










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.3-4 San José Dec. 2002
Abstract
The West Indian Topshell Cittarium pica is artisanally collected on rocky shores along the Caribbean Coast of Costa Rica. There are neither data on the state of its exploitation nor exist any regulation of the fishery. From October 2000 to March 2001, the population dynamics of this species were studied at an unexploited and two exploited sites to determine the present impact of the fishery on the resource. Average population density with 14 ind./m2 about three times higher at the unexploited than at the exploited sites. Length-frequeney histograms showed a strong shift towards smaller specimens at the exploited sites, which is also reflected in significantly higher rates of total mortality (Z = 4.05 and 4.47) when compared to the unexploited site (Z = 1.47). Von Bertalanffy growth parameters were estimated as k = 0.19-0.28 (yr-1) and L![]() = 104 mm. No significant differences were found among sites. From these values a range of the growth performance index
= 104 mm. No significant differences were found among sites. From these values a range of the growth performance index ![]() was computed (
was computed (![]() = 3.31-3.48) which lies at the lower end of the values reported for other tropical marine gastropods. The size at first maturity for both sexes combined was estimated as 29.20 ± 1.14 mm. Exploitation rates >0.6 for both exploited sites and a large fraction of small specimens (<30mm) in the catches suggest overexploitation and recruitment overfishing. Based on the estimated maximum sustainable yield we recommend regulative measures for the fishery such as a control of a minimum landing size of 40 mm and a closure of the fishery during its reproductive period (from July to November).
= 3.31-3.48) which lies at the lower end of the values reported for other tropical marine gastropods. The size at first maturity for both sexes combined was estimated as 29.20 ± 1.14 mm. Exploitation rates >0.6 for both exploited sites and a large fraction of small specimens (<30mm) in the catches suggest overexploitation and recruitment overfishing. Based on the estimated maximum sustainable yield we recommend regulative measures for the fishery such as a control of a minimum landing size of 40 mm and a closure of the fishery during its reproductive period (from July to November).
Key Words
Cittarium pica, Gastropods, ELEFAN, Fishery, Population Ecology, Caribbean, Costa Rica.
The gastropod Cittarium pica (= Livona pica, L. 1758) is fished throughout its whole distribution range in the Caribbean, where it used to rank second after the conch Strombus gigas (L.) as a shellfish-resource (Randall 1964 ). Bell (1992) and Cervigón et al. (1993) , however, stated a heavy overexploitation of this species in recent years and observes its disappearance from the markets. A management program for this species is only known from the Virgin Islands and Puerto Rico. In the Virgin Islands the minimum landing size is 62 mm in shell length and the fishery is closed between April and September (Mateo, pers. comm.), while in Puerto Rico the minimum landing size is about 63 mm (Lilyestroem, pers. comm.).
In Costa Rica the whelk, as the species is commonly called here, is hand-collected on a small scale and not commercialised at the public market (INCOPESCA, pers. comm.). For this reason no information is available on the present condition of the stock. The objectives of this study were to determine the condition of the stock, to ascertain the level of exploitation and to contribute with recommendations for an adequate management of this species in Costa Rica.
Material and methods
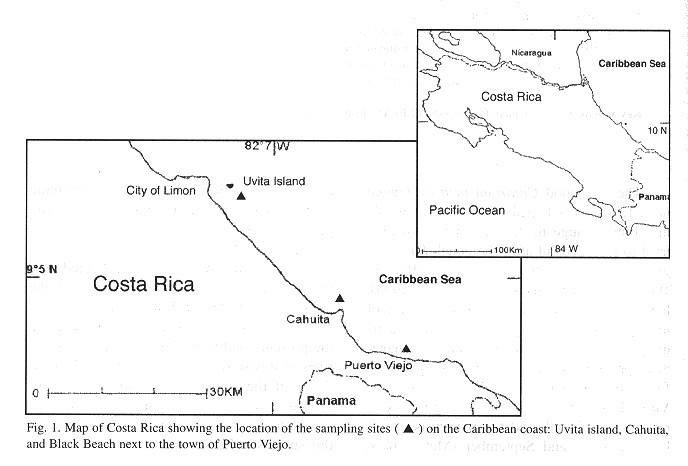
Study area and sampling sites: The Caribbean coast of Costa Rica stretches between Nicaragua to the North and Panama to the South and belongs to the Southern part of the Caribbean Sea (Fig. 1 ). lt is about 200 km long and except for the small island of Uvita in front of the port city of Limón (10º N, 83º W) and three points in the Southern half the coast (Fig. 1 ) the coast is relatively straight and dominated by sandy beaches, rocky shores, and river mouths. Tides are diurnal with a range of about 0.5 m ( Jiménez 2001 ).
The rocky part of the coast is limited to severas patches between Moín and Manzanillo (Fig. 1 ) which are only a few hundred m long and about 10 m wide. Most of these rocky patches consist of living and fossil reefs full of holes and crevices. The coral patches on the Caribbean coast of Costa Rica are impacted by siltation and bleaching (Cortés and Risk 1985 ; Jiménez 2001 ).
C. pica can be found at the majority of these patches but densities are generally low (<10 ind./m 2 in most areas). Within this stretch of rocky patches along the coast, the Uvita island, located about 1 km offshore from the port city of Limón (Fig. 1 ) was chosen as an unexploited site. Uvita has the status of a National Monument (Columbus anchored there in 1502) and as such the collection of animals and plants is prohibited.
For the assessment of the actual impact of the fishery, one unexploited and two exploited sites were compared in terms of population density, length-frequency distribution and mortality. Additionally, growth and mortality rate, the size at first maturity and a sustainable relative yield per recruit were determined to understand the population dynamics and production characteristics of this species in Costa Rica and to provide the scientific basis for fishery recommendations.
lt was assumed that low population densities and smaller individuals are to be found in areas of more intense fishing and that the comparison of areas of different fishing impact would allow for a better understanding of the population dynamics of this species as influenced by the fishery (Moreno et al. 1984 , Castilla and Duran 1985 , Hockey and Bosman 1986 , Ortega 1987 , Keough et al. 1993 ). In addition, fishery operation and catch volumes were recorded and people involved in the fishery were interviewed for their knowledge and expertise as resource users (Schoenhuth and Kievelitz 1993 , Pomeroy 1995 , Debrot 2000 , Ruddle 1994 ).
As exploited sites, two rocky areas of a length of about 1 km near the small village of Cahuita, 40 km South-East of Limón were chosen (Fig. 1 ). All three sites were selected due to their great similarity in environmental characteristies, such as a shore slope <10º, wave action and the substrate consisting of fossil reef and similar species composition of algae and invertebrates. Hereinafter these sites are referred to as Uvita, Cahuita, and Black Beach.
Density and size distribution: A total of 295 randomly allocated squares (1 m x 1 m) were counted along transects perpendicular to the coastline, for the presence of C. pica (95 at Black Beach, 96 at Cahuita, and 104 at Uvita), with a 1 m 2 plastic frame. To prevent double counting of snails moving through the habitat, the transects were placed at a minimum distance of 2 m between each other. To obtain the average snail density at each site, the number of specimens of all squares was divided by the total number of squares counted:
Additionally, length-frequency histograms were constructed from the measurements of the snails obtained from each site. Shell length was measured across the widest diameter of the shell base ( Debrot 1990b ) using a calliper with an accuracy of 0.01 mm. Due to the prevailing high wave conditions, the lower part of the shore edge and the vertical drop zone of the shore, where large specimens (>60 mm) are concentrated ( Randall 1964 , Debrot 1990a , pers. observations), could not adequately be sampled at any of the three sites. Hence, they are not included in the calculations.
Growth: Length frequency data were also used to calculase the von Bertalanffy growth rate 'k' and the asymptotic length 'L![]() ' by model progression analysis using the FISAT programme ( Sparre and Venema 1992 ) The non-seasonalized von Bertalanffy equation was used as samples were only taken during one period of the year (rainy season):
' by model progression analysis using the FISAT programme ( Sparre and Venema 1992 ) The non-seasonalized von Bertalanffy equation was used as samples were only taken during one period of the year (rainy season):
where L t = Length at time t; L![]() = asymptotic length; k growth constant; t o = age at length 0; here to was considered zero, as any specific information en absolute age at length t o was not available (Appeldoom 1988).
= asymptotic length; k growth constant; t o = age at length 0; here to was considered zero, as any specific information en absolute age at length t o was not available (Appeldoom 1988).
Based on the findings of Debrot (1990b) and Randall (1964) the asymptotic length 'L![]() ' was assumed to range between 90 mm and 110 mm and the instantaneous growth rate 'k' between 0.1 and 0.6. These value ranges were used as input for the FISAT program and the best possible fit of the growth curve was obtained by first using the "surface analysis" and then the "automatic search routine" of the ELEFAN 1 program (Sparre and Venema 1992 ).
' was assumed to range between 90 mm and 110 mm and the instantaneous growth rate 'k' between 0.1 and 0.6. These value ranges were used as input for the FISAT program and the best possible fit of the growth curve was obtained by first using the "surface analysis" and then the "automatic search routine" of the ELEFAN 1 program (Sparre and Venema 1992 ).
As different cohorts could not clearly be distinguished visually in the Cahuita samples, the Bhattacharya-method of the ELEFAN 1 program was used to visualize the different cohorts in this sample.
In addition, 546 snails were tagged in Cahuita and the Munro-plot was used to calculate the growth rate using the formula:
with k = growth constant; L![]() = asymptotic length as calculated with the length-frequencies; L 1= length at tagging; L 2 = length at recapture, t 2 - t 1 = time interval between tagging and recapture (Koch and Wolff 1996 ).
= asymptotic length as calculated with the length-frequencies; L 1= length at tagging; L 2 = length at recapture, t 2 - t 1 = time interval between tagging and recapture (Koch and Wolff 1996 ).
The whelks were marked with nail polish of a colour similar to red calcareous algae often overgrowing the shells of Cittarium pica. To distinguish between single individuals, a number was written with a water-resistant ink on the nail polish. Finally, glue was put on top to seal the tag, following Koch (1992) . The animals were recaptured after three weeks on average, measured with a calliper and repainted as the tag tended to fade after three weeks.
New snails were marked at each recapture date to compensase for losses due to mortality, collection, or tag shedding. To reduce emigration of the tagged specimens all marked snails were always released at the same point (Appeldoom 1984 ).
The growth performance index ![]() was calculated from the growth parameter estimates because it facilitates the intra and interspecific comparison of the growth performance ( Pauly and Munro 1984 ):
was calculated from the growth parameter estimates because it facilitates the intra and interspecific comparison of the growth performance ( Pauly and Munro 1984 ):
with k = growth constant and L![]() = asymptotic length.
= asymptotic length.
Mortality: Total mortality "Z" : For the calculation of the instantaneous annual mortality rate "Z" the length converted catch curve (Pauly 1983 , Munro 1984 ) was applied to the pooled length frequency data of the different sites using the estimated growth parameter. With this method 'Z' is calculated from the right descending arm of the curve:
with C = number of specimens, L 1; L 2 = length at time point t 1; t 2, ![]() t = time interval between L1 and L2 , c =conversion factor and Z = mortality rate. The calculation was done with the FISAT program (Sparre and Venema 1992 ).
t = time interval between L1 and L2 , c =conversion factor and Z = mortality rate. The calculation was done with the FISAT program (Sparre and Venema 1992 ).
Natural mortality "M" and fishing mortality "F": At the unexploited site Uvita the fishing mortality was assumed as F=0 and thus resulting in M=Z. For the calculation of the fishery mortality "F" at the two fished sites Cabuita and Black Beach, the above M value was subtracted from the Z- value in order to get the fishing mortality (F=Z-M) (Sainsbury 1982a , Appeldoom 1984 ,1988 ).
Exploitation rate "E": With the known values of 'F' and 'Z' the exploitation rate 'E' was calculated according to (Sparre and Venema 1992 ):
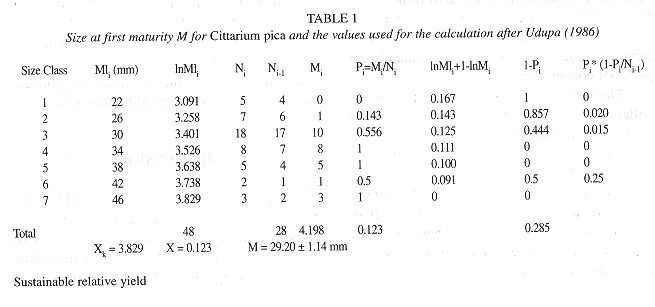
Size at first maturity: The mean size at first maturity was obtained following a method suggested by Udupa (1986) . which also allows for the calculation of the 95% confidente intervals around the estimate. The sample is organised in size classes and the formula of Spearman-Karber is used to calculase the size at first maturity 'M':
with: Xk = In of first size at which 100% of individuals are fully mature; X = In of the average size increment; MI = Midlength per size class; In Ml i+1 - Mli +1 - Ml 1; P i = Proportion of fully mature individuals per size class; Ni = Total number of individuals per size class.
The 95% confidence intervals were calculated with the same parameters as above and the formula:
Sexing of mature specimens was done by visual inspection of the gonad colour (Randall 1964 , Bell 1992 ). The data of both sexes and of all sites were pooled since the total number of mature specimens in the samples was quite low with 4 from Cahuita, 13 from Black Beach and 14 from Isla Uvita.
Relative sustainable yield: The relative yield per recruit was calculated with the FISAT program (Sparre and Venema 1992). Input values were the mean value of the estimated growth rates k = 0.235, the asymptotic length L![]() = 104mm, the natural mortality M 1.47 and the length at first capture Lc.
= 104mm, the natural mortality M 1.47 and the length at first capture Lc.
The relative yield per recruit was calculated first for the actual length at first capture Lc = 25 mm and the exploitation rate E = 0.65 being the mean of the exploitation rates at the two sites.
Then the length at first capture was modified within a range above the size at first maturity that would allow the snails to reproduce at least once before collection (Lc>30mm) to find a size limitation for the fishery that would provide a sustainable yield per recruit.
The fishing mortality itself was not varied because a size limitation for the collection is more likely to be accepted than a limitation of the number of snails being taken.
Observations on the fishery: Catch volumes were registered and fishery operation was observed by accompanying fishermen on several occasions. The quantity and composition of the catches was registered and interviews were done in an informal manner without distributing questionnaires. lt was taken care of asking similar questions (on topics listed below) thus the obtained information could be cross-checked. The topics covered in the interviews are given below. Order and exact formulation of the questions depended on the situation and the interviewed person.
- Knowledge about the species.
- Temporal dynamics of fishery and snails.
- Fishery operation.
- Social position of the interviewee.
- Yield success and utilization.
- Predictions for fishery development.
- Market issues.
- Predictions for snail dynamics over time.
Results
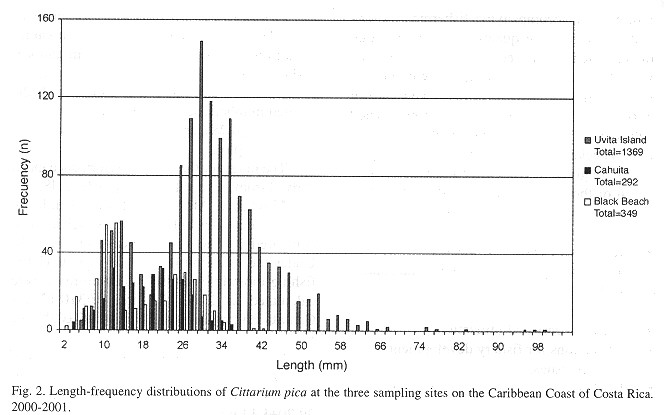
Density and Size distribution: Average densities at the exploited sites (Cahuita and Black Beach) are 3±3 and 4±2, respectively being three times lower than at the unexploited site Uvita (14±5). The length frequency histograms show a significantly higher proportion of smaller specimens at the exploited sites (F-test, p<0,05), (Fig. 2 ).
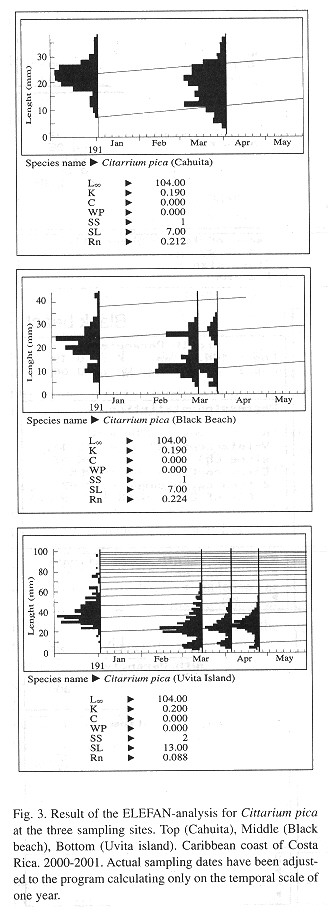
Growth: The model progression analysis using the ELEFAN method gave similar values for all three sites with L ![]() = 104 mm and k = 0.19- 0.20 ( Fig. 3 )
= 104 mm and k = 0.19- 0.20 ( Fig. 3 )
Based on 268 recaptures from which the painted number could be identified, a mean k-value of = 0.28 per year was calculated with the Munro-plot.
The range for the growth performance index derived from these values is: ![]() = 3.31- 3.48.
= 3.31- 3.48.
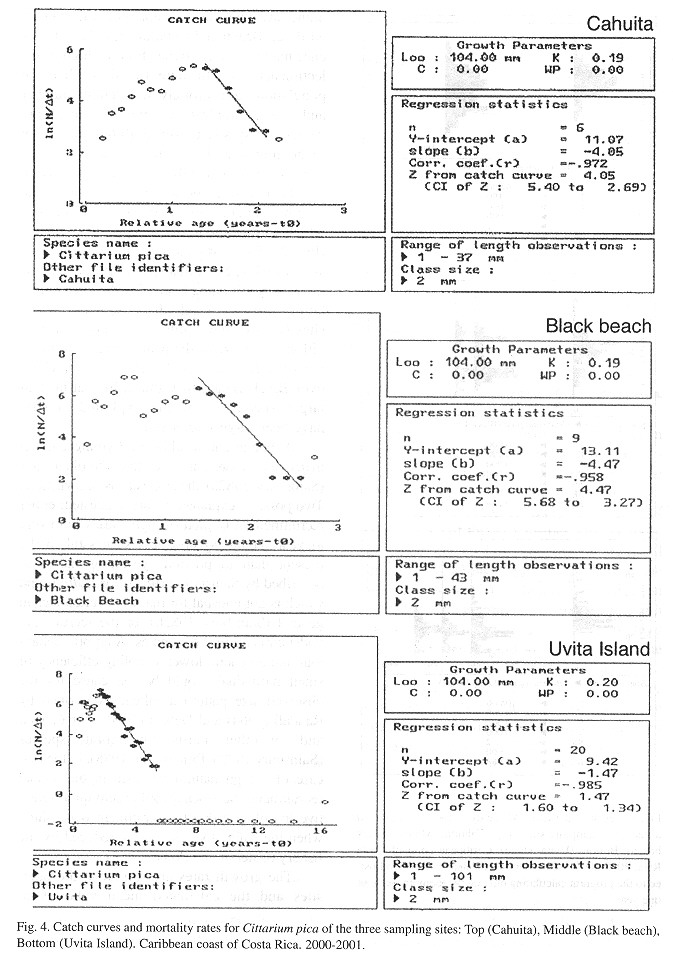
Mortality: Mortality rates are similar at the exploited sites but three times higher when compared to the unexploited site (Fig. 4 ). Under the assumption that M = Z at Uvita, fishing mortalities and exploitation rates were calculated as 2.59±1.35 and 0.63±0.12 for Cahuita and as 3.00±1.21 and 0.67±0.09 for Black Beach, respectively.
Size at first maturity: The calculation of the size at first maturity for both sexes combined is given in Table 1 . and resulted in M 29.20±1.14 mm.
Sustainable relative yield: Based on the input data the relative yield per recruit for the actual length at first capture was calculated to be 2.7 * 10-3. A sustainable relative yield was identified at the length at first capture Lc = 40 mm. This length allows the snails to reproduce and maintains a relative yield per recruit close to the actual one of 2.3 * 10-3.
Observations on the fishery: Cahuita is a small fishing village with about 4 000 people living in its total catchment area. The National Park next to Cahuita and the associated tourism are an important source of income for the village. The fishery is small scale focusing on finfish and spiny lobster. The catch is either for personal consumption or sold at the village.
There are two different types of C. pica fishery in Cahuita. One is a seasonal fishery between March and April when the ocean is calm enough to allow access to the vertical drop zone of the rocky shore below the low tide line. During this time, people from Cahuita collect bigger specimens (>40 mm) that can be found in the lower part of the shore by snorkelling around the rocks. About 100 snails with an average size of 60 mm are collected per household in one season, mainly for personal consumption.
The other type of fishery is a permanent collection of snails of 20-40 mm by people walking in the upper, horizontal zone of the rocky shore that could be accessed most of the time during low tide.
A range of 14 to 50 snails are collected per walk, and the frequency of visits to the shore to collect whelks varied strongly between once every other week to every day depending on the fishermen and weather conditions.
Mainly Iow income people are involved in the fishery. The whelks are sold to other people in the village, or are used for as food, or the shells are processed and sold as jewelry.
All people that were interviewed knew of the existence of this species but had generally very little knowledge about its biology and population dynamics. Still about 70% of the interviewees stated a decline of the population around the village over the years. People with a higher educational background in biology (e.g. natural tourist guides) related the decline to overfishing, while others attributed the decline to the 1991 earthquake (7.6 Richter scale, Denyer 1991 ) but could not explain the mechanism behind (the shoreline was uplifted and exposed more than 0.5 m, probably diminishing available submerged habitat area).
Discussion
Cittarium pica is collected either by walking or by snorkeling along the Caribbean coast of Costa Rica, except for at Uvita Island, where it is protected. Our study results show that the average snail density is three times higher at Uvita island compared to the exploited Black Beach and Cahuita sites. This difference may be related to the observed higher collection activity at the latter two sites, where the populations are strongly reduced in number and - as the largest available specimens are depleted first- size classes > 40-60 mm are not to be found at these sites, while these size classes are present at the protected site (Uvita).
Two distinct peaks can be seen in the length-frequency histograms for all three sites which correspond to the first two year classes (Fig. 2 ). The first peak is similar for all three sites which suggest that spawning and recruitment is synchronized between the sites correlating with the small distance between the sites (Fig. 1 ) and similar environmental conditions. The size of the peaks (= relative number of specimens) is decreasing from Uvita over Black Beach to Cahuita indicating that larger specimens of the exploited cohorts have been largely removed.
Although one would expect younger specimens being more numerous than the older ones (Sainsbury 1982a ) the opposite is shown here. Two possible explanations are suggested: either recruitment of C. pica strongly varies from year to year (being possibly less successful in the present than in previous years) as has been describes by Sainsbury (1982a) for Haliotis and which is not unusual for marine invertebrates in general (Sainsbury 1982b ), or the recruits are understimated in the samples. A cryptic behaviour and a general lower sampling efficiency of small individuals could be the cause for the observed size pattern as already describes by Randall (1964) and Debrot (1990a) for C. pica and for other marine invertebrate species ( Sainsbury 1982a , Prince et al. 1988 ). In the first case of a high natural variance in the annual recruitment, the stocks would seem quite sensitive to collapse due to recruitment overfishing when the adult stock is heavily reduced by the fishery ( Caterall and Poiner 1987 ).
The growth rates are similar at the other sites and the calculated mean value is in between the values found by Debrot (1990a) with k = 0.30-0.36, and k = 0.14-0.25 by Randall (1964) .
The goodness of fit for the ELEFAN growth curves is low (Fig. 3 ) and the confidence limits around the parameter estimates are correspondingly rather large. However, the estimates seem realistic, because independent calculations for all sites gave similar results and the values are comparable with literature reports for other marine gastropods ( Koch 1996 ). One further reason for the poor fit of the growth curve could be that the von Bertalanffy model is not the most suited one for describing growth in C. pica, especially for the part that concerns the younger age groups (the older and larger specimens were not quantitatively assessed in our study). A curve of a sigmoid form, with inflexion somewhere in the latter juvenile phase, might be better suited as proposed for Concholepas concholepas (Wolff 1994 ).
When the estimate for the growth performance index (![]() = 3.31-3.48) is compared with the range of values compiled by Wolff (1994) for marine gastropods of all latitudes (
= 3.31-3.48) is compared with the range of values compiled by Wolff (1994) for marine gastropods of all latitudes (![]() = 2.2-4.7) and the tropics (
= 2.2-4.7) and the tropics (![]() = 3.6-4.7), it is found that the growth performance of C. pica lies at the low end of the range observed for most other tropical marine gastropods (
= 3.6-4.7), it is found that the growth performance of C. pica lies at the low end of the range observed for most other tropical marine gastropods (![]() = 3.6-4.7). lt may be speculated that the growth rates obtained in this study are a little too low for the following reasons: field sampling took place only during the rainy (winter) season, when the waves are rough, rainfall is higher and temperatures are lower; and spawning took place during the study period in November and December followed by a high inflow of new recruits in January which might have had negatively affected growth. Possibly growth is somewhat faster in the dry season (summer) when the average water temperature is higher and the animals do not need to use energy for reproduction.
= 3.6-4.7). lt may be speculated that the growth rates obtained in this study are a little too low for the following reasons: field sampling took place only during the rainy (winter) season, when the waves are rough, rainfall is higher and temperatures are lower; and spawning took place during the study period in November and December followed by a high inflow of new recruits in January which might have had negatively affected growth. Possibly growth is somewhat faster in the dry season (summer) when the average water temperature is higher and the animals do not need to use energy for reproduction.
The natural mortality rate (M = 1.34-1.60) estimated for the population of Uvita island is higher than that estimated by Debrot (1990a) (M = 0.83) but lies in the range of values (1.01.7) for marine gastropods given by several authors (Sainsbury 1982a ,b , Appeldoom 1987 , 1988 , Debrot 1990b ). Still in relation to the relative slow growth of the species the estimate seems rather too high (less than 30% would survive one year). Emigration out of the accessible part of the habitat while growing and the tendency to hide in crevices (which results in a lower capture efficiency) can be expected to have somewhat increased the M estimate which was based on the catch curve.
In addition, it has to be considered that only small to middle-sized specimens were used for the calculation. The mortality is inversely related to size (Appeldoom 1987 ) thus the calculation of the rate was not based on a balanced set of data.
Total mortality at Uvita island is significantly lower than at Cahuita and Black Beach (Fig. 4 ), which is indicative for the strong fishing impact at these sites.
The exploitation rate (E around 0.65) calculated here suggest overexploitation (Gulland 1971). C. pica seems to be impacted in two ways at Black Beach and Cahuita, since collection of the larger size classes (>60 mm) takes place once a year on a quite intensive level (through snorkelling in the subtidal), while permanent harvest of the smaller size classes occurs on a rather low level but over the whole year's period (through walking collectors).
The size at first maturity as calculated in this study shows that immature snails are already collected. Thus a large part of the reproductively active individuals are removed. This should impact the spawning and recruitment success of the population over time ( Caterall and Poiner 1987 ) and might increase the risk of the population to collapse.
This risk exists probably not only around Cahuita but all along the Caribbean coast except for a few inaccessible spots and of course for the protected arca at Uvita island, as indicated by some preliminary sampling at severas spots between Moín and Manzanillo.
Based on the results for the calculation of a sustainable relative yield per recruit, a minimum collection size of 40 mm is recommended. The majority of new recruits is from spawnings in November and December and enter the rocky shore in January. According to the estimated growth rate they need two years before they take actively part in reproduction. Caterall and Poiner (1987) demonstrated that gastropods are not fully reproductive at the size of first maturity which is about the size the snails reach in their first year. To ensure that a sufficient number of snails reach the second spawning period in winter alter their settlement, we additionally suggest a seasonal closure between July and December. By these measures (seasonal closure of the fishery and Lc = 40 mm) and their suitable combination, the total yield can be improved because the closures increase the egg production (and subsequent recruit number). The collectors would benefit since fewer fishing visits would be needed to harvest the same biomass because snail numbers (through larger recruitment) and the average length (through higher landing size) would increase ( Sluczanowski 1984 ).
These measures would not affect the fishermen working in the subtidal, but would heavily affect those walking between the rocks, collecting year-around specimens of (mostly) smaller sizes. Therefore we suggest to determine the socio-economic impact of these measures and to assess the need for alternatives to compensase for the loss that these regulations would mean lo the fishermen.
Mariculture of this species could he an interesting alternative as already suggested by Bell (1992) . lf suitability for mariculture could be combined with a good market potential, a mariculture project could be envisioned, either in Cahuita or at another village along the coast and involving local people. In this way a new employment area could be created which would contribute to the development of the village in general and a decrease in the fishing pressure on the natural population.
Further research on the population dynamics should be done as the results of this study are based on a rather temporally and spatially limited data set. Another interesting topic for future research would be the role of the C. pica population on Uvita as a source of recruits to the mainland shore.
Acknowledgments
This study was part of a M.Sc. Thesis of the senior author within the ISATEC - programme at the University of Bremen. Field work in Costa Rica was conducted under the Memorandum of Understanding between ZMT and CIMAR.
We would like to thank the personnel of CIMAR for their logistic support and the German Agency for Academic Exchange (DAAD) for funding. Special thanks go to Rosaura Steele, (Gestión Ambiental, J.A.P.D.E.V.A.,Costa Rica) for her help and granting the permit to conduct part of this study at Uvita island, National Monument.
Resumen
El caracol Cittarium pica (West Indian Top Shell) es recolectado en forma artesanal en zonas rocosas de la costa Caribe de Costa Rica. A la fecha no hay datos sobre esta extracción ni existe regulación de su pesquería. La dinámica poblacional de esta especie fue evaluada, desde octubre del 2000 hasta marzo del 2001, en dos sitios en los cuales la especies es recolectada (Playa Negra y Cahuita), y en un sitio protegido de la actividad pesquera (Isla Uvita). La densidad promedio de la población fue 14 ind/m2, cerca de tres veces más alta en el sitio protegido que en los dos no protegidos. Los histogramas de frecuencia de tallas mostraron un fuerte sesgo hacia los ejemplares más pequeños en los sitios no protegidos, lo que se refleja también en tasas de mortalidad total significativamente más altas (Z = 4.05 y 4.47) cuando se les compara con el sitio protegido (Z = 1.47). Los parámetros de crecimiento según von Bertalanffy fueron estirnados en k = 0.19 - 0.28 / año y L![]() = 104 mm. No se encontró diferencias significativas entre los sitios. A partir de estos valores el índice
= 104 mm. No se encontró diferencias significativas entre los sitios. A partir de estos valores el índice ![]() (performance index
(performance index ![]() ) estuvo en un ámbito de 3.31 a 3.48, el cual se encuentra entre los valores bajos informados para otros gastrópodos tropicales. La edad a la primera madurez sexual para ambos sexos combinados fue estimada en 29.20 ± 1.14 mm. Las tasas de explotación fueron mayores a 0.6 para los sitios no protegidos y un alto componente de ejemplares pequeños (menos de 30 mm) en las recolectas, sugieren una sobre explotación de los adultos y sobrepesca en el reclutamiento. Con base en la estimación de la captura máxima sostenible (maximum sustainable yield), se recomienda algunas medidas reguladores de la pesquería como el control de un tamaño mínimo de desernbarque de 40 mm y la veda de la pesquería durante los rneses de reproducción (de julio a noviembre).
) estuvo en un ámbito de 3.31 a 3.48, el cual se encuentra entre los valores bajos informados para otros gastrópodos tropicales. La edad a la primera madurez sexual para ambos sexos combinados fue estimada en 29.20 ± 1.14 mm. Las tasas de explotación fueron mayores a 0.6 para los sitios no protegidos y un alto componente de ejemplares pequeños (menos de 30 mm) en las recolectas, sugieren una sobre explotación de los adultos y sobrepesca en el reclutamiento. Con base en la estimación de la captura máxima sostenible (maximum sustainable yield), se recomienda algunas medidas reguladores de la pesquería como el control de un tamaño mínimo de desernbarque de 40 mm y la veda de la pesquería durante los rneses de reproducción (de julio a noviembre).
References
Appeldoom R.S. 1984. The effect of size on mortality of small juvenile conchs (Strombus gigas L. and S. costatus G.). J. shellfish. Res. 4: 37- 43. [ Links ]
Appeldoom R.S. 1987. Assessment of mortality in an offshore population of queen conch, Strombus gigas L., in Southwest Puerto Rico. Fish. Bull. 85: 797- 804. [ Links ]
Appeldoom R.S. 1988a. Ontogenetic changes in natural mortality rate of queen conch, Strombus gigas (Mollusca: Mesogastropoda). Bull. Mar. Sci., 42: 159- 165. [ Links ]
Appeldoom R.S. 1988b. Age determination, growth, mortality and age of first reproduction in adult queen conch, Strombus gigas L., off Puerto Rico. Fish. Res. 6: 363-378. [ Links ]
Bell L.J. 1992. Reproduction and larval development of the West Indian Topshell, Cittarium pica (Trochidae), in the Bahamas. Bull. Mar. Sci. 51: 250-266. [ Links ]
Castilla J.C. & L.R. Durán. 1985. Human exclusion from the rocky intertidal zone of central Chile: the effects en C. concholepas (Gastropoda). Oikos 45: 391- 399. [ Links ]
Catterall C.P. & IR. Poiner. 1987. The potential impact of human gathering en shellfish populations, with reference to some NE Australian intertidal fiats. Oikos 50: 114-122. [ Links ]
Cervigón E, R. Cipriani, W. Fischer, L. Garibaldi, M. Hendrickx, A. Lemus, R. Marquéz, J.M. Pontiers, G. Robaina & B. Rodríguez. 1993. Field Guide to the commercial marine and brackish-water resources of the Northern coast of South America. FAO species identification sheets for fishery purposes, Rome, 490 p. [ Links ]
Cortés. J.A. & M.J. Risk. 1985. A reef under siltation stress: Cahuita, Costa Rica. Bull. Mar. Sci. 36: 339-356. [ Links ]
Debrot A.O. 1987 Biology and Management of the West Indian Topshell Cittarium pica of the Exuma Islands, Bahamas. Ph.D. Thesis University of Miami, Miarni USA. 193 p.
Debrot A.O. 1990a. Temporal aspects of population dynamies and dispersal bebaviour of the West Indian Topshell, Cittarium pica (Linnaeus), at selected sites in the Exuma Cays, Bahamas. Bull. Mar. Sci. 47: 431-447. [ Links ]
Debrot A.O. 1990b. Survival, growth, and fecundity of the West Indian Topshell, Cittarium pica (Linnaeus), in various rocky intertidal habitats of the Exuma Cays, Bahamas. The Veliger 33: 363-371. [ Links ]
Debrot A.O. & I. Nagelkerke. 2000. User perceptions en coastal resource state management options in Curazao. Rev. Biol. Trop. 48 Supl. 1: 95- 106. [ Links ]
Denyer, P. 1998. Historic-prehistoric earthquakes, seismic hazards, and Tertiary and Quartemary geology of the Gandoca-Mazanillo National Wildlife, Limón, Costa Rica. Rev. Biol. Trop. 46. Supl. 6: 237-250. [ Links ]
Hockey P.A.R. & A.L. Bosman. 1986. Man as an intertidal predator in Transkei: disturbance, community convergence and management of a natural food resource. Oikos 46: 3-14. [ Links ]
Hughes R.N. & D.J. Roberts. 1980. Growth and reproductive rates of Littorina rudis from three contrastes shores in North Wales, UK. Mar. Biol. 58: 47-54. [ Links ]
Jiménez, C. 2001. Bleaching and mortality of reef organisms during a warning event in 1995 on the Caribbean coast of Costa Rica. Rev. Biol. Trop. 49 Supl. 2: 233-238. [ Links ]
Keough M.J., G. Quinn & A. King. 1992. Correlations between human collecting and intertidal mollusc populations en rocky shores. Conservation Biol. 7: 378- 390. [ Links ]
Koch Y & M. Wolff. 1996. The mangrove snail Thais kiosquiformis Duclos: A case of life history adaptation to an extreme environment. J. Shellfish Res. 15: 421- 432. [ Links ]
Koch Y 1992. Zur Populationsökologie und ökologischen Rolle der Mangrovenschnecke Thais kiosquiformis im Golf von Nicoya, Costa Rica. Diplomarbeit University of Bremen, Bremen, Germany. 64 p. [ Links ]
Moreno C.A., J.R Sutherland & E Jara. 1984. Man as a predator in the intertidal zone of Southern Chile. Oikos 42: 155- 160. [ Links ]
Munro J.L. 1984. Estimation of Natural mortality rates from selectivity and catch length-frequency data. Fishbyte 2: 11-14. [ Links ]
Ortega S. 1987. The effect of human predation en the size distribution of Siphonaria gigas (Mollusca: Pulmonata) en the Pacific coast of Costa Rica. The Veliger 29: 251- 255. [ Links ]
Pauly D. 1983. Length-converted catch curves: a powerful tool for fisheries research in the tropics (part I). Fishbyte 1: 9-13. [ Links ]
Pauly D. & J.D. Munro. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte 2: 21. [ Links ]
Pérez M.P. & D.A. Aranda. 2000. Distribución, abundancia, densidad y morfometría de Strombus gigas (Mesogastropoda: Strombidae) en el arrecife Alacranes, Yucatán, México. Rev. Biol. Trop. 48 Supl. 1: 51- 57. [ Links ]
Pomeroy R.S. 1995. Community-based and co-management institutions for sustainable coastal fisheries management in Southeast Asia. Ocean & Coastal Management 27: 143-162. [ Links ]
Prince J.D., T.L. Sellers, W.B. Ford & S.R. Talbot. 1988. Recruitment, growth, mortality and population structure in a Southem Australian population of Haliotis ruba (Mollusca: Gastropoda). Mar. Biol. 100: 75- 82. [ Links ]
Randall H.A. 1964. A study of the growdi and other aspects of the biology of the West Indian Topshell Cittarium pica (Linnaeus). Bull. Mar. Sci. of the Gulf and Caribbean 14: 424-443. [ Links ]
Ruddle K. 1994. Local knowledge in the future management of inshore tropical marine resources and environments. Nature & Resources 30: 28-37. [ Links ]
Sainsbury K.J. 1982a. Population dynamics and fishery management of the paua, Haliotis iris. I. Population structure, growth, reproduction, and mortality. NZ J. Mar. Freshw. Res. 16: 147- 161. [ Links ]
Sainsbury K.J. 1982b. Population dynamics and fishery management of the paua, Haliotis iris. II. Dynamics and Management as examined using a size class population model. N.Z J. Mar. Freshw. Res 16: 163-173. [ Links ]
Schoenhuth, M. & U. Kievelitz. 1993. Participatory learning approaches (RRA & PRA) An introductory guide. Schriftenreihe der GTZ, Nr. 248. Universum Verlagsanstalt GmbH, Wiesbaden 1993, 183 p. [ Links ]
Sluczanowski P.R. 1984. A management oriented model of an abalone fishery whose substocks are subject to pulse fishing. Can. J. Fish. Aquat. Sci 41: 1008- 1014. [ Links ]
Sparre P. & Venema S.C. 1992. Introduction to tropical fish stock assessment. Part l-Manual. FAO Fisheries Technical Paper 306/1 Rev. 1376 p. [ Links ]
Udupa K.S. 1986. Statistical method of estimating the size at first maturity in fishes. Fishbyte 4: 8-10. [ Links ]
Wolff M. 1994. Population dynamics, life histories and management of selected invertebrates of the Southeast Pacific upwelling system. Habilitationsschrift University of Bremen, Bremen, Germany. [ Links ]
1 . Centre for Tropical Marine Ecology (ZMT), University of Bremen, Fahrenheitstrasse 6, 28359 Bremen Germany; finecr@yahoo.com ; mwolff@zmt.uni-bremen.de
2 . Centro de Investigación en Ciencias del Mar y Limnología, (CIMAR), Universidad de Costa Rica, 2060 San José, Costa Rica; javargas@biologia.ucr.ac.cr

