










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.2 San José Jun. 2002
en un bosque lluvioso tropical de Costa Rica
Alejandro G. Farji-Brener 1, Oscar Valverde 2, Leonardo Paolini 3, María de los Angeles La Torre 4, Estela Quintero 5, Elisa Bonaccorso 6, Luisa Arnedo 7 y Richard Villalobos 8
1 Lab. Ecotono, Depto. de Ecología, CRUB, Unidad Postal Universidad del Comahue, (8400) Bariloche, Argentina. Fax: 54 2944 422111; alefarji@yahoo.com. Autor a quien dirigir correspondencia.
2 Fundación Corredor Biológico Talamanca-Caribe, Ave. 8, Calle 11-13, San José, Costa Rica.
3 LIEY, Fac. de Ciencias, Universidad Nacional de Tucumán, Argentina.
4 Herbario Forestal, Universidad Agraria La Molina, Lima 12, Perú.
5 Fac. de Cs., Biología, Universidad de Antioquía, AA 1226, Medellín, Colombia.
6 Biología de Organismos, Universidad Simón Bolívar, Caracas 1080A, Venezuela.
7 Fac. de Cs., Biología, Universidad Javeriana, Cra 7, Calle 39-45, Bogotá, Colombia.
8 Proyecto Bio Darién, A. P. 2016, Ancón, Panamá.
Received 25-I-2001. Corrected 06-III-2002. Accepted 15-III-2002.
Abstract
Water retention on the leaf surface can be maladaptive to the plant because it increases the colonization of epiphylls and interferes with the physiologic processes of the leaf, diminishing the photosynthetic capacity. To test if leaf driptips facilitate leaf drying after rainfall in a tropical rain forest of Costa Rica, we (1) experimentally measured the capacity to retain water on leaf surfaces of 30 plant species before and after dritip removal, and (2) analyzed the development of driptips along forest strata. We expected leaf driptips to be less developed in the upper strata due to the environmental conditions of the canopy (i.e., high solar radiation, strong winds and low relative humidity), which favor the natural drying of leaves. The presence of driptips increased 100% the water run off capacity of leaves in all the analyzed species. Also, the development of leaf driptips was smaller in canopy species than in understory species. Additionally, they became less developed in canopy species as trees increased in height. These results support the hypothesis that the adaptive role of driptips is to facilitate the drying of leaf surfaces.
Key words: Costa Rica, driptips, forest strata, leaf drying, tropical rain forest.
"Without bold, regular patterns in nature,
ecologists do not have anything
very interesting to explain."
J. Lawton (1996)
Los factores abióticos pueden influir sobre las características foliares de tal forma que diferentes especies vegetales en un mismo ambiente presenten hojas morfológicamente similares (Crawley 1997). Por ejemplo, en ambientes áridos, un gran número de especies presenta hojas pequeñas y coriáceas, características que reducen la evapotranspiración. En ambientes tropicales de alta montaña, muchas especies presentan hojas con tricomas y envés de color claro, lo cual minimiza las grandes variaciones térmicas diarias (ver Niklas 1997). En los bosques húmedos tropicales, muchas especies poseen hojas con acumen o ápice alargado en forma de "punta de gotero" ("driptips" en inglés), lo cual ha sido interpretado como una adaptación para aumentar el drenaje del agua de la superficie foliar (Jungner 1891, Richards 1966, Dean y Smith 1978, Roth 1984). Un rápido secado luego de una lluvia podría minimizar la probabilidad de que la hoja sea colonizada por epífilas y hongos (Ivey y De-Silva 2001), los cuales disminuyen dramáticamente la cantidad de luz que llega al interior de la hoja (Coley et al. 1993). Adicionalmente, la permanencia de una capa de agua sobre la superficie foliar puede afectar los procesos fisiológicos que ocurren en la hoja, disminuyendo la tasa fotosintética (Dean y Smith 1978).
Si la presencia del acumen tiene como valor adaptativo facilitar el drenaje de la superficie foliar, esta característica no solo debería ser común en un gran número de especies, sino que también debería funcionar de forma semejante en las diferentes especies que habitan los bosques húmedos tropicales. Sin embargo, su efecto como facilitador del drenaje ha sido demostrado experimentalmente en pocas especies (Dean y Smith 1978, Ivey y De-Silva 2001). Adicionalmente, el desarrollo del acumen en las hojas debería relacionarse con su ubicación dentro de la estructura vertical del bosque. Si consideramos una sección vertical de un bosque tropical húmedo, existe un gradiente de condiciones climáticas entre los estratos superiores e inferiores. Las hojas que se encuentran en el estrato superior se secan más rápido que las que se encuentran en el estrato inferior porque reciben poco drenaje de hojas superiores, mayor irradiación solar, corrientes de aire más fuertes y están en un ambiente con menor humedad relativa. En consecuencia, si el acumen favorece el drenaje de la superficie foliar y éste es un carácter adaptativo, los acúmenes deberían estar menos desarrollados en los estratos superiores del bosque. Pese a que esta variación del tamaño relativo del acumen en el gradiente vertical del bosque ha sido propuesta como una evidencia de la función adaptativa de las hojas acuminadas (Richards 1966, Roth 1984, Williamson et al. 1983), no existen datos cuantitativos que documenten este patrón.
Nuestros objetivos fueron (1) demostrar experimentalmente si la presencia del acumen en las hojas facilita el drenaje del agua en un gran número de especies y (2) presentar las variaciones morfológicas del acumen a lo largo de un gradiente vertical del bosque de dos formas: comparando su desarrollo relativo (a) entre especies de dosel y sotobosque y (b) entre los diferentes estadios (plántula, juvenil y adulto) de especies típicas de dosel. Si el acumen facilita el drenaje del agua de las hojas y esta función es ventajosa para la planta, nosotros esperamos: (1) que su presencia disminuya el porcentaje del agua retenida en la superficie foliar en una amplia gama de especies, (2) que las hojas de especies de dosel posean acúmenes menos desarrollados que las especies de sotobosque y (3) que en las especies de dosel el desarrollo del acumen sea menor a medida que la planta asciende verticalmente dentro del bosque.
Materiales y métodos
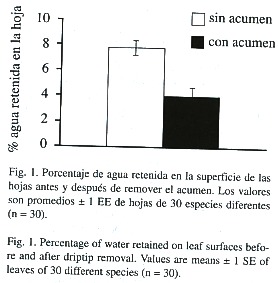
Realizamos este trabajo durante febrero del 2000 en la reserva Rara Avis, ubicada en Heredia, Costa Rica (10°20 N, 83°50 W), un bosque muy húmedo tropical a 650imsnm, con una precipitación promedio anual de 6i500imm. Para determinar la influencia del acumen sobre el drenaje del agua en la superficie foliar, recolectamos al azar hojas acuminadas de diferentes especies de plantas. A lo largo de aproximadamente 5ikm, cada 50im seleccionamos las tres especies con hojas acuminadas más cercanas (una planta adulta por especie). De cada planta recolectamos cinco hojas, las cuales llevamos al laboratorio. En total, encontramos 30 especies de plantas con hojas acuminadas que incluían diferentes formas de hojas y longitudes de acúmenes. Al final del muestreo, cada especie estuvo representada por 3 - 10 plantas y 15 - 50 hojas. En el laboratorio numeramos las hojas y elegimos con una tabla de números al azar tres hojas por especie, con las cuales realizamos el experimento. A cada hoja, luego de ser humedecida, le agregamos 5iml de agua en su parte axial con un gotero. El agua fue depositada lentamente en los diferentes sectores de la base de la hoja, simulando gotas de lluvia. Las hojas fueron colocadas con una inclinación promedio de 30°, con lo cual el agua se dirigía naturalmente hacia su ápice. Medimos el volumen del agua drenada esperando hasta 30is después de que la hoja paraba de gotear. Luego, a la misma hoja le recortamos el acumen con una tijera dejando su borde redondeado y repetimos el experimento. De esta forma pudimos cuantificar el efecto de la presencia del acumen sobre el drenaje del agua de la superficie foliar. El porcentaje de agua retenida en la superficie de cada hoja fue calculado como 100 - [(ml de agua drenada x 100) / 5]. En todos los experimentos usamos tres hojas de cada especie, utilizando el promedio para los análisis estadísticos. El porcentaje del agua retenida fue comparado entre hojas con y sin acumen con una prueba de t-pareada, utilizando cada hoja como su propio control.
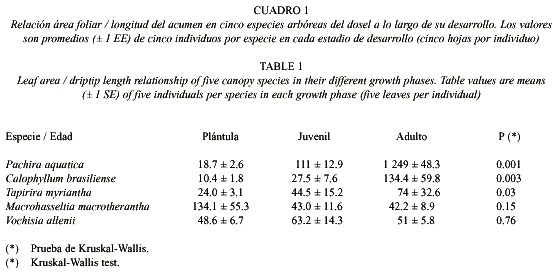
Para analizar el desarrollo relativo del acumen en los diferentes estratos del bosque, recorrimos durante un día áreas de bosque poco disturbado identificando todas las especies leñosas que pudimos. Definimos tres estratos verticales de acuerdo con la altura más común del dosel superior (aproximadamente 20im en este caso). Las especies leñosas fueron consideradas de sotobosque cuando los individuos adultos eran menores de 5im de altura, de estrato intermedio cuando tenían entre 5 - 15im de altura y de dosel cuando eran mayores de 15im de altura. Para analizar variaciones ontogénicas en la morfología foliar de las especies de dosel, utilizamos solo aquellas especies de las cuales pudimos localizar e identificar individuos de menos de 0.50im ("plántulas"), de 1 - 3im ("juveniles") y de más de 10im ("adultos"). Seleccionamos al azar tres individuos por especie y cinco hojas / individuo (ubicadas en diferentes sectores de la planta), a las cuales les medimos su largo, ancho (en el punto central de la lámina de la hoja), y el largo del acumen (a partir del punto de intersección de la proyección de los bordes de la hoja). Con la medida del largo y el ancho de cada hoja estimamos su área utilizando la fórmula 2/3 largo x ancho. Pese a que esta fórmula es muy general y no necesariamente estima el área de la hoja con exactitud, es útil para comparar en términos relativos áreas de diferentes tipos de hojas. Finalmente, estimamos la importancia relativa del acumen (IRA) como la relación área foliar / longitud del acumen, en donde menores valores del IRA implican una mayor importancia relativa del acumen. De esta forma pudimos comparar el desarrollo relativo del acumen entre especies con hojas de tamaño diferente. Debido a la imposibilidad de dividir entre cero, las hojas con bordes redondeados fueron consideradas con un acumen de 0.1icm de longitud para poder calcular la relación área foliar / longitud del acumen. Para comparar el desarrollo relativo del acumen entre especies de dosel, estrato intermedio y sotobosque, utilizamos el IRA promedio por especie. Para comparar dicha variación entre los diferentes estadios de desarrollo de las especies de dosel, utilizamos para cada especie un IRA promedio por individuo. Utilizamos el análisis de varianza de una vía no-paramétrico (Kruskal-Wallis) para realizar las comparaciones, dado que los datos no se ajustaban a los supuestos de normalidad y homocedacea (Zar 1999).
Resultados
La presencia del acumen facilitó e incrementó en un 100% la capacidad de drenaje en todas las hojas de las especies analizadas en el experimento, ya que el porcentaje de agua retenida en la superficie foliar fue siempre menor cuando las hojas tenían acumen que cuando no lo tenían (T = 6.2, n = 30, P < 0.001, Fig. 1). Por otra parte, encontramos un total de 37 especies arbóreas: 10 de dosel, 7 de estrato intermedio y 20 de sotobosque (Anexo 1). La relación área foliar / longitud del acumen fue significativamente mayor en las hojas de las especies de dosel (1i568 ± 754) comparadas con las hojas de las especies de sotobosque (221 ± 59) y estrato intermedio (226 ± 88.3) (promedio ± 1 error estándar, F = 3.5, g.l. = 2, p = 0.04). De las diez especies de dosel, sólo pudimos encontrar plántulas, juveniles y adultos en cinco especies. Tres de esas cinco especies (Pachira aquatica, Calophyllum brasiliense y Tapirira myriantha) presentaron cambios significativos en la relación área foliar / longitud del acumen entre los diferentes estadios de desarrollo. En estas especies el acumen fue disminuyendo en importancia relativa a medida que los individuos crecían en altura (Cuadro 1).
Discusión
Dos líneas de evidencia presentadas en este estudio apoyan la hipótesis de que el acumen es una estructura foliar cuyo significado funcional es facilitar el secado de las hojas después de la lluvia. Primero, la presencia del acumen facilitó el drenaje del agua en la superficie foliar de un gran número de especies. Segundo, el desarrollo relativo del acumen fue más importante en los estratos bajos del bosque que en los altos. Este patrón se presentó tanto al comparar especies de diferentes gremios de regeneración, como al comparar distintos estadios de desarrollo de las especies de dosel.
Independientemente de que las especies analizadas diferían en sus características foliares como forma y textura, la capacidad de las hojas de drenar agua incrementó el doble con la presencia del acumen. Mantener una capa de agua en la superficie de las hojas puede ser desventajoso para las plantas. Por ejemplo, se ha demostrado que una mayor humedad incrementa la colonización de hongos y epífilas en la superficie foliar, reduciendo la intercepción lumínica de las hojas hasta un 85% (Coley et al. 1993, Ivey y DeSilva 2001). Plantas regadas experimentalmente durante un año pueden incrementar hasta 11 veces su cobertura de epífilas, la cual en dos años cubre el 45% de la superficie foliar reduciendo la fotosíntesis entre 20 - 30% (Sand-Jensen 1977, Coley et a.l 1993, Coley y Kursar 1996). Sin embargo, pese a que una mayor humedad determina una mayor colonización por epífilas y que este estudio demuestra que la presencia del acumen disminuye el agua de la superficie foliar, aún no existen estudios experimentales que demuestren si la presencia del acumen disminuye la probabilidad de colonización por epífilas (Coley y Kursar 1996, pero ver Ivey y DeSilva 2001). Independientemente de la presencia de epífilas, la acumulación del agua sobre la superficie foliar per se puede incrementar la probabilidad de infección por patógenos (Gregory 1971) y alterar el intercambio gaseoso de la hoja, disminuyendo la tasa de crecimiento de la planta (Dean y Smith 1978).
Por otra parte, el mayor desarrollo relativo del acumen encontrado en los estratos bajos del bosque, donde la humedad es mayor, apoya la idea de que esta estructura tiene como valor adaptativo facilitar el drenaje de la superficie foliar. En los estratos superiores del bosque las hojas están expuestas a escaso drenaje recibido de otras hojas, mayor irradiación solar, vientos más fuertes y menor humedad relativa. Estas características facilitan el secado de las hojas luego de una lluvia, por lo cual el desarrollo del acumen en los estratos superiores del bosque no sería funcionalmente tan importante como en los estratos inferiores. Si las características ambientales del dosel son factores de selección que determinan un menor desarrollo relativo del acumen en las especies arbóreas, este menor desarrollo debería presentarse también en otros sitios del bosque cuyas características ambientales sean similares a las del dosel. Acorde con esta idea, la variabilidad que encontramos en la relación área foliar / longitud del acumen parece asociarse no solo a los diferentes estratos verticales del bosque, sino también a los requerimientos de micro-hábitat de cada especie. Por ejemplo, dentro de las especies de sotobosque, la que presentó el acumen menos desarrollado fue Tetrochidium euryphyllum, una especie de tamaño reducido pero muy asociada a claros u orillas de caminos, donde las condiciones de radiación y humedad relativa pueden ser similares a las encontradas en el dosel. Por el contrario, las especies con el acumen más desarrollado como Eugenia sp. o Schefflera sp. son típicas de sotobosque cerrado, donde la insolación es menor y la humedad relativa mayor (Anexo 1).
Se ha sugerido que al disminuir el tamaño de la gota que cae de la hoja, una posible función adaptativa del acumen es minimizar la erosión del suelo alrededor de la planta (Williamson 1981). Si ésta fuera la principal función adaptativa del acumen, las hojas de los estratos cercanos al suelo deberían poseer acúmenes menos desarrollados que las hojas de los estratos intermedios, ya que el nivel de erosión por goteo se incrementa con la altura desde la cual cae la gota (Williamson et al. 1983). Sin embargo, nuestros datos no avalan esa hipótesis, ya que las plántulas de las especies de dosel mostraron hojas con acúmenes muy desarrollados a menos de 30icm del suelo (Cuadro 1).
Este estudio demuestra experimentalmente que la presencia del acumen disminuye el porcentaje de retención de agua en la superficie foliar, independientemente de las diferencias morfológicas que pueden existir entre las hojas de diferentes especies (ver Ivey y DeSilva 2001). Adicionalmente, el desarrollo del acumen tanto a nivel de especie como a nivel de individuo es más importante en los estratos del bosque en donde el secado de la hoja es mas lento (Anexo 1). Por otra parte, en el bosque seco las especies con hojas acuminadas están casi ausentes, mientras que son muy frecuentes en los bosques lluviosos tropicales (Gómez 1986). Por ejemplo, en el bosque tropical de Sinharaja, Sri Lanka, el 90% de las especies arbóreas poseen hojas acuminadas (Roth 1984). En Costa Rica, el 92% de las especies que habitan las selvas húmedas poseen hojas con acumen (Gómez 1986), característica también común en la mayoría de las especies de los bosques montanos húmedos (Kappelle 1996, Kappelle y Leal 1996). Por lo tanto, su efecto de acelerar el drenaje de la superficie foliar y su mayor presencia, tanto en los estratos del bosque como en los ambientes con mayores niveles de humedad, apoyan la hipótesis de que el acumen es una estructura foliar cuyo valor adaptativo es el de facilitar el secado de las hojas.
Agradecimientos
A la Organización para Estudios Tropicales (OET - OTS) por su apoyo logístico y técnico y a M.L. Tadey y dos revisores anónimos por sus comentarios.
Resumen
La acumulación de agua en la superficie foliar puede ser nociva para la planta porque incrementa la colonización de epífilas e interfiere con los procesos fisiológicos de la hoja, disminuyendo la capacidad fotosintética. Para probar si la presencia del acumen en las hojas facilita el drenaje de la superficie foliar en un bosque tropical lluvioso de Costa Rica, (1) medimos experimentalmente la capacidad de retener agua en la superficie foliar de 30 especies antes y después de remover el acumen y (2) analizamos el desarrollo relativo del acumen en los diferentes estratos verticales del bosque. Esperábamos que los acúmenes se encontraran menos desarrollados en los estratos superiores, debido a que las condiciones ambientales allí existentes (i.e., alta insolación, fuertes vientos, baja humedad relativa) favorecen el secado de las hojas. La presencia del acumen incrementó en un 100% la capacidad de drenar agua de la superficie foliar en todas las especies analizadas. Además, el desarrollo del acumen fue menor en las especies de dosel comparadas con las especies de estratos más bajos. Adicionalmente, las hojas de las especies de dosel se hacen menos acuminadas a medida que el árbol crece en altura. Estos resultados apoyan la hipótesis de que la función adaptativa del acumen es facilitar el secado de la superficie foliar.
Referencias
Coley, P., T. Kursar & J.L. Machado. 1993. Colonization of tropical rain forest leaves by epiphylls: Effects of site and host plant leaf lifetime. Ecology 74: 619-623. [ Links ]
Coley, P. & T. Kursar. 1996. Causes and consequences of epiphyll colonization, p. 337-362. In S. Mulkey, R. Chazdon & A. Smith (eds.). Tropical forest plant ecophysiology. Chapman and Hall, Nueva York. [ Links ]
Crawley, M. 1997. Life history and environment, p. 73- 131. In M. Crawley (ed.). Plant ecology. Blackwell Science, Nueva York. [ Links ]
Dean, J. & A. Smith. 1978. Behavioral and morphological adaptations of a tropical plant to high rainfall. Biotropica 10: 152-154. [ Links ]
Gómez, L.D. 1986. Vegetación de Costa Rica. Volumen I. Universidad Estatal a Larga Distancia, Costa Rica. 322 p. [ Links ]
Gregory, P. 1971. The leaf as spore trap, p. 239-244. In R. Feece & C. Dickinson (eds.). Ecology of leaf surface microorganisms. Academic, Nueva York. [ Links ]
Ivey, C. & N. DeSilva. 2001. A test of the function of drip tips. Biotropica 33: 188-191. [ Links ]
Jungner, J. 1891. Anpassungen der Pflanzen an das Klima in den Gegendren der regenreichen Kamerungebirge. Bot. Zbl. 47: 353-360. [ Links ]
Kapelle, M. 1996. Los bosques de roble (Quercus) de la Cordillera de Talamanca, Costa Rica. Biodiversidad, ecología, conservación y desarrollo. INBIO-Univ. de Amsterdam, Heredia, Costa Rica. 319 p. [ Links ]
Kapelle, M. & M. Leal. 1996. Changes in leaf morphology and foliar status along a successional gradient in Costa Rican upper montane Quercus forest. Biotropica 28: 331-344. [ Links ]
Mabberley, D.J. 1990. The plant book. Cambridge, Londres. 707 p. [ Links ]
Niklas, K.J. 1997. The evolutionary biology of plants. Chicago University, Chicago. 449 p. [ Links ]
Lawton, J. 1996. Patterns in ecology. Oikos 75: 145-147. [ Links ]
Richards, P. 1966. The tropical rain forest. Cambridge University, Londres. 450 p. [ Links ]
Roth, I. 1984. Stratification of tropical forests as seen in leaf structure. Dr. W. Junk, The Haghe, Holanda. 345 p. [ Links ]
Sand-Jensen, K. 1977. Effect of epiphytes on eelgrass photosynthesis. Aquat. Bot. 3: 55-63. [ Links ]
Williamson, G. 1981. Driptips and splash erosion. Biotropica 13: 228-231. [ Links ]
Williamson, G., A. Romero, J. Armstrong, T. Gush, A. Hruska, P. Klass & J. Thompson. 1983. Driptips, drop size and leaf drying. Biotropica 15: 232-234. [ Links ]
Zar, J.H. 1999. Biostatistical analysis. Prentice Hall, Nueva Jersey. 663 p. [ Links ]


