 ind/ha. Compared to 0.28 ind/ha, the density calculated by Cant (1978) , this value is quite low (Table 5 ). However, very few population size studies have been conducted on Ateles geoffroyi, and a range of comparable density values is not yet available. Freese et al. (1982) measured primate densities in parts of Peru and Bolivia and arrived at population density values of 0.80 ind/ha, 0.305 ind/ha, and 0.02 ind/ha for Ateles caraya, Ateles seniculus, and Ateles paniscus, respectively.
Based on the population density of this study, 11 spider monkeys should inhabit the refuge. However, only, four individuals have been observes and the indirect estimate is high. lt is interesting that the four individuals were found in primary forest. This is consistent with habits of spider monkeys elsewhere and an important factor making it the most endangered Costa Rican non-human primate species.
Nasua narica: The estimation of 0.080-+0.14 ind/ha is 3 times less than that estimated by Eisenberg and Thorington (1973) and 9 times less than that found by Vaughan and McCoy (1984) (Table 5 ). Difference may be accounted for by the distinct method used by Eisenberg and Thorington (1973) , who based their calculations on group home range, yet Vaughan and McCoy (1984) employed the King method used here. Based on the species density of this study, a population of 24 individuals is estimated to inhabit the refuge. N. narica is one of the most commonly observes mammals in Punta Leona, and this estimation of population size is thought to be considerably low. Discrepancies may be the result of the varying period of activity of this animal. Though primarily diurnal, coatis are known to become more nocturnal in areas of human activity (Kaufmann 1983 ) and such a shift in activity period could severely affect the accuracy of species densities and populations calculated from a day time census.
Dasypus novemcinctus: According to the present work, an estimated 29 nine-banded armadillos inhabit the refuge. This is based on a species density of 0.094-+0.12 ind/ha, which may be the first calculated density for D. novemcinctus in Costa Rica. This species had a mean sighting distance of 5.8±3.3 m, the shortest of any species in this study, which accounts for the large size of the estimated population. Ibis value may even be conservative, in that these small, solitary mammals were silent and discrete when stationary or traveling. All sighted individuals were foraging noisily in leaf matter on the forest floor.
Sciurus variegatoides: Variegated squirrels were sighted from all but one transect. Based on the 12 animals seen and a population density of 0.078±0.10 ind/ha, Punta Leona supports an estimated population of 23 individuals. The population density calculated here is approximately 3 times less than similar values obtained by Eisenberg (1980) and Vaughan and McCoy (1984) (Table 5 ) and is considerably less than the 3.00 ind/ha arrived at by Eisenberg (1980) in Isla Barro Colorado, Panamá. In general, populations of Sciurus vatiegatoides. were widely spaced throughout the refuge and limited to dry forest and scrub habitat (Table 6 ).
Tamandua mexicana: Only two tamanduas, or banded anteaters, were sighted during censusing, resulting in a population density of 0.057+-0.04 ind/ha. No other studies have calculated densities for this species in Costa Rica. The mixture of wet and dry forest within Punta Leona and the large number of ant and termite colonies comprise ideal conditions for T mexicana, and the estimated population size of 17 individuals in the refuge may be low. As with N. nasua, variable activity periods make densities for this species difficult to measure, as individuals of T mexicana may he diurnal, nocturnal, or both ( Montgomery and Lubin 1977 ).
Punta Leona contains two sympatric primate species: Cebus capucinus and Ateles geoffroyi. Although refuge staff members say that howler monkeys (Allouatta palliata) were formally abundant in the area, none were heard or seen during this study and they now appear to be locally extinct. lt is said that in years past, poaching occurred within the refuge and that this activity included the hunting of monkeys. Disease also decimated many populations of howler monkeys in Costa Rica severas decades ago. Subsistence hunting continues to be practiced in the area and primate populations in and around Punta Leona may continue to be at risk. Prior to this study there was also some doubt regarding the northern limit of squirrel monkey (Saimiri oerstedii) populations on the western coast of Costa Rica and whether or not they were present within the refuge. None were observed during this census and it is likely that they occur only farther south.
Although a number of people within the refuge have reported seeing sloths, neither the presence of two-toed (Choleopus hoffmanni) nor three-toed (Bradypus variegatus) sloths were witnessed during this census. Sloths are one of the most important vertebrate primary consumers in neotropical wet forests (Montgomery 1983 ) and are known to have one of the most dense distributions ( Eisenberg and Thorington 1973 ). The exclusion of inconspicuous or evasive animals such as sloths and deer (which have also been reported in the refuge) is one shortcoming of indirect censusing methods such as the King method used here. Another disadvantage is not observing nocturnal species, such as the common opossum, Didelphis marsupialis (Vaughan and Hawkins 1999 ).
Advantages and disadvantages of the King method. Indirect methods for estimating species densities within a given area are usually based on the measure of a portion of the total population of a species. As such, factors related to the ecology of the studied species and the study area may compromise accuracy. The varied behavior of animal species, existence of complex habitat stratification and terrain difficulties, and inconsistencies on the part of observers may al¡ serve as sources of error.
One of the most important factors affecting any censusing technique is visibility. Densities on which estimated population sizes are based rely on the ability of the observer to perceive animals present in the forest. In this study, visibility varied greatly, and was affected by several factors: the type of habitat, the vertical position of the animal in the canopy, weather conditions and the time of day. The thin understory of undisturbed forest generally facilitated the sighting of animals. In contrast, animals were often heard in thick undergrowth but could not be identified. Visibility of arboreal animals depended on leaf size and branch structure of the occupied tree and decreased light conditions of the early morning and dusk made identification of animal species difficult.
Differences in transect character and location may have also led to inaccuracies. Transects differed in terms of the number of species and individuals observed from each one. Although attempts were made to equally survey all habitat types within the refuge, the extent of exposure to human activity (some transects were cleared through undisturbed forest; others consisted of old roads or currently used roads) and degree of deforestation varied among transects. In addition, the relative steepness of the transect affected the velocity an observer could walk it, and thus may have affected the amount of time devoted to searching through the surrounding forest and censusing technique.
Lastly, the species observed in this census were not equally visible. The presence of Dasypus novemcinctus, for example, was not easily recorded due to the discrete nature of its behavior (Vaughan and Schoenfelder 1999 ) and smaller mammals such as squirrels could not be easily seen unless they were moving. Other species, such as Tamandua mexicana and Nasua narica were large and obvious, but their activity patterns may have interfered with the accuracy of population densities, since their periods of greatest activity likely did not coincide with the timing of the census. For slow and camouflaged species such as sloths, which were not seen here, other indirect methods of estimating densities, such as the counting of feces, nests, or tracks, could be employed. Despite the fact that primates may flee or become quiet on approach, primate species generally lend themselves to this type of study. Both Cebus capucinus and Ateles geoffroyi made considerable noise while traveling through the canopy and were easily located due to their size and numbers.
Because of unknown activity periods, indirect censusing methods such as the King method provide a limited measure of species diversity. They do, however, provide useful estimates of species abundance, in the form of population densities from which estimated population sizes can be calculated. In this study, a variation of sighting distance was used which ensured a conservative estimate of population sizes. The utilization of the closest distance between animal and observer at fírst sight, instead of strictly perpendicular sighting distances, decreases the chances of overestimating populations and results in more protective, rather than passive, wildlife management plans. This variation is also more practical on steep and winding transects, where obtaining perpendicular measures of sighting distance is difficult.
This study was conducted by a single observer, which decreases efficiency of time and expense, but largely eliminates human error. When numerous observers are used, differences in ability to locate animals, calculate sighting distances, and determine the number of animals in groups lead to error which is further increased by the small number of observations made by each observer (Vaughan and McCoy 1984 ). In addition, here transects were walked and population densities were determined for 10 days and mean values were used to arrive at population sizes. The averaging of replicates greatly improves accuracy and precision and accounts for daily variation in animal activity due to temperature and weather. Further, it is important that the sample be representative, and that transects be stratified by habitat type in proportion to their inclusion in the study area. Transect distances must also be accurately measured, as small variations in distance can lead to great error in density values.
Despite some limitations, the estimates of population size and density obtained in this study present certain advantages. The King method is relatively simple compared to other indirect censusing methods and is much more practical than direct censusing in large areas with appreciable animal populations. Because censuses employing the King method can be conducted relatively fast, it is possible to conduct seasonal or yearly censuses in the same area or compare population densities between different regions with a minimum of time and effort.
Although population sizes given here are conservative, the low diversity and relatively low abundance of primate and mammal species in Punta Leona may be cause for concern. The refuge is in severas ways an island, since ¡t borders the ocean, is cut off from the mainland by highway, and is otherwise surrounded by pastureland or privately owned forest. In particular, the sighting of only tour spider monkeys during this census is alarming. Refuge personnel recall larger numbers in the past and increased human activity may be responsable for migration out of the refuge. Punta Leona Hotel and Club continues to grow and a previously isolated portion of forest adjacent to where the spider monkeys were sighted has been recently cleared and developed. Reproductive rates of wild populations of Ateles geoffroyi are very low and the genus appears to be extremely sensitive to human perturbation of habitat. lt may be one of the first primate species to decline with severe disturbance (Eisenberg 1983), and this has already occurred in Cabo Blanco Absolute Nature Reserve, Costa Rica, where A. geoffroyi is now locally extinct (Lippold 1989 ). This study underlines the importance of primary forest, primarily wet forest, to neotropical mammal populations and to primates in particular. As an indicator species, the predicament of A. geoffroyi suggests that Punta Leona Hotel and Club management would be wise to re-evaluate local wildlife management strategy, especially given that a large number of their clientele are attracted by the surrounding forest and its wildlife.
Acknowledgements
We thank the administrators and staff of the Punta Leona Hotel and Club for their kind permission to work within the refuge and use their facilities. We are endebted to all the people at the Associated Colleges of the Midwest, who provided much needed personal and logistical support. We thank Anne Houde for her endless help and shared knowledge of animal behavior and statistics and William Moskoff and Lynn Westley for their valuable comments. In addition, we are also grateful to the Barrenechea and Chavez families for their generous support and unconditional friendship.
Resumen
Se estimó el tamaño poblacional de seis especies de mamíferos, usando el método de King, durante la estación seca tardía (marzo) de 1996 en el Refugio de Vida Silvestre de Punta Leona, Costa Rica. El mono carablanca (Cebus capucinus), el pizote (Nasua narica) y el armadillo de 9 bandas (Dasypus novecinctus) presentaron las poblaciones más grandes en el refugio, con 148, 46, y 8 observaciones respectivamente. El mono araña centroamericano (Ateles geoffroyi), el tamandúa hormiguero (Tamandua mexicana) y la ardilla (Sciurus variegatoides) también se tomaron en cuenta durante el censo. Además se calculan las densidades poblacionales, se describen los tipos de hábitat y se discute el uso de estos, así como los periodos de actividad de las especies encontradas.
References
Baldwin, J. & J. Baldwin. 1972. The ecology and behavior of squirrel monkeys (Saimiri oerstedii) in a natural forest in western Panamá. Folia Primatol. 18: 161-182. [ Links ]
Cant, J.G.H. 1978. Population survey oí the spider monkey Ateles geoffroyi at Tikal, Guatemala. Primates 19: 525-535.
Chapman, C., L. Fedigan & L. Fedigan. 1988. A comparison of transect methods of estimating population densities of Costa Rican primates. Brenesia 30: 67-80. [ Links ]
Defler, T. & D. Pintor. 1985. Censusing primates by transect in a forest of known primate density. Int. J. Primatol. 6: 243-249. [ Links ]
Eisenberg, J. 1980. The density and biomass of tropical mammals. pp. 35-55. In M. Soulé & B. Wilcox (eds.). Conservation Biology. Sinauer, Massachusetts. 395 p. [ Links ]
Eisenherg, J. & R. Thorington. 1973. A preliminary analysis of neotropical mammal fauna. Biotropica 5: 150-161. [ Links ]
Freese, C.H. 1975. Censusing Alouatta palliata, Ateles geoffroyi, and Cebus calcinas in the Costa Rican dry forest. pp. 4-9. In R.W. Thoringtyon, Jr. & R Heltne (eds.). Neotropical primates: Field studies and conservation. National Academy of Sciences, Washington. [ Links ]
Freese, C.H., P.G. Heltne, N. Castro & G. Whitesides. 1982. Pattems and determinants of monkey densities in Peru and Bolivia, with notes on distributions. Int. J.Primatol. 3: 53-90. [ Links ]
Freese, C.H. & Oppenheimer. 1981. The capuchin monkey: genus Cebus sp. pp. In A. Coibra-Filho and R. Mittermeier (eds.). Ecology and behavior of neotropical primates. Vol. 1. Academia Brasileira de Ciencias, Río de Janeiro. [ Links ]
Giles, R. 1974. Wildlife management techniques. The Wildlife Society, Washington, D.C. 633 p. [ Links ]
Janzen, D. 1983. Costa Rican Natural History. University of Chicago, Chicago, IL. 816 p. [ Links ]
Kaufmann, J. 1983. Nasua narica (Pizote, Coati). pp. 478480. In D. H. Janzen (ed.). Costa Rican Natural History. University Chicago, Chicago, IL. 816 p. [ Links ]
Lippold, L. 1989. A census of primates in Cabo Blanco Absolute Nature Reserve, Costa Rica. Brenesia 31: 93-97. [ Links ]
McCoy, M. 1980. Inventario animal - transecto - método King. Universidad Nacional, Heredia, Costa Rica. 9 p. [ Links ]
Montgomery, G. 1983. Bradypus variegatus (Perezoso de Tres Dedos, Three-toed Sloth). pp. 453-456. In D.H. Janzen (ed.). Costa Rican Natural History. University Chicago, Chicago, ¡L. 816 p. [ Links ]
Montgomery, G., & Y Lubin. 1977. Prey influences on movements of neotropical anteaters. pp. 103-131. In R.L. Phillips & C. Jonkel (eds.). Proceedings of the 1975 Predator Symposium. Montana Forest and Conservation Experiment Station, University of Montana. [ Links ]
Moscow, D. & C. Vaughan. 1987. Troop movement and food habits of white-faced monkeys in a tropical-dry forest. Rev. Biol. Trop. 35: 287-297. [ Links ]
Oppenheimer, J.R. 1968. Behavior and ecology of the white-faced monkey, Cebus capucinus. Ph.D. thesis. University of Illinois, Urbana. 181 p. [ Links ]
Sokal, R. & F. Rohlf. 1981. Biometry. Second edition. W.H. Freeman, San Francisco, CA. 859 p. [ Links ]
Vaughan, C. & L. Hawkins. 1999. Late dry season habitat use of common opossum, Didelphis marsupialis (Marsupialia: Didelphidae) in neotropical lowermontane agricultural areas. Rev. Biol. Trop. 47: 263-269. [ Links ]
Vaughan, C. & M. McCoy. 1984. Estimación de las poblaciones de algunos mamíferos en el Parque Nacional Manuel Antonio, Costa Rica. Brenesia 22: 207-217. [ Links ]
Vaughan, C. & C. Rodriguez. 1997. Managing beyond the borders: The Costa Rican System of Conservation Areas. In G. Meffe & R. Carroll (eds.). Principles of Conservation Biology. Sinauer, Sunderland, MA, USA. [ Links ]
Vaughan, C. & S. Shoenfelder. 1999. Dry season activity, movement, habitat and den utilization of nine-banded armadillo (Dasypus novemcinctus) in neotropical dry forest, Costa Rica. Rev. Biol. Trop. 47: 1117-1119. [ Links ]
Williams, H. & C. Vaughan. 2001. White-faced monkey (Cebus capucinus) ecology and management in neotropical agricultural landscapes during the dry season. Rev. Biol. Trop. 49: 1199-1206. [ Links ]
Wilson, W. & A. Johns. 1982. Diversity and abundance of selected animal species in undisturbed forest, selectively logged forest and plantations in East Kalimantan, Indonesia. Biol. Conserv. 24: 205-218. [ Links ]
1 . Associated Colleges of the Midwest, Apartado 10265, San José, Costa Rica
2 . Dept. of Wildlife Ecology, University of Wisconsin, Madison, WI 53706, USA; cvaughan@wisc.edu
3 . Regional Wildlife Management Program, Universidad Nacional, Heredia, Costa Rica.

